氣溫升高與干旱脅迫對寧夏枸杞光合作用的影響
2015-01-19 02:49:38宋麗華
生態(tài)學(xué)報(bào) 2015年18期
趙 琴, 潘 靜, 曹 兵,*, 宋麗華
1 寧夏大學(xué)農(nóng)學(xué)院, 銀川 750021 2 寧夏防沙治沙職業(yè)技術(shù)學(xué)院, 永寧 750105
氣溫升高與干旱脅迫對寧夏枸杞光合作用的影響
趙 琴1, 潘 靜2, 曹 兵1,*, 宋麗華1
1 寧夏大學(xué)農(nóng)學(xué)院, 銀川 750021 2 寧夏防沙治沙職業(yè)技術(shù)學(xué)院, 永寧 750105
以寧夏枸杞1年生苗木為材料,采用開頂式生長室模擬增溫環(huán)境,設(shè)置兩個(gè)溫度水平(正常環(huán)境溫度,增溫 = 正常環(huán)境溫度 + 2.5—3.7 ℃)和3個(gè)土壤水分水平(正常水分條件(田間最大持水量的70%—75%)、中度干旱處理(田間最大持水量的50%—55%)和重度干旱處理(田間最大持水量的35%—40%)),研究氣溫升高和干旱脅迫對寧夏枸杞光合作用的影響。結(jié)果表明:(1)在增溫條件下,中度和重度干旱處理下的凈光合速率比對照(正常供水)分別下降17.5%、48.9%,氣孔導(dǎo)度平均下降了3.9%,水分利用效率僅為正常環(huán)境溫度下的57.8%。(2)在氣溫升高和干旱脅迫交互作用下,枸杞葉片的凈光合速率、氣孔導(dǎo)度、胞間CO2濃度顯著下降的同時(shí),增溫處理又加劇了枸杞植株的蒸騰耗水,從而導(dǎo)致枸杞葉片水分的利用效率和CO2同化能力降低。(3)氣溫升高和干旱脅迫交互作用降低了枸杞的PSⅡ活性中心的光能轉(zhuǎn)換效率、使光合機(jī)構(gòu)和PSⅡ反應(yīng)中心受到損傷,從而導(dǎo)致枸杞光合作用效率下降。(4)氣溫升高加劇了干旱脅迫對寧夏枸杞葉片凈光合速率和水分利用效率的減小作用,即氣溫升高加劇了干旱脅迫對寧夏枸杞光合作用的抑制作用。
氣溫升高; 干旱脅迫; 寧夏枸杞; 光合作用; 葉綠素?zé)晒?/p>
由大氣CO2濃度升高導(dǎo)致“溫室效應(yīng)”不斷加劇,使全球地表平均氣溫在20世紀(jì)期間增加了(0.6±0.2)℃[1-3]。據(jù)預(yù)測,21世紀(jì)末氣溫將升高1.5—4.5 ℃[1]。全球氣候變暖將使降水格局發(fā)生變化,進(jìn)而引起地區(qū)間水資源量的差異[4]。中國北方,尤其是西北地區(qū)為我國生態(tài)環(huán)境脆弱區(qū),降雨量少、蒸發(fā)量大,干旱成為植物存活與生長的限制性因子。由大氣CO2濃度升高導(dǎo)致的氣溫升高,可能使得土壤蒸發(fā)量增大,干旱化趨勢加重,影響植物生長發(fā)育[5]。氣候變化對植物生長與生理的影響成為植物生理生態(tài)學(xué)的研究熱點(diǎn)[4-5]。
寧夏枸杞(LyciumbarbarumL.)果實(shí)富含多種營養(yǎng)成分和微量元素,有很高的營養(yǎng)價(jià)值和藥用保健功效[6-7],具有抗旱、耐鹽堿、耐瘠薄的特點(diǎn)[8];因其生態(tài)適應(yīng)性強(qiáng)、經(jīng)濟(jì)價(jià)值高,是我國重要的特種經(jīng)濟(jì)植物資源之一,生態(tài)、 經(jīng)濟(jì)、社會(huì)效益顯著。枸杞產(chǎn)業(yè)是寧夏特色經(jīng)濟(jì)林產(chǎn)業(yè)和農(nóng)業(yè)優(yōu)勢特色產(chǎn)業(yè)重要組成部分,作為枸杞主產(chǎn)區(qū)的寧夏,在全球氣候變化的背景下,近五十多年來也呈現(xiàn)出明顯的氣候變暖趨勢[9-12],且氣溫升高與干旱加劇現(xiàn)象將更加明顯[11-12],這些變化對寧夏枸杞生長與生理影響值得關(guān)注。本研究以寧夏枸杞為對象,測定氣溫升高與干旱脅迫下其光合作用的變化,以揭示全球氣候變化對枸杞生長與生理的影響機(jī)制,為枸杞育種與栽培提供參考。
