不同氣候變化情境下中國木靈蘚屬和蓑蘚屬植物的潛在分布格局
2015-01-19 07:06:35郭水良
生態學報 2015年19期
關鍵詞:植物
沈 陽,于 晶,郭水良
上海師范大學生命與環境科學學院,上海 200234
不同氣候變化情境下中國木靈蘚屬和蓑蘚屬植物的潛在分布格局
沈 陽,于 晶,郭水良*
上海師范大學生命與環境科學學院,上海 200234
蓑蘚屬(Macromitrium)和木靈蘚屬(Orthotrichum)是木靈蘚科(Orthotrichaceae)的兩個最大的屬,前者呈現泛熱帶分布式樣,后者呈偏溫帶性分布式樣。應用當前和基于RCP4.5(Medium-Low Representative Concentration Pathways)二氧化碳排放情景下2050和2070的11個生物氣候變量,以及木靈蘚屬(Orthotrichum)66個、蓑蘚屬(Macromitrium)131個國內分布記錄,應用最大熵模型(MaxEnt),預測了氣候變暖背景下這兩屬植物在我國38個自然保護區潛在分布范圍的變化。11個生物氣候變量包括晝夜溫差月均值、年溫度變化范圍、溫度季節變化、最熱月份最高溫、最干季度平均溫度和最冷季度平均溫度、最濕月份雨量、最干月份雨量、雨量變化方差和最暖季度平均雨量。選擇了10%的遺漏錯誤對應的累積值作為分布預測閾值,為了更好地展示氣候變化下這兩個屬的潛在分布區變化,選擇了我國境內的38個自然保護區并計算了不同氣候條件下兩種蘚類植物屬在這些自然保護區的綜合氣候適應指數。38個自然保護區包括福建武夷山、甘肅祁連山、廣東南嶺、廣西花坪、廣西十萬大山、貴州梵凈山、貴州雷公山、海南尖峰嶺、河北五臺山、河南雞公山、河南小秦嶺、湖北神農架、湖北星斗山、吉林長白山、江西廬山、遼寧白石砬子、遼寧醫巫閭山、內蒙古大黑山、寧夏賀蘭山、青海三江源、山東云臺山、山西歷山、山西蘆芽山、陜西佛坪、陜西太白山、四川貢嘎山、四川臥龍、云南哀牢山、云南大圍山、云南納板河、浙江鳳陽山、浙江古田山、浙江九龍山、浙江普陀山、浙江清涼峰、浙江天目山、浙江烏巖嶺和重慶大巴山。結果表明,隨著氣候變暖,蓑蘚屬和木靈蘚屬在我國的適生面積都將減少,蓑蘚屬植物在我國的潛在分布區面積從當前的31.5%下降到12.23%,木靈蘚屬的從當前的65.81%下降到44.94%,氣候變化對蓑蘚屬植物分布的影響要大于木靈蘚屬。對于蓑蘚屬植物,當前氣候條件下的38個自然保護區中,除了寧夏賀蘭山和甘肅祁連山外,其他的36個保護區都有適合于蓑蘚屬植物分布的氣候條件。到了2070,河南雞公山、山東云臺山、浙江普陀山、浙江九龍山、浙江古田山、內蒙古大黑山、山西蘆芽山、江西廬山、遼寧醫巫閭山、廣西花坪、遼寧白石砬子、河北五臺山、山西歷山、河南小秦嶺、陜西佛坪、吉林長白山、貴州梵凈山、貴州雷公山和福建武夷山這19個自然保護區不再或幾乎不再適合于蓑蘚屬的分布。對于木靈蘚屬植物,在當前的氣候條件下,38個自然保護區中,除了海南尖峰嶺保護區,其余的37個保護區均處于木靈蘚屬植物潛在分布范圍之內。與當前氣候條件相比,2070年38個保護區的氣候條件均變得不利于木靈蘚屬分布,其中廣西花坪、江西廬山、云南納板河、廣西十萬大山、河南雞公山、山東云臺山、浙江普陀山、浙江天目山、浙江古田山、浙江九龍山、福建武夷山、貴州梵凈山、遼寧醫巫閭山、河南小秦嶺和陜西佛坪等15個保護區已不在木靈蘚屬的潛在分布范圍。
蓑蘚屬;木靈蘚屬;MaxEnt;氣候變暖;地理分布;預測
全球氣候變暖將嚴重影響物種的生理生態特性和地理分布,多數物種地理分布格局將發生深刻地變化[1]。氣候變暖條件下,歐洲西南部比利牛斯山脈的高山和亞高山植被隨著21世紀氣候的變暖其潛在生境將強烈地減少[2]。Lyons等研究了氣候變暖對美國威斯康新溪流中50種魚類分布的影響,隨著氣候變暖,所研究的3種冷水魚類、16種涼水魚類和4種暖水魚類的分布區將縮小,另外有4種暖水魚類的分布區維持不變,其余的23種暖水魚類的分布區將擴大[3]。Hagen等研究發現,氣候變暖將使喜冷性蛾類趨于分布于更高的海拔[4]。