1 材料與方法
1.1 試驗(yàn)地點(diǎn)
本試驗(yàn)于2012 年4 月到10 月在寧夏大學(xué)試驗(yàn)農(nóng)場進(jìn)行。該場位于寧夏中部黃河沖積平原永寧縣境內(nèi)(北緯38°47′07″,東經(jīng)106°04′00″,海拔2600 m),地處西北內(nèi)陸,屬中溫帶干旱氣候區(qū),年均≥10 ℃積溫3300 ℃,無霜期140—160 d,年均日照時(shí)數(shù)3000 h,日溫差13 ℃,年降水量180至200 mm,年均氣溫8.5 ℃。
1.2 試驗(yàn)材料
供試材料為寧杞1號1年生扦插苗,購于寧夏農(nóng)林科學(xué)院枸杞研究所。
1.3 試驗(yàn)設(shè)計(jì)與方法
試驗(yàn)采用雙因素設(shè)計(jì):溫度為正常環(huán)境氣溫(T1,對照)和增溫 (T2) 兩個(gè)處理;3個(gè)土壤水分處理:正常水分條件(W1,田間最大持水量的70%—75%,對照)、中度干旱處理(W2, 田間最大持水量的50%—55%)和重度干旱處理(W3, 田間最大持水量的35%—40%);共6個(gè)處理組合,每處理3次重復(fù),每重復(fù)為5株盆栽枸杞苗,共90盆。
采用自制開頂式生長室模擬控制溫度,由金屬框架和厚度為3.8 mm的無色透明玻璃制成。氣室為高2.5 m、底部為邊長1.1 m的正八邊形(內(nèi)徑3 m),上部0.5 m向內(nèi)傾斜呈45°,即頂部開口直徑為2 m。模擬增溫處理的開頂式生長室八面均安裝有無色透明玻璃,而正常環(huán)境氣溫處理(對照)的生長室東西與南北對立四面留裝玻璃,以便通風(fēng),使室內(nèi)與外界自然環(huán)境條件保持一致,采用溫度記錄儀觀測兩種處理下的空氣溫度變化。(白天模擬增溫處理的平均溫度比對照高 3.7 ℃,夜晚比對照高2.5 ℃)。
盆栽土壤水分采用自動(dòng)化滴灌系統(tǒng)控制(GG-006A型土壤濕度控制自動(dòng)灌溉系統(tǒng),上海艾美克電子有限公司)。4月下旬將枸杞苗木單株定植于盆內(nèi)(高23 cm,口徑33 cm),置于開頂器室內(nèi),正常供水,5月中旬開始處理。
1.4 測定指標(biāo)
試驗(yàn)處理90 d時(shí),于9:00—11:00,每處理選擇5—8片功能葉片,采用LCi便攜式光合測定系統(tǒng)測定枸杞葉片的凈光合速率(Pn) 、蒸騰速率(E) 、氣孔導(dǎo)度(Gs)、胞間CO2濃度(Ci)等指標(biāo),并計(jì)算水分利用效率(WUE =Pn/E);采用FMS-2葉綠素?zé)晒鈨x測定初始熒光(Fo),最大熒光(Fm),計(jì)算可變熒光(Fv=Fm-Fo),PSⅡ原初光能轉(zhuǎn)換效率(Fv/Fm),PSⅡ的潛在活性(Fv/Fo)(測定前葉片暗適應(yīng)20—30 min,每個(gè)處理重復(fù)8次)。
1.5 數(shù)據(jù)處理與分析
采用Excel軟件進(jìn)行數(shù)據(jù)整理,DPS7.05進(jìn)行統(tǒng)計(jì)分析。