隨著氣候變化問題的加劇,人們也開始應用生物分布模型,結合地理信息系統來預測氣候變化下生物分布區變化[5],其中最大熵模型(Maximum Entropy Model,簡稱Maxent)應用得非常廣泛。例如,李明陽等預測了氣候變曖情況下外來有害生物美國南方松大小蠹在氣候變化下的生境變化[1]。于晶等[6]和麻亞鴻等[7]用這一方法來預測部分苔蘚植物的潛在分布區,胡理樂等應用運用Maxent軟件,基于21個環境因子,預測了不同氣候變化情景下21世紀50和80年代中國五味子分布范圍[8]。Kumar基于19個生物氣候變量,應用MaxEnt軟件預測發現,喜瑪拉雅錫金地區杜鵑花屬植物在氣候變暖條件下的分布區會劇烈地收縮[9]。
苔蘚植物結構簡單,表面積大,是變水植物,對溫度和降水的變化的響應更為敏感[10-11],全球氣候的變化肯定會影響到苔蘚植物的分布格局[12]。蓑蘚屬(Macromitrium)和木靈蘚屬(Orthotrichum)是蘚類植物中的兩個大屬[13-15],多數是樹附生類群,其中蓑蘚屬植物具有泛熱帶分布特點,木靈蘚屬有偏溫帶的分布式樣[14]。麻亞鴻等基于這兩個屬在我國當前的地理分布數據,應用MaxEnt和地理信息系統,預測了當前氣候條件下兩屬植物的潛在分布范圍[7]。
本文的目的是基于當前氣候數據和2050和2070的預測氣候數據,應用MaxEnt 3.3.2和ArcGIS 9.3軟件,根據木靈蘚屬和蓑蘚屬的現代地理分布信息,預測氣候變化下蓑蘚屬和木靈蘚屬在中國的地理分布格局的變化,通過這兩個屬的研究,使人們更好地認識氣候變化對不同氣候適應特點的苔蘚植物分布的影響。
1 方法
1.1 數據來源
地圖資料:ArcGIS 9.3中需要的地圖數據從國家基礎地理信息系統 (http://nfgis.nsdi.gov.cn/) 下載的1∶400萬的中國行政區劃。
分布數據:蓑蘚屬和木靈蘚屬是分類上的困難類群。在野外,它們在“屬”這一級上的分別鑒別相對容易,但是鑒定到種比較困難,該科植物相對權威的分類學專著“中國蘚類植物志”第五卷英文版2013年才出版,因此在此之前的分類鑒定中肯定錯誤不少。考慮到這兩屬植物是比較穩定的分類單位,同一屬植物往往有相對穩定的地理分布格局,因此本文選擇屬的水平上開展預測研究。蓑蘚屬和木靈蘚屬在中國的地理分布見麻亞鴻等的文獻[7]。
氣候數據:從世界氣候數據網站下載當前條件1950—2000年、2050年、2070年3個時間段上的19個生物氣象數據變量(http://www.worldclim.org),數據的空間分辨率為2—5 arc-minutes, 即每一柵格大致相當于22 km2范圍。2050和2070年的數據為北京氣候中心發展的氣候系統模式1.1版本(BCC-CSM1.1)基于RCP45排放模型下模擬得到的。RCP45假設了一個全球協調一致的緩解舉措,2100年預計CO2濃度高達542 μL/L狀態,這是一個中等標準的預期排放量[16-17]。為了避免高度相關的氣候數據對預測結果的影響,計算了中國境內19個氣候因子的相關性,選擇相關系數小于0.7的因子參與分布區預測,本文中選擇了11個氣候變量(表1)。

表1 參與木靈蘚屬和蓑蘚屬植物在中國分布區預測的11個氣候因子
1.2 軟件來源
從國家基礎地理信息系統網站 (http://nfgis.nsdi.gov.cn/)下載中國地圖。ArcGIS 9.3 為美國ESRI公司開發的地理信息系統平臺,MaxEnt 3.3.2下載自普林斯頓大學網站(www.cs.princeton.edu/~schapire/MaxEnt/)。
1.3 預測方法
首先在MaxEnt的samples窗口導入植物的地理分布數據,在Environmental layer窗口中導入當前的11個氣候變量,在Projection layers directory窗口中導入2050年(或2070年)的11個相應氣候變量。選擇了啟發式估測和刀切法,設置了5次重復,產生了5個預測隨機模型,選擇平均情況下的模型進行預測。
隨機選取25%的分布點作為測試集,剩余的作為訓練集,其它參數均為軟件默認值,輸出格式為ASCII柵格圖層,最后導入GIS軟件,得到不同氣候條件下蓑蘚屬植物在中國潛在分布范圍的預測圖層,并執行重分類程序進行10個等級分類,分別輸出當前和2050年(或2070)時蓑蘚屬植物在中國范圍內的分布區預測重分類后的Raster文件。