圖1 氣溫升高與干旱脅迫對寧夏枸杞凈光合速率的影響 Fig.1 Effect of elevated temperature and drought stress on net Photosynthesis rate of L. barbarum.T: 溫度處理temperature;W: 干旱處理water; ns: P≥ 0.05; * P < 0.05;** P < 0.01; *** P < 0.001,圖中不同大寫字母表示各處理在α=0.01水平差異極顯著
2 結(jié)果與分析
2.1 氣溫升高與干旱脅迫對寧夏枸杞氣體交換特性的影響
2.1.1 氣溫升高與干旱脅迫對凈光合速率的影響
光合作用是植物最重要的生理生化活動(dòng),為植物提供有機(jī)營養(yǎng)物質(zhì)和能量,是生物界賴以生存的基礎(chǔ)[13]。光合作用受光照強(qiáng)度、水分、溫度、CO2濃度、土壤等環(huán)境因子的影響,其中溫度和水分是影響植物光合作用和蒸騰作用的主要生態(tài)因子[14]。
由圖1可以看出,不同溫度與土壤水分處理組合下,寧夏枸杞葉片的凈光合速率(Pn)有顯著差異(P= 0.0002);隨著干旱脅迫程度的加劇,Pn顯著下降;在正常環(huán)境氣溫條件下,中度和重度干旱處理分別比正常供水處理下降32.4%、54.7%;在增溫條件下,與對照(正常供水)相比,中度和重度干旱處理下的Pn分別下降17.5%、48.9%,說明干旱脅迫降低Pn;增溫處理下枸杞葉片的平均Pn比正常環(huán)境氣溫的平均Pn降低12.7%,但差異不顯著(P>0.05)。
2.1.2 氣溫升高與干旱脅迫對氣孔導(dǎo)度和胞間CO2濃度的影響
氣孔是植物吸收CO2和蒸騰失水的通道,當(dāng)植物處于干旱脅迫時(shí),會(huì)通過調(diào)整氣孔的開度來防止體內(nèi)水分的散失并維持一定的光合作用。由圖2可知,氣溫升高和干旱脅迫交互作用對枸杞葉片氣孔導(dǎo)度(Gs)的影響達(dá)到極顯著水平(P= 0.0061)。在正常環(huán)境氣溫下,隨著干旱脅迫程度的加劇,Gs顯著下降,中度和重度干旱處理的Gs分別比正常供水處理的下降35.2%和64.6%;在增溫條件下,隨著干旱脅迫程度的加劇,Gs表現(xiàn)為先下降后升高的趨勢,重度干旱處理的Gs為正常供水處理的72.7%;與正常環(huán)境氣溫處理相比,增溫處理下枸杞葉片的平均Gs下降3.9%。
由圖3可看出,氣溫升高和干旱脅迫交互作用對枸杞葉片胞間CO2濃度(Ci)的影響也達(dá)到極顯著水平(P= 0.001)。在正常環(huán)境氣溫下,中度和重度干旱處理的Ci分別比正常供水處理的下降11.5%和20.6%;增溫處理下,隨著干旱脅迫的加劇,胞間CO2濃度表現(xiàn)為先降低后增加的趨勢,但差異不顯著;增溫處理有降低Ci的作用,但降幅不大。

圖2 氣溫升高與干旱脅迫對寧夏枸杞葉片氣孔導(dǎo)度的影響 Fig.2 Effect of elevated temperature and drought stress on Stomatal conductance of L. barbarum

圖3 氣溫升高與干旱脅迫對寧夏枸杞胞間CO2濃度的影響 Fig.3 Effect of elevated temperature and drought stress on intercellular CO2 of L. barbarum
2.1.3 氣溫升高與干旱脅迫對蒸騰速率和水分利用效率的影響