目前還沒有一個廣泛接受的方法決定Maxent預測的分布閾值,即在此閾值上為預測種有可能分布,而此閾值下則不可能分布[18]。本文選擇了10%的遺漏錯誤(omission error, 即預測結果中,有10%的分布點不在預測范圍內)為閾值,進一步生成閾值以上的蓑蘚屬植物在當前和氣候變化下的分布預測圖。
總之,要實現貧困地區普通高校教師隊伍的穩定,避免過度的“一元與單項”性流動,除了從國家、學校和個人的角度出臺制定相關的制度外,還要根據形勢的發展和社會的進步不斷探索最新的引人用人留人制度,并嚴格恪守誠信,積極促進人才的成長與發展,真正從根源上穩住教師隊伍,從而實現我國高等教育的均衡發展,促進高等教育快速健康發展。
同樣方法獲取木靈蘚屬植物在不同氣候條件下的分布預測圖。
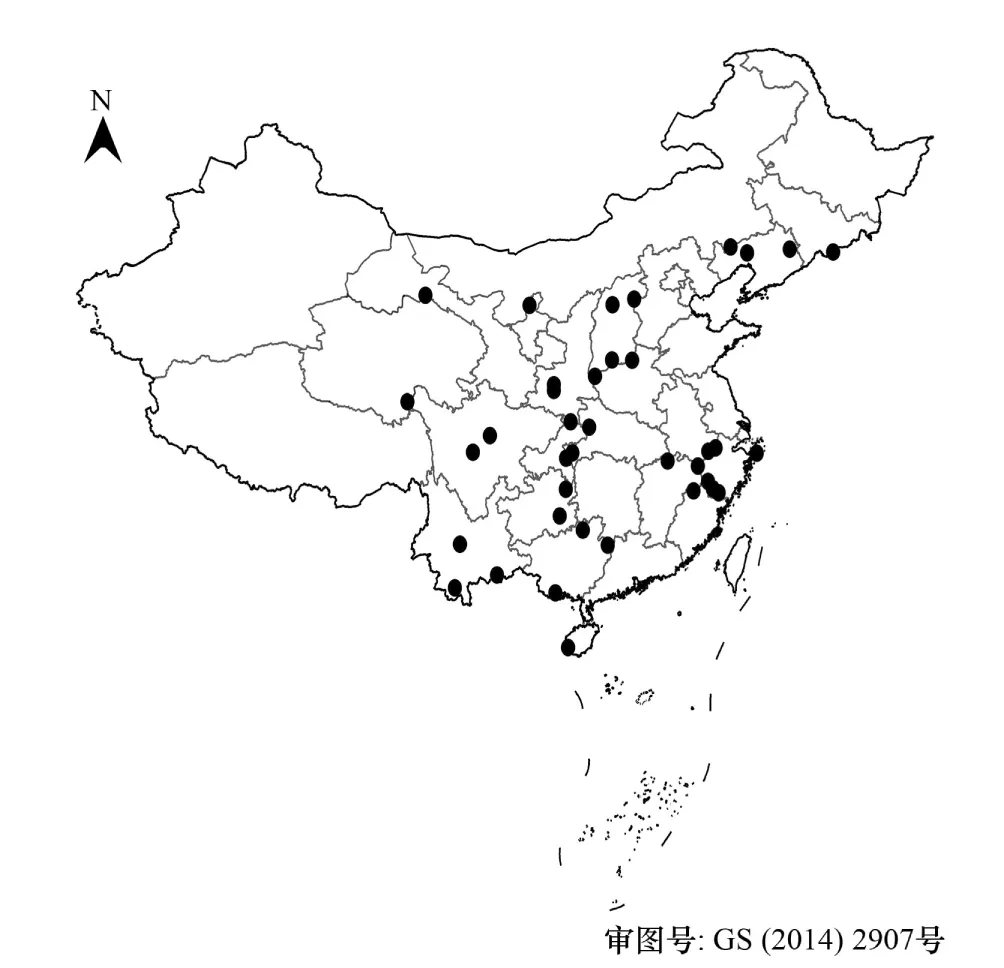
圖1 中國38個自然保護區的分布圖Fig.1 Locations of 38 reserves in China
為了更精細地反映蓑蘚屬和木靈蘚屬在不同氣候條件下在我國的潛在分布區變化情況,在我國不同地區選擇了38個自然保護區,預測這兩個屬植物在當前和氣候變暖條件下在這些保護區的潛在分布。這些保護區分別位于福建的武夷山、甘肅祁連山、廣東南嶺、廣西的花坪和十萬大山、貴州梵凈山和雷公山、海南的尖峰嶺、河北的五臺山、河南雞公山和小秦嶺、湖北神農架和星斗山、吉林長白山、江西廬山、遼寧白石砬子和醫巫閭山、內蒙古大黑山、寧夏賀蘭山、青海三江源、山東云臺山、山西歷山和蘆芽山、陜西佛坪和太白山、四川貢嘎山和臥龍、云南哀牢山、大圍山和納板河、浙江的鳳陽山、古田山、九龍山、普陀山、清涼峰、西天目山和烏巖嶺,和重慶大巴山(圖1)。
首先查取38個自然保護區的經度和緯度范圍,按相應保護區的經緯度范圍,作出封閉的長方形范圍圖代表這些保護區范圍,應用ArcGIS 9.3,輸出這些保護區的 shp格式地圖,再基于重分類后的預測文件,獲取不同氣候條件下蓑蘚屬和木靈蘚屬在相應保護區內各個預測等級下的面積百分數,進一步計算相應條件下的氣候適宜性指數[6]。
2 結果與分析
對蓑蘚屬和木靈蘚屬氣候適宜性預測的平均AUC值 (Area Under roc Curve)分別為0.859和0.782, 表明預測效果可以接受。蓑蘚屬和木靈蘚屬在我國當前、2050和2070年氣候條件下分布區預測結果分別見圖2和圖3。
當前氣候條件下,蓑蘚屬在臺灣山脈、黃山—天目山山脈、武夷山山脈、浙東—浙南山地、大別山、秦嶺—大巴山、雪峰山、南嶺等有很高的氣候適應性 (圖2);木靈蘚屬的潛在分布范圍比蓑蘚屬更廣(圖3)。與蓑蘚屬植物相比,木靈蘚屬更適合于偏北和更干燥的環境,特別是在華北、東北、西北地區和西南地區有較廣泛的分布,而在華南地區的分布行范圍小。
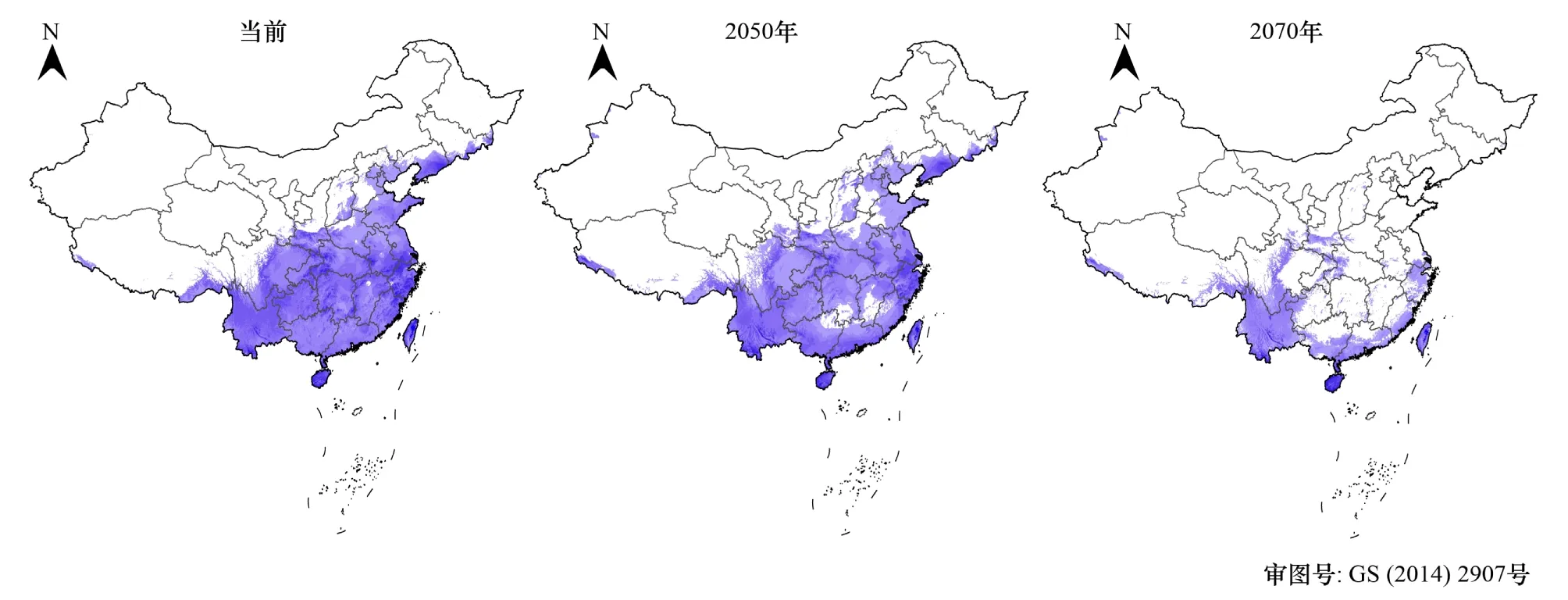
圖2 蓑蘚屬在當前、2050和2070氣候條件下在中國的分布區預測Fig.2 Predicted distribution regions of Macromitrium in China under current, 2050 and 2070 weather conditions陰影部分為氣候適應閾值以上的預測區域
根據本文標準,蓑蘚屬植物在我國的氣候適應閾值為0.23。38個自然保護區中,寧夏賀蘭山和甘肅祁連山的氣候適應指數在0.23以下,表明當前氣候條件下這兩個地區不在蓑蘚屬落在分布范圍之內,其他的36個保護區都有適合于蓑蘚屬分布的氣候條件。蓑蘚屬植物在浙江鳳陽山、海南尖峰嶺、浙江天目山、浙江清涼峰、遼寧白石砬子、浙江烏巖嶺、湖北星斗山、湖北神農架和云南納板河自然保護區的氣候適應指數最高,它們是蓑蘚屬最適宜分布的自然保護區。
2050年氣候條件下,38個地區中有12個地區(包括云南納板河、四川貢嘎山、廣西十萬大山、河北五臺山、山西蘆芽山、吉林長白山、云南哀牢山、青海三江源、四川臥龍、內蒙古大黑山、云南大圍山和甘肅祁連山)的蓑蘚屬植物氣候適應綜合指數高于當前氣候條件(表2和圖4),說明這些區域總體上變得更適應于蓑蘚屬的分布;其他的26個地區的氣候總體上變得不適合于蓑蘚屬植物的分布。如果按最高的氣候適應指數來分析,當前環境下,甘肅祁連山和寧夏賀蘭山不在蓑蘚屬的潛在分布范圍;到了2050年,除了以上兩個地區外,廣西花坪和內蒙古大黑山的氣候條件也不再適合于蓑蘚屬植物的分布。