蒸騰作用能夠通過蒸騰拉力引起根系吸收水分和礦物質(zhì),蒸騰速率大小反應(yīng)出植物耗水情況。由圖4可知,不同溫度與土壤水分處理間寧夏枸杞葉片的蒸騰速率(Tr)間差異極顯著(P= 0.0096)。隨著干旱脅迫程度的加劇,Tr顯著下降;重度干旱處理下的Tr僅為正常供水處理的57.1%;隨著氣溫的升高,Tr增高,與正常環(huán)境氣溫相比,增溫處理的平均Tr升高了10.4%。
水分利用效率(WUE)指植物消耗單位水分所形成的干物質(zhì)重量,是植物光合、蒸騰特性的綜合表現(xiàn)[15]。由圖5可看出,不同溫度與土壤水分處理間,枸杞葉片的水分利用效率(WUE)差異極顯著(P= 0.0001);在正常環(huán)境氣溫處理中,隨著干旱脅迫程度的加劇,WUE明顯下降,重度干旱處理的WUE為正常供水處理的35.5%;增溫處理下枸杞葉片的平均WUE為正常環(huán)境氣溫處理的57.8%,說明干旱脅迫和氣溫升高降低了枸杞的水分利用效率。
2.2 氣溫升高與干旱脅迫對寧夏枸杞葉綠素?zé)晒馓匦缘挠绊?/h3>
葉綠素?zé)晒夥治黾夹g(shù)具有快速、簡便、無損傷等優(yōu)點(diǎn),被稱為天然探針,在分析葉片光合作用過程方面具有獨(dú)特的作用,它能夠內(nèi)在地反映光系統(tǒng)對光能的吸收、傳遞、耗散、分配,快速且較靈敏地診斷植物光合狀況,以及揭示植物響應(yīng)外界環(huán)境因素的方式[16-17]。

圖5 氣溫升高與干旱脅迫對寧夏枸杞水分利用效率的影響 Fig.5 Effect of elevated temperature and drought stress on water use efficiency of L. barbarum
2.2.1 氣溫升高與干旱脅迫對枸杞葉綠素初始熒光、最大熒光、可變熒光的影響
初始熒光(Fo)是PSⅡ反應(yīng)中心處于完全開放時(shí)的熒光產(chǎn)量。Fo它減少表明光合色素含量減少,而增加則表明PSⅡ反應(yīng)中心受到損傷[18]。由表1看出,在正常環(huán)境氣溫下,隨著干旱脅迫的加劇,枸杞葉片的Fo略有下降;增溫處理下,干旱脅迫有增大初始熒光的作用趨勢;與正常環(huán)境氣溫處理相比,增溫處理的平均Fo升高了21.5%,說明氣溫升高增大了枸杞植株的初始熒光,導(dǎo)致枸杞植株光合作用效率下降。
最大熒光(Fm)是PSⅡ反應(yīng)中心完全關(guān)閉時(shí)的熒光產(chǎn)量,它能反映PSⅡ的電子傳遞情況。由表1可知,增溫處理的Fm明顯低于正常環(huán)境氣溫處理,增溫處理的平均Fm是正常環(huán)境氣溫處理的87.6%;在正常環(huán)境氣溫處理下,干旱脅迫有降低Fm的作用趨勢。說明氣溫升高降低了枸杞植株的最大熒光。
可變熒光(Fv)作為PSⅡ反應(yīng)中心活性大小的相對指標(biāo),與PSⅡ氧化一側(cè)的水裂解釋放O2過程有關(guān)[19]。由表1可看出,溫度對枸杞葉綠素?zé)晒鈪?shù)Fv的影響作用差異顯著(P= 0.037);在同一干旱水平下,與正常環(huán)境氣溫處理相比,增溫正常供水處理、中度干旱處理、重度干旱處理的Fv分別為正常環(huán)境氣溫處理的74.2%、74.5%、87.1%,說明增溫降低了Fv;在正常環(huán)境氣溫下,干旱脅迫有降低可變熒光的作用趨勢。說明氣溫升高和干旱脅迫降低了枸杞植株的可變熒光。
2.2.2 氣溫升高與干旱脅迫對PSⅡ最大光化學(xué)效率、PSⅡ的潛在活性的影響
Fv/Fm(PSⅡ最大光化學(xué)效率)是綠色植物葉片光化學(xué)反應(yīng)的一個(gè)重要參數(shù),能反映出PSⅡ反應(yīng)中心原初光能轉(zhuǎn)化效率[20]。由表1可知,溫度處理對枸杞葉片F(xiàn)v/Fm的影響作用差異顯著(P= 0.0131);與正常環(huán)境氣溫處理相比,增溫處理的平均Fv/Fm下降了 11.1%,說明增溫有降低枸杞葉片PSⅡ最大光化學(xué)效率的趨勢。