按最合適氣候指數,到了2070年,除三江源自然保護區外,其余的37個自然保護區的蓑蘚屬氣候適應指數均比現在的下降。其中,河南雞公山、山東云臺山、浙江普陀山、浙江九龍山、浙江古田山、內蒙古大黑山、山西蘆芽山、江西廬山、寧夏賀蘭山、遼寧醫巫閭山、廣西花坪和遼寧白石砬子、河北五臺山、山西歷山、河南小秦嶺、陜西佛坪、吉林長白山、貴州梵凈山、貴州雷公山和福建武夷山這19個自然保護區2070年的氣候不再或幾乎不再適合于蓑蘚屬植物的分布,其余的18個自然保護區雖然氣候條件變差,但是仍然適合于蓑蘚屬植物的分布(表2)。
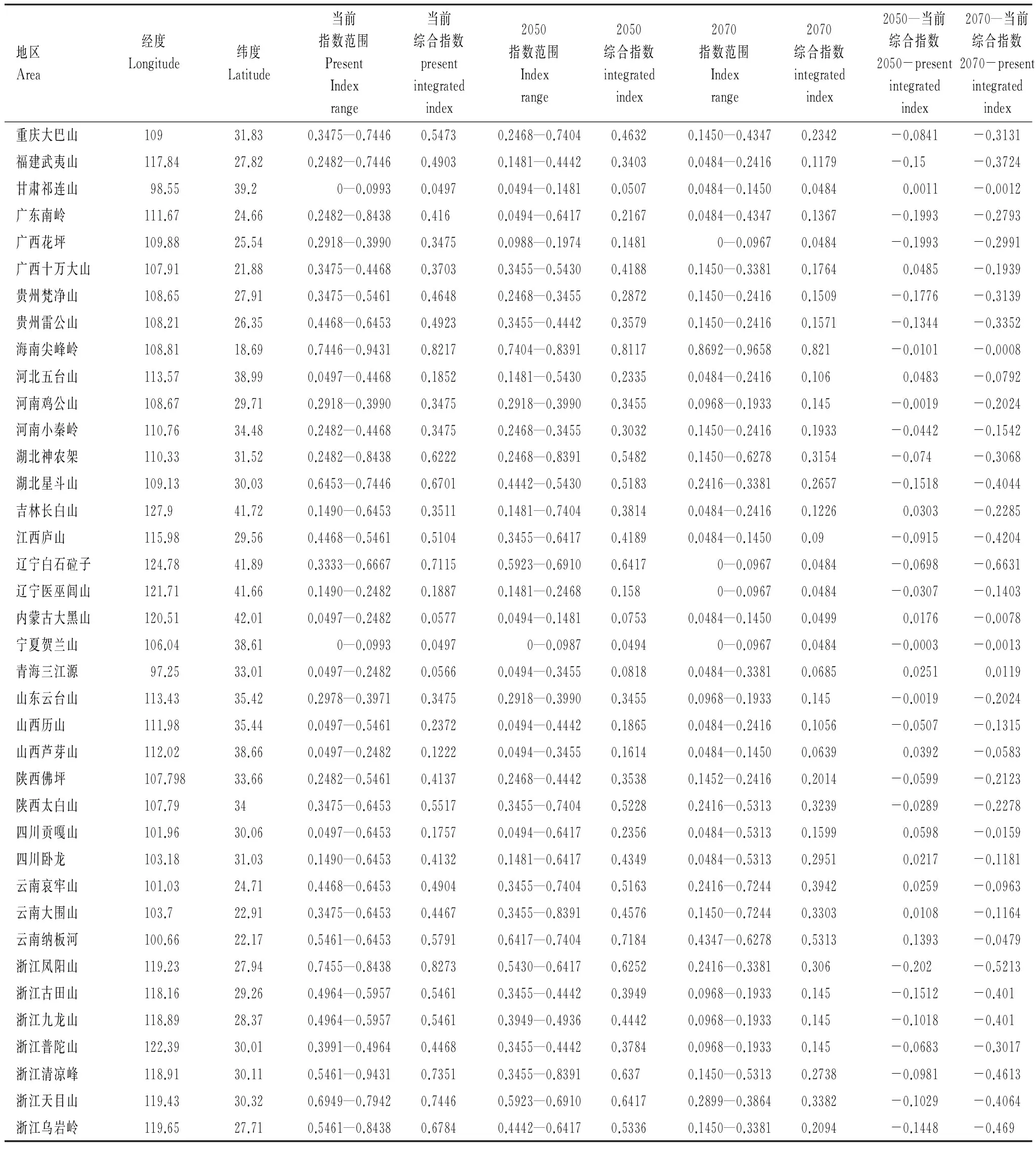
表2 當前、2050年和2070年三個時間點上蓑蘚屬植物在38個自然保護區的氣候適應指數及其變化
圖2直觀地展示了隨著氣候變暖蓑蘚屬植物氣候適應閾值以上的潛在分布區變化情況。很明顯,隨著氣候的變暖,蓑蘚屬植物的潛在適宜分布區會萎縮,最適范圍由當前的華東、華南、西南、華中和東北的遼東半島等區域縮小至主要在華南和西南的部分地區。就全國范圍來講,當前2050、2070年蓑蘚屬的潛在分布區面積百分比分別為31.21%、31.5%和12.23%,即從當前氣候到2050年,蓑蘚屬的潛在分布區面積并無明顯變化,且部分自然保護區的氣候條件更適合于蓑蘚屬植物的分布,但是從2050年到2070年間,蓑蘚屬植物的潛在分布區面積從31.5%下降到12.23%,期間幾乎所有的38個自然保護區的氣候條件變更不適合于蓑蘚屬植物的分布。全國范圍來講,隨著氣候變化,蓑蘚屬的綜合氣候適應指數持續地變小,當前、2050和2070狀態下分別為0.1715、0.1593和0.0995。
在當前的氣候條件下,木靈蘚屬植物在38個自然保護區中,除了海南尖峰嶺保護區,其余的保護區均處于木靈蘚屬植物潛在分布范圍之內(圖3和表3)。
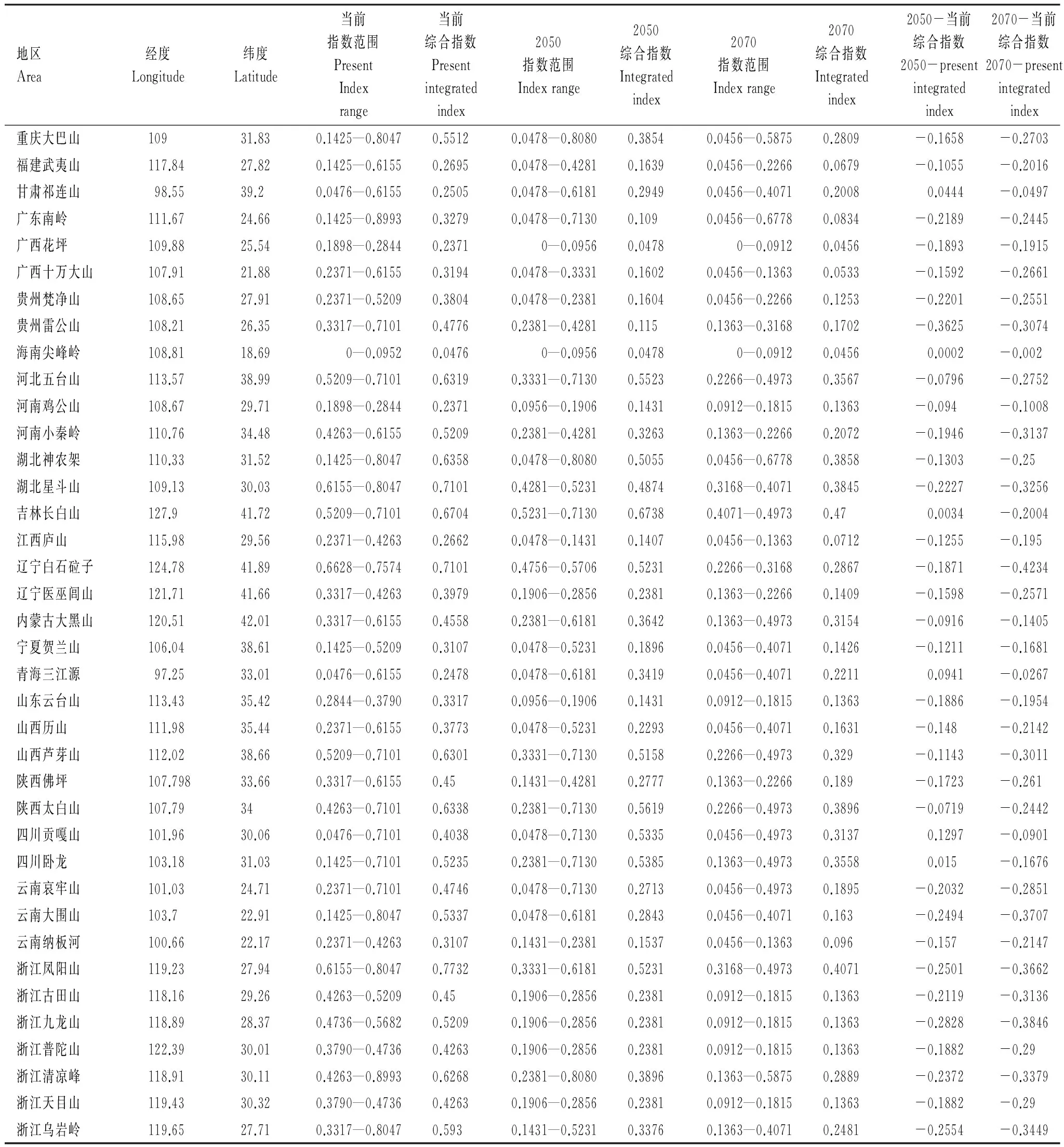
表3 當前、2050年和2070年三個時間點上木靈蘚屬植物在38個自然保護區的氣候適應指數及其變化
從當前狀態到2050年,海南尖峰嶺、吉林長白山、四川臥龍、甘肅祁連山、青海三江源和四川貢嘎山的木靈蘚屬氣候適應指數變高,其余的32個保護區的氣候適應指數均變低,反映氣候條件在惡化。廣西花坪、江西廬山、山東云臺山、河南雞公山、貴州梵凈山和云南納板河等目前處于木靈蘚屬的潛在分布范圍,但是到了2050年它們的氣候條件不再或幾乎不再適合于木靈蘚屬的分布(圖3和表3)。
2070年狀態下38個保護區的木靈蘚屬綜合氣候適應指數均比當前狀態的小。廣西花坪、江西廬山、云南納板河、廣西十萬大山、河南雞公山、山東云臺山、浙江普陀山、浙江天目山、浙江古田山、浙江九龍山、福建武夷山、貴州梵凈山、遼寧醫巫閭山、陜西佛坪、河南小秦嶺等15個地區當前處在木靈蘚屬的氣候適應范圍內,到2070年均變得不適合木靈蘚屬植物的分布(表3)。對于木靈蘚屬來講,隨著氣候的變暖,木靈蘚屬的氣候適應范圍持續變小,該屬的潛在分布區面積從當前的65.81%下降到2050年的50.06%,到2070年進一步下降到44.94%,綜合氣候適應指數也從當前的0.2933下降到2050年的0.2366,到2070年進一步下降到0.1928。
蓑蘚屬植物與木靈蘚屬植物的現代地理分布格局存在明顯差異。蓑蘚屬植物偏溫暖潮濕分布,在我國的東部和東南部比木靈蘚屬的氣候適應能力高些,而木靈蘚屬偏溫帶干旱的環境,在我國西北部、華東和東北有較高的分布概率(圖2,圖3)。總體上,木靈蘚屬比蓑蘚屬具有更廣的氣候適應范圍,更高的綜合氣候適應指數(圖2,圖3,表4)。
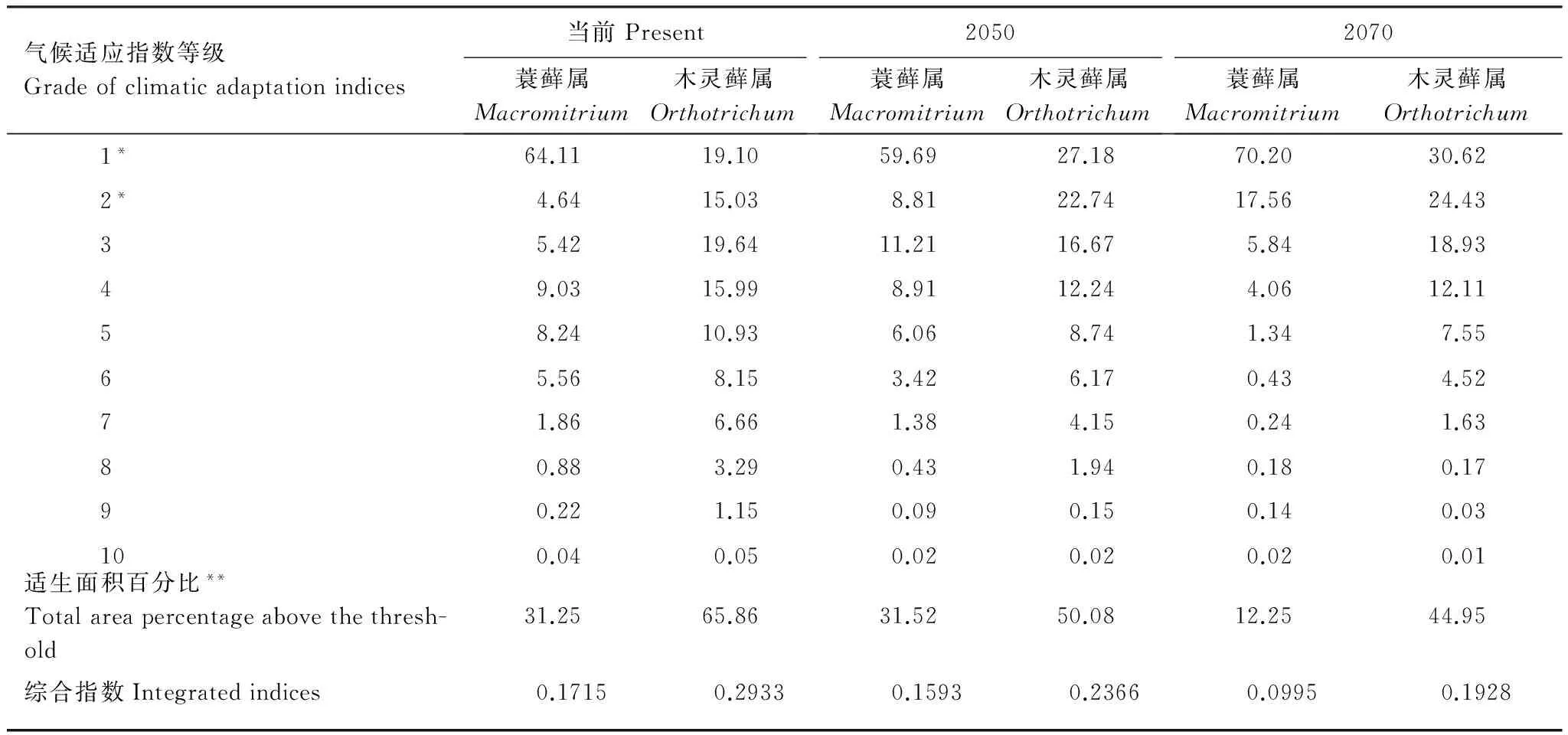
表4 當前、2050年和2070年氣候狀態下蓑蘚屬和木靈蘚屬在各氣候適應等級上在我國的面積比和綜合氣候適應指數
3 討論