Fv/Fo表示PSⅡ的潛在活性,由表1看出,溫度處理對枸杞Fv/Fo的影響作用差異顯著(P= 0.0194);在同一干旱水平下,增溫處理的Fv/Fo明顯低于正常環(huán)境氣溫處理,增溫處理的平均Fv/Fo為正常環(huán)境氣溫處理平均Fv/Fo的69.6%;在正常環(huán)境氣溫處理下,干旱脅迫降低了PSⅡ潛在活性;但模擬增溫處理后,干旱脅迫對枸杞葉片PSⅡ潛在活性的降低作用不明顯。
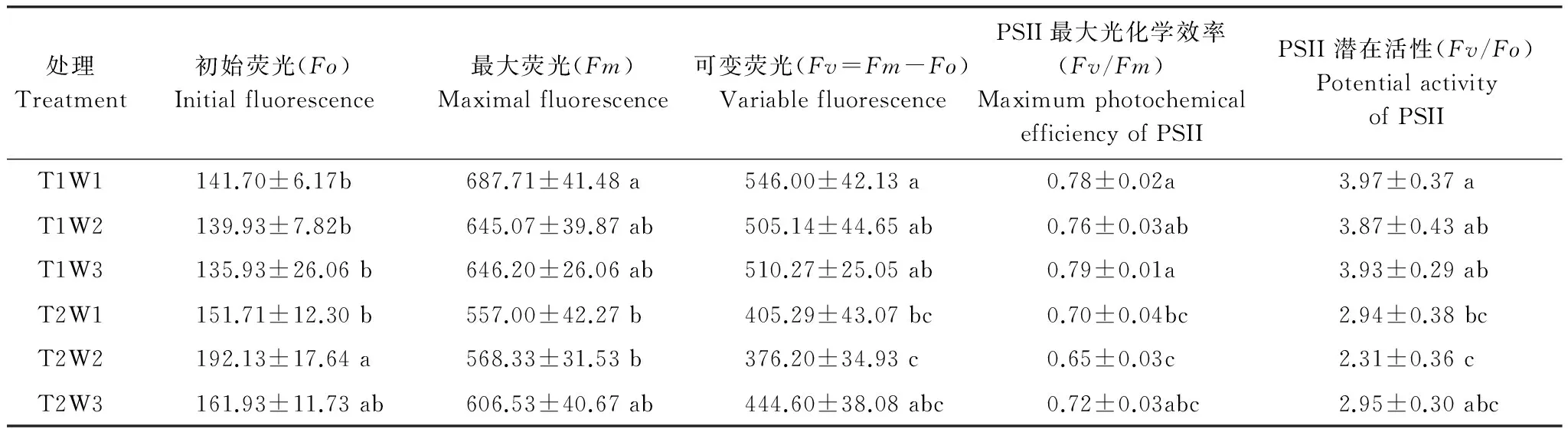
表1 氣溫升高與干旱脅迫對枸杞植株葉綠素?zé)晒鈪?shù)的影響
3 結(jié)論與討論
水分和溫度是影響植物光合作用的主要環(huán)境因子[21-22]。當(dāng)植物處于干旱脅迫狀態(tài)時(shí),其葉片氣孔導(dǎo)度會(huì)下降,造成葉內(nèi)細(xì)胞間隙CO2濃度降低從而導(dǎo)致光合速率下降。目前關(guān)于影響光合速率下降的原因認(rèn)為:一個(gè)是氣孔因素,主要受氣孔導(dǎo)度的影響,另一個(gè)為非氣孔因素,受葉肉細(xì)胞光合活性的影響[23]。在模擬增溫的環(huán)境下,正常供水和中度干旱處理的寧夏枸杞葉片氣孔導(dǎo)度和胞間CO2濃度表現(xiàn)為降低趨勢,而重度干旱處理的氣孔導(dǎo)度和胞間CO2濃度與中度干旱處理相比,又表現(xiàn)為升高的趨勢。說明正常供水和中度干旱處理中,枸杞葉片凈光合速率降低的原因可能是氣孔因素,即氣孔關(guān)閉導(dǎo)致的;這與付士磊[24]在研究干旱脅迫對楊樹光合生理指標(biāo)的影響的試驗(yàn)結(jié)果一致;而對于增溫、重度干旱處理下,枸杞葉片凈光合速率的降低時(shí),胞間CO2濃度上升,這說明凈光合速率降低的主要原因是非氣孔因素[21,23],即葉肉細(xì)胞的光合活性下降,主要是因?yàn)樗置{迫導(dǎo)致核酮糖-1,5-二磷酸羧化酶活性降低,因而CO2與核酮糖-1,5-二磷酸(RuBP)的羧化反應(yīng)速率降低[25]。
有研究表明,植物在高溫脅迫下常伴隨著Fo上升;在25—40 ℃時(shí)Fo增幅不大[26],而在45—50 ℃高溫下,熒光參數(shù)Fo升高,PSⅡ反應(yīng)中心發(fā)生可逆失活[27]。本試驗(yàn)中,增溫處理的Fo比正常環(huán)境氣溫處理的有所增加,但增幅不大,說明氣溫升高導(dǎo)致PSⅡ反應(yīng)中心受到一定程度的破壞。Fm是PSⅡ反應(yīng)中心完全關(guān)閉時(shí)的熒光產(chǎn)量,F(xiàn)m下降,表明脅迫條件下PSⅡ反應(yīng)中心處于完全關(guān)閉時(shí)的量子產(chǎn)量下降,通過PSⅡ的電子傳遞量減少,F(xiàn)m的下降是衡量植物葉片發(fā)生光抑制的重要特征[28-29];采用模擬增溫處理寧夏枸杞苗木,葉片的Fv、Fm、Fv/Fo、Fv/Fm均顯著下降;而在正常環(huán)境氣溫處理中,干旱脅迫又在一定程度上降低了Fm、Fv、Fv/Fo,說明氣溫升高與干旱脅迫對寧夏枸杞光合機(jī)構(gòu)造成了損傷[30]。通常,植物處于干旱脅迫時(shí)Fv/Fm會(huì)顯著降低[31-32],但試驗(yàn)中發(fā)現(xiàn),干旱脅迫對枸杞Fv/Fm的影響不顯著,這可能是因?yàn)殍坭娇购敌暂^強(qiáng)的原因,還待于進(jìn)一步研究。