本文利用MaxEnt模型對木靈蘚科兩個大屬在氣候變化下我國的適生區進行了預測,反映了氣候變化會強烈地影響到蘚類植物分布格局。
世界上約有30余種氣候模式使用不同的排放情景對未來氣候變化進行模擬預測,不同模式給出的結果差異很大[19]。由于設置CO2排放的情景不同,對于全球CO2的變化有3個典型的預測,即RCP2.6、RCP4.5和RCP8.5,在RCP2.6模型中,全球平均的CO2濃度將在2050年達到440 μL/L,在RCP8.5的模型中,CO2的濃度將達到935 μL/L,而且在2250年將達到1961 μL/L。本文中采用了中等程度的CO2增高下預測的氣候數據,以此數據預測蓑蘚屬和木靈蘚屬在中國的分布范圍,發現此強度的氣候變化下對蓑蘚屬和木靈蘚屬在中國的分布范圍已造成了巨大的影響。RCP8.5是假設全球的CO2排放按目前這種狀態,不進行專門的協調減排出現的情形,可以預計,如果在RCP8.5的排放情形下,我國的苔蘚植物分布格局將受到更強烈的影響,眾多溫帶性的種類會從中國蘚類植物區系中消失。
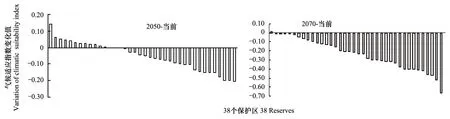
圖4 2050年和2070年氣候條件下蓑蘚屬在38個保護區的綜合氣候適應指數與當前的相比的變化Fig.4 Variation of integrated climatic adaptation indices of Macromitrium in 38 Reserves in 2050 and 2070 compared those under present climatic conditions 橫坐標上的38個保護區以變化值由正到負的程度排列
全球氣候變化是一個復雜的生物地球化學過程,這一過程中,年平均溫度的變化相對緩慢,而溫度的季節性變動系數上升的幅度更強烈,最濕季節的降水量在全球范圍會呈現出更加不均衡趨勢[1]。從本文的研究可以看出,溫度的季節性波動、最濕季節的降水量等對蓑蘚屬和木靈蘚屬植物會造成強烈影響。氣候變化下降水的時空不均衡分布和溫度季節性變動幅度的增大, 會對整個苔蘚植物的分布造成強烈影響。
生物分布的預測過程中使用的地理背景會對物種預測結果造成很大影響[20]。如果地理背景太小,不能夠代表預測類群整個分布范圍,會使該類群的預測范圍被低估[21]。Acevedo等應用8個不同大小的地理背景來預測4種生物的地理分布范圍,發現地理背景范圍緊鄰目前種類的分布范圍時預測效果最好[20]。本文中包括了蓑蘚屬的131個地理分布數據和木靈蘚屬的66個地理分布數據,覆蓋了這兩個屬在我國的分布范圍,結合AUC值,因此預測的結果可信。
應用MaxEnt進行的植物分布區預測中,人們多數應用從世界氣候數據網站下載的19個生物氣候數據變量(http://www.worldclim.org), 這19個氣候數據變量是根據12個月中溫度和雨量記錄演生而來[22]。但是,Kruijer等認為高度相關的變量會影響預測結果,即19個變量之間存在多重線性重復(multicollinearity)的問題[23],為了避免這一問題,建議采用泊松相關系數在0.7到-0.7之間的氣候變量,本文也采用了這一策略。 不過,在今后基于生物氣候數據,應用MaxEnt進行物種分布區預測時,建議采取不同的氣候變量或變量組合,借助于現在快速運算能力的計算機,取AUC最高的方案,而所需要變量又最少的標準選擇預測模型。
Maxent 模型是一種基于生態位原理的生境適宜性模型,與其他的物種模型相比,具有較高的預測精度,在物種分布數據較少的情況下依然會取得較高的預測精度,因此在當前的生物地理分布范圍預測中得到了廣泛的應用[24]。但是,對于象木靈蘚屬和蓑蘚屬來講,僅僅考慮到氣候因素也會一定程度上影響物種分布區的預測精度,因為這兩個屬主要是樹附生的。因此,將來的研究工作中,如果將植被數據引入到它們的分布預測中,會提高預測精度。本文主要考慮到需要預測氣候變化下兩種的分布范圍,缺乏在氣候變化情況下全球植被的分布數據,因此僅僅考慮了11種生物氣候數據。