綜上所述,在氣溫升高和干旱脅迫交互作用下,枸杞葉片的凈光合速率Pn、氣孔導(dǎo)度Gs、胞間CO2濃度Ci顯著下降的同時(shí),增溫處理又加劇了枸杞植株的蒸騰耗水,從而導(dǎo)致枸杞葉片水分的利用效率和CO2同化能力降低。氣溫升高和干旱脅迫交互作用降低了枸杞的PSⅡ活性中心的光能轉(zhuǎn)換效率、使光合機(jī)構(gòu)和PSⅡ反應(yīng)中心受到損傷,從而導(dǎo)致枸杞光合作用效率下降。因此,氣溫升高加劇了干旱脅迫對寧夏枸杞葉片凈光合速率和水分利用效率的減小作用,即氣溫升高加劇了干旱脅迫對寧夏枸杞光合作用的影響。
[1] Houghton J T, Ding Y, Griggs D J, Noguer M, van der Linden P J, Dai X S, Maskell K, Johnson C A. Climate Change 2001: The Scientific Basis. Cambridge: Cambridge University Press, 2001.
[2] Karl T R, Knight R W, Baker B. The record breaking global temperatures of 1997 and 1998: evidence for an increase in the rate of global warming. Geophysical Research Letters, 2000, 27(5): 719-722.
[3] Rind D. Complexity and climate. Science, 1999, 284(5411): 105-107.
[4] Houghton J T, Jenkins G J, Ephraums J J. Climate Change: the IPCC Scientific Assessment. Cambridge: Cambridge University Press, 1990.
[5] 周平. 全球氣候變化對我國農(nóng)業(yè)生產(chǎn)的可能影響與對策. 云南農(nóng)業(yè)大學(xué)學(xué)報(bào), 2001, 16(1): 1-4.
[6] 王亞軍, 安巍, 石志剛, 趙建華. 枸杞藥用價(jià)值的研究進(jìn)展. 安徽農(nóng)業(yè)科學(xué), 2008, 36(30): 13213-13214, 13218-13218.
[7] 張?jiān)葡? 王萍, 劉敦華. 枸杞活性成分的研究進(jìn)展. 農(nóng)業(yè)科學(xué)研究, 2008, 29(2): 79-83.
[8] 安沙舟, 劉曉媛. 枸杞. 北京: 中國中醫(yī)藥出版社, 2001: 105-108.
[9] 萬佳, 延軍平. 寧夏近51年氣候變化特征分析. 資源開發(fā)與市場, 2012, 28(6): 511-514.
[10] 鄭廣芬, 陳曉光, 孫銀川, 張智, 納麗. 寧夏氣溫、降水、蒸發(fā)的變化及其對氣候變暖的響應(yīng). 氣象科學(xué), 2006, 26(4): 412-421.
[11] 陳曉光, 蘇占勝, 陳曉娟. 全球氣候變暖與寧夏氣候變化及其影響. 寧夏工程技術(shù), 2005, 4(4): 301-304.
[12] 陳豫英, 陳楠, 鄭廣芬, 穆建華, 馬篩艷, 納麗, 邵建. 近45a寧夏氣溫、降水及植被指數(shù)的變化分析. 自然資源學(xué)報(bào), 2008, 23(4): 626-634.
[13] 潘瑞熾. 植物生理學(xué) (第五版). 北京: 高等教育出版社, 2004: 56-57, 84-86.
[14] 呼和牧仁, 周梅, 翟洪波, 李良, 邵仁旭. 影響樹木光合作用因素的研究進(jìn)展. 內(nèi)蒙古農(nóng)業(yè)大學(xué)學(xué)報(bào): 自然科學(xué)版, 2009, 30(2): 287-291.
[15] 侯晶東, 曹兵, 宋麗華. CO2濃度倍增對寧夏枸杞光合特性的影響. 南京林業(yè)大學(xué)學(xué)報(bào): 自然科學(xué)版, 2012, 36(5): 71-76.
[16] 李超. 不同苜蓿品種葉綠素?zé)晒饧八稚硖匦缘谋容^研究 [D]. 北京: 中國農(nóng)業(yè)科學(xué)院, 2012.
[17] 孫景寬, 張文輝, 陸兆華, 劉新成. 干旱脅迫下沙棗和孩兒拳頭葉綠素?zé)晒馓匦匝芯? 植物研究, 2009, 29(2): 216-223.
[18] 李尚中, 樊廷錄, 王勇, 趙剛, 王磊, 唐小明, 黨翼, 趙暉. 不同覆膜集雨種植方式對旱地玉米葉綠素?zé)晒馓匦浴a(chǎn)量和水分利用效率的影響. 應(yīng)用生態(tài)學(xué)報(bào), 2014, 25(2): 458-466.
[19] 梁新華, 許興, 徐兆楨, 裘志新, 時(shí)海娟. 干旱對春小麥旗葉葉綠素a熒光動(dòng)力學(xué)特征及產(chǎn)量間關(guān)系的影響. 干旱地區(qū)農(nóng)業(yè)研究, 2001, 19(3): 72-77.
[20] 張守仁. 葉綠素?zé)晒鈩?dòng)力學(xué)參數(shù)的意義及討論. 植物學(xué)通報(bào), 1999, 16(4): 444-448.
[21] 王宇超, 王得祥, 彭少兵, 何帆. 干旱脅迫對木本濱藜生理特性的影響. 林業(yè)科學(xué), 2010, 46(1): 61-67.
[22] 云建英, 楊甲定, 趙哈林. 干旱和高溫對植物光合作用的影響機(jī)制研究進(jìn)展. 西北植物學(xué)報(bào), 2006, 26(3): 641-648.
[23] 姚慶群, 謝貴水. 干旱脅迫下光合作用的氣孔與非氣孔限制. 熱帶農(nóng)業(yè)科學(xué), 2005, 25(4): 80-85.
[24] 付士磊, 周永斌, 何興元, 陳瑋. 干旱脅迫對楊樹光合生理指標(biāo)的影響. 應(yīng)用生態(tài)學(xué)報(bào), 2006, 17(11): 2016-2019.
[25] 蘇培璽, 嚴(yán)巧娣. C4荒漠植物梭梭和沙拐棗在不同水分條件下的光合作用特征. 生態(tài)學(xué)報(bào), 2006, 26(1): 75-82.
[26] 杜國棟, 呂德國, 趙玲, 王素素, 蔡倩. 高溫對仁用杏光合特性及PSⅡ光化學(xué)活性的影響. 應(yīng)用生態(tài)學(xué)報(bào), 2011, 22(3): 701-706.
[27] Crafts-Brandner S J, Law R D. Effect of heat stress on the inhibition and recovery of the ribulose-1, 5-bisphosphate carboxylase / oxygenase activation state. Planta, 2000, 212(1): 67-74.
[28] Demmig B, Winter K, Krüger A, Czygan F C. Photoinhibition and zeaxanthin formation in intact leaves. Plant Physiology, 1987, 84(2): 218-224.
[29] Dodd I C, Critchley C, Woodall G S, Stewart G R. Photoinhibition in differently coloured juvenile leaves ofSyzygiumspecies. Journal of Experimental Botany, 1998, 49(325): 1437-1445.
[30] 付春霞, 張?jiān)? 王衍安, 范曉丹, 閆玉靜, 張友朋. 缺鋅脅迫對蘋果葉片光合速率及葉綠素?zé)晒馓匦缘挠绊? 中國農(nóng)業(yè)科學(xué), 2013, 46(18): 3826-3833.