此外,Guralnick基于歷史氣候和物種分布資料的研究發現,隨著氣候變暖,那些位于分布區南緣的山地型哺乳動物會向高海拔的區域分布,而平地型哺乳動物則向高緯度方向分布,并認為基于最近一次冰期后的氣候變暖的物種分布區響應能夠更好地預測今后氣候變暖對物種分布的影響[25]。但是這些方法需要有比較好的化石記錄,由于苔蘚植物缺少足夠多的化石資料,難于使用這類方法用于苔蘚植物分布區的預測。
[1] 李明陽, 巨云為, 吳文浩, 何燕杰, 徐光彩. 氣候變化情景下外來森林病蟲害潛在生境動態分析——以美國南方松大小蠹為例. 北京林業大學學報, 2009, 31(4): 64-69.
[2 ] Pérez-García N, Font X, Ferré A, Carreras J. Drastic reduction in the potential habitats for alpine and subalpine vegetation in the Pyrenees due to twenty-first-century climate change. Regional Environmental Change, 2013, 13(6): 1157-1169.
[3] Lyons J, Stewart J S, Mitro M. Predicted effects of climate warming on the distribution of 50 stream fishes in Wisconsin, U. S. A. Journal of Fish Biology, 2010, 77(8): 1867-1898.
[4] Hagen S B, Jepsen J U, Ims R A,Yoccoz N G. Shifting altitudinal distribution of outbreak zones of winter mothOperophterabrumatain sub-arctic birch forest: a response to recent climate warming?. Ecography: Pattern and Diversity in Ecology, 2007, 30(2): 299-307.
[5] Porretta D, Mastrantonio V, Amendolia S, Gaiarsa S, Epis S, Genchi C, Bandi C, Otranto D, Urbanelli S. Effects of global changes on the climatic niche of the tickIxodesricinusinferred by species distribution modeling. Parasites & Vectors, 2013, 6(1): 1-8.
[6] Yu J, Ma Y H, Guo S L. Modeling potential geographic distribution of epiphytic mossMacromitriumjaponicum(Bryophyta: Musci) in China via a Maximum Entropy Algorithm and ArcGIS. Annuales Botanici Fennici, 2013, 50(1/2): 35-42.
[7] 麻亞鴻, 李丹丹, 于晶, 郭水良. 中國蓑蘚屬與木靈蘚屬分布式樣與氣候因子的關系. 生物多樣性, 2013, 21(2): 177-184.
[8] 胡理樂, 張海英, 秦嶺, 閆伯前. 中國五味子分布范圍及氣候變化影響預測. 應用生態學報, 2012, 23(9): 2445-2450.
[9] Kumar P. Assessment of impact of climate change onRhododendronsin Sikkim Himalayas using Maxent modelling: limitations and challenges. Biodiversity Conservation, 2012, 21(5): 1251-1266.
[10] Bates J W, Thompson K, Grime P. Effects of simulated long termclimatic change on the bryophytes of a lime stone grass land community. Global Change Biology, 2005, 11(5): 757-769.
[11] Toet S, Cornelissen J H C, Aerts R, van Logtestijn R S P, de Beus M, Stoevelaar R. Moss responses to elevated CO2and variation in hydrology in a temperate lowland peatland. Plant Ecology, 2006, 182(1/2): 27-40.
[12] 孫守琴, 田維莉. 苔蘚植物對氣候變化的響應研究進展. 貴州師范大學: 自然科學版, 2010, 28(4): 103-107.
[13] Crosby M R, Magill R E, Allen B, He S. A checklist of the mosses. St. Louis, Missouri Botanical Garden, 1999: 1-306.
[14] Jia Y, He S, Guo S L. Orthotrichaceae // Wu P C, Crosby M R. Moss flora of China, English version, Erpodiaceae-Climaciaceae, 5, Science Press (Beijing, New York) & Missouri Botanical Garden Press (St. Louis), 2011, 22-116.
[15] Guo S L, Ma Y H, Cao T, Lou Y X. A synopsis ofMacromitrium(Orthotrichaceae) in China. Cryptogamie, Bryologie, 2012, 33(4): 341-355.
[16] Dufresne J L, Foujols M A, Denvil S, Caubel A, Marti O, Aumont O, Balkanski Y, Bekki S, Bellenger H, Benshila R, Bony S, Bopp L, Braconnot P, Brockmann P, Cadule P, Cheruy F, Codron F, Cozic A, Cugnet D, de Noblet N, Duvel J P, Ethé C, Fairhead L, Fichefet T, Flavoni S, Friedlingstein P, Grandpeix J Y, Guez L, Guilyardi E, Hauglustaine D, Hourdin F, Idelkadi A, Ghattas J, Joussaume S, Kageyama M, Krinner G, Labetoulle S, Lahellec A, Lefebvre M P, Lefevre F, Levy C, Li Z X, Lloyd J, Lott F, Madec G, Mancip M, Marchand M, Masson S, Meurdesoif Y, Mignot J, Musat I, Parouty S, Polcher J, Rio C, Schulz M, Swingedouw D, Szopa S, Talandier C, Terray P, Viovy N, Vuichard N. Climate change projections using the IPSL-CM5 Earth System Model: from CMIP3 to CMIP5. Climate Dynamics, 2013, 40(9/10): 2123-2165.
[17] 杭月荷. CMIP5多模式對中國極端降水的模擬評估及未來情景預估 [D]. 南京: 南京信息工程大學, 2013.