[31] 蒲光蘭, 周蘭英, 胡學(xué)華, 鄧家林, 劉永紅, 肖千文. 干旱脅迫對金太陽杏葉綠素?zé)晒鈩?dòng)力學(xué)參數(shù)的影響. 干旱地區(qū)農(nóng)業(yè)研究, 2005, 23(3): 44-48.
[32] Qiu B S, Zhang A H, Liu Z L, Gao K S. Studies on the photosynthesis of the terrestrial cyanobacteriumNostocflagelliformesubjected to desiccation and subsequent rehydration. Phycologia, 2004, 43(5): 521-528.
Effects of elevated temperature and drought stress on photosynthesis ofLyciumbarbarum
ZHAO Qin1, PAN Jing2, CAO Bing1,*, SONG Lihua1
1SchoolofAgriculture,NingxiaUniversity,Yinchuan750021,China2CollegeofPreventionandControlofDesertification,Yongning750105,China
Climate change, mainly caused by elevated atmospheric CO2concentrations, affects plant growth and physiology, and much attention has been paid to this subject recently. Global warming will affect the quantity and geographical distribution of precipitation. In western China, water and shortages in precipitation are key factors for plant growth and survival. Global warming will result in increased evaporation of water in soil, which will cause some areas to experience more severe droughts. High air temperatures increase drought stress in plants, which in turn accelerates the damage caused by high temperatures. For these reasons, the physiological and morphological responses of plants to global warming have become a critical issue.Lyciumbarbarumis an economic forest tree species in Ningxia, China, and is unique because of the high quality of its fruit, which contains nutrients and microelements, especially polysaccharides, taurine, and carotenoids.Lyciumbarbarumshows resistance to drought, saline and alkaline soils, and to cold temperatures; furthermore, it has the ability to adapt to a wide range of ecological conditions, which makes it a significant ecological, social, and economic asset for the Ningxia region.Lyciumbarbarumhas been widely planted and has become one of the major agricultural crops in Ningxia. However, the response ofL.barbarumto elevated temperatures and water shortages under climate change remain unknown. Here, we hypothesized that the stressors resulting from elevated temperature and drought would not affect photosynthesis inL.barbarum. We subjected 1-y-oldL.barbarumseedlings to the following controlled conditions: open-top chambers to simulate different temperature conditions (ambient temperature, AT; elevated temperature, ET = AT+2.5—3.7 ℃); a combination of three different soil water contents (control group, W1, approximately 70%—75% of maximum soil water content; moderate drought stress, W2, approximately 50%—55% of maximum soil water content; and severe drought stress, W3, approximately 35%—40% of maximum soil water content). We then studied the effects of elevated temperature and drought stress on photosynthesis ofL.barbarumby testing the photosynthetic and fluorescence indices of seedlings. Our results showed that under elevated temperature conditions, net photosynthetic rates of seedlings in the moderate and severe drought stress treatments were reduced by 17.5% and 48.9%, respectively, that average stomatal conductance was reduced by 3.9%, and that water use efficiency was 57.8% that of the control group. Therefore, elevated temperature and soil drought stress reduced net photosynthetic rates, stomatal conductance, and intercellular CO2concentrations of the seedlings and increased transpiration rates, which reduced water utilization efficiency in the seedlings. The elevated temperature and soil drought stress reduced the optical energy transfer efficiency of the Photosystem Ⅱ activity center in leaves, which resulted in the reduced photosynthetic efficiency inL.barbarumseedlings. These results indicate that elevated temperature would increase the negative effects of drought stress on the net photosynthetic rate in the seedlings. In summary, the elevated temperature and drought stress reduced photosynthesis inL.barbarum.
elevated temperature; drought stress;Lyciumbarbarum; photosynthesis; chlorophyll fluorescence
教育部新世紀(jì)優(yōu)秀人才支持計(jì)劃項(xiàng)目(NCET-11-1022)
2014-01-09;
日期:2014-11-19
10.5846/stxb201401090073
*通訊作者Corresponding author.E-mail: bingcao2006@126.com
趙琴, 潘靜, 曹兵, 宋麗華.氣溫升高與干旱脅迫對寧夏枸杞光合作用的影響.生態(tài)學(xué)報(bào),2015,35(18):6016-6022.
Zhao Q, Pan J, Cao B, Song L H.Effects of elevated temperature and drought stress on photosynthesis ofLyciumbarbarum.Acta Ecologica Sinica,2015,35(18):6016-6022.
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學(xué)學(xué)報(bào)(自然科學(xué)版)(2021年1期)2021-06-09 08:43:00
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀(jì)智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