[18] Phillips S J, Anderson R P, Schapire R E. Maximum entropy modeling of species geographic distributions. Ecological Modelling, 2006, 190(3/4): 231-259.
[19] 呂佳佳, 吳建國. 氣候變化對植物及植被分布的影響研究進展. 環境科學與技術, 2009, 32(6): 85-95.
[20] Acevedo P, Jiménez-Valverde A, Lobo J M, Real R. Delimiting the geographical background in species distribution modeling. Journal of Biogeography, 2012, 39(8): 1383-1390.
[21] Barve N, Barve V, Jiménez-Valverdea A, Lira-Noriegaa A, Mahera S P, Peterson A T, Soberóna J, Villalobos F. The crucial role of the accessible area in ecological niche modeling and species distribution modeling. Ecological Modelling, 2011, 222(11): 1810-1819.
[22] Hijmans R J, Graham C H. The ability of climate envelope models to predict the effect of climate change on species distributions. Global Change Biology, 2006,12(2): 2272-2281.
[23] Kruijer J D, Raes N, Stech M. Modeling the distribution of the moss speciesHypopterygiumtamarisci(Hypopterygiaceae, Bryophyta) in Central and South America. Nova Hedwigia, 2010, 91(3/4): 399-420.
[24] Elith J, Graham C H, Anderson R P, Dudík M, Ferrier S, Guisan A, Hijmans R J, Huettmann F, Leathwick J R, Lehmann A, Li J, Lohmann L G, Loiselle B A, Manion G, Moritz C, Nakamura M, Nakazawa Y, Overton J M M, Peterson A T, Phillips S J, Richardson K, Scachetti-Pereira R, Schapire R E, Soberón J, Williams S, Wisz M S, Zimmermann N E. Novel methods improve prediction of species′ distributions from occurrence data. Ecography, 2006, 29(2): 129-151.
[25] Guralnick R. Differential effects of past climate warming on mountain and flatland species distributions: a multispecies North American mammal assessment. Global Ecology and Biogeography, 2007, 16(1): 14-23.
MacromitriumandOrthotrichumdistribution patterns under different climate warming scenarios in China
SHEN Yang, YU Jing, GUO Shuiliang
CollegeofLifeandEnvironmentalScience,ShanghaiNormalUniversity,Shanghai200234,China
MacromitriumandOrthotrichumare two largest genera in the Orthotrichaceae (Bryopsida) family.Macromitriumare distributed across tropical zones, whileOrthotrichumare distributed throughout temperate zones. Based on eleven bioclimatic variables modeled under recent (1950—2000), 2050, and 2070 RCP45CO2emission scenarios, 66 geographical records ofOrthotrichum, and 131 records ofMacromitriumin China, this paper used the algorithm of maximum entropy (Maxent) to predict the potential Chinese distributions of these two genera in 2050 and 2070. The eleven bioclimatic variables are: mean diurnal temperature range; temperature seasonality; annual temperature range; the highest temperature of the warmest month; mean temperatures of the driest quarter and coldest quarter; precipitations of the wettest month, driest month, warmest quarter, and coldest quarter; and precipitation seasonality. Thirty-eight nature reserves were selected and an integrated climate suitability index was calculated for each of them under different climate conditions. These 38 reserves are: Wuyishan (Fujian); Qilianshan (Gansu); Nanling (Guangdong); Huaping and Shiwandashan (Guangxi); Fanjingshan and Leigongshan (Guizhou); Jianfengling (Hainan); Wutaishan (Hebei); Jigongshan and Xiaoqinling (Henan); Shennongjia and Xingdoushan (Hubei); Changbaishan (Jilin); Lushan (Jiangxi); Baishilizi and Yiwulvshan (Liaoning); Daheishan (Inner Mongolia); Helanshan (Ningxia); Sanjiangyuan (Qinghai); Yuntaishan (Shandong); Lishan, Luyashan, Foping, and Taibaishan (Shaanxi); Gonggashan and Wolong (Sichuan); Ailaoshan, Daweishan, and Nabanhe (Yunnan); Fengyangshan, Gutianshan, Jiulongshan, Putuoshan, Qingliangfeng, Xitianmushan, and Wuyanling (Zhejiang); and Dabashan (Chongqing). The corresponding cumulative value for an omission error of 10% was selected as a distribution threshold. The primary results were: (1) The suitable bioclimatic habitats forMacromitriumandOrthotrichumwould shrink considerably with predicted climate warming. The potential distribution range of Macromitrium in China would declime from 31.5% under recent climate condition to 12.23% under 2070 climate condition, while that of Orthotrichum would declime from 65.81% to 44.94%. The negative effects of climatic warming on the potential distribution would be greater forMacromitriumthan forOrthotrichum; (2) All of the nature reserves, except for Helanshan and Qilianshan, had suitable niches forMacromitriumunder the recent climatic scenario. Under the 2070 predicted climate scenario, the potential distribution ofMacromitriumwould not include Jigongshan, Yuntaishan, Putuoshan, Jiulongshan, Gutianshan, Daheishan, Luyashan, Lushan, Yiwulvshan, Huaping, Baishilazi, Wutai, Lishan, Xiaoqinling, Foping, Changbaishan, Fanjingshan, Leigongshan, and Wuyishan (half of the 38 nature reserves in this study); and (3) All of the nature Reserves, except Jianfengling, had suitable niches forOrthotrichumunder the recent climate scenario. By 2070, the weather adaptation indices forOrthotrichumin all of the 38 reserves would be lower than those under the current conditions. According to our predictions for 2070,Orthotrichumwould be excluded from the reserves in Huaping, Lushan, Shiwandashan, Jigongshan, Taishan, Putuoshan, Xitianmushan, Gutianshan, Jiulongshan, Wuyishan, Fanjingshan, Yiwulvshan, Xiaoqinlingshan, and Foping.
Macromitrium;Orthotrichum; maxent; climate warming; geographical distribution; prediction
國家自然科學基金(31370233,30970184);上海市科委重點項目(12490502700); Bilateral cooperation project of University of Ostrava and Ostrava town, Czech republic (Taxonomical revision of ChineseOrthotrichum, no: 00955-RRC-2015)
2014-03-23;
2014-11-26
10.5846/stxb201403230523
*通訊作者Corresponding author.E-mail: guoshuiliang@163.com
沈陽,于晶,郭水良.不同氣候變化情境下中國木靈蘚屬和蓑蘚屬植物的潛在分布格局.生態學報,2015,35(19):6449-6459.
Shen Y, Yu J, Guo S L.MacromitriumandOrthotrichumdistribution patterns under different climate warming scenarios in China.Acta Ecologica Sinica,2015,35(19):6449-6459.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
