辣白菜中乳酸菌的分離鑒定
2015-01-03 03:40:36孟令帥鄒婷婷烏日娜岳喜慶武俊瑞
食品科學 2015年11期
孟令帥,張 穎,鄒婷婷,烏日娜,2,岳喜慶,武俊瑞,*
辣白菜中乳酸菌的分離鑒定
孟令帥1,張 穎1,鄒婷婷1,烏日娜1,2,岳喜慶1,武俊瑞1,*
(1.沈陽農業大學食品學院,遼寧 沈陽 110866;2.江南大學食品學院,食品科學與技術國家重點實驗室,江蘇 無錫 214122)
從沈陽市7 個區25 份朝鮮族家庭制作的傳統發酵辣白菜中分離出81 株乳酸菌疑似菌株,初步鑒定34 株為桿菌,47 株為球菌。進一步采用16S rDNA序列分析對81 株菌進行分子鑒定,通過序列分析進行屬種鑒定。結果表明:81 株菌均為乳酸菌,分別來自2 個屬6 個種,45 株為屎腸球菌,25 株為植物乳桿菌,4 株為干酪乳桿菌,3 株為戊糖乳桿菌,2 株為短乳桿菌,2 株為堅強腸球菌。研究結果為我國東北辣白菜中乳酸菌作進一步研究奠定了基礎。
辣白菜;乳酸菌;16S rDNA;同源性分析
辣白菜是韓國、朝鮮和我國東北地區特有的一種傳統發酵泡菜食品[1-3],在我國東北人民飲食中,作為一種非常重要的配菜,至今仍深受消費者喜愛。辣白菜的主要原料是大白菜,其制作方式是:先用鹽水將白菜腌制,低溫貯存幾天,然后,將辣椒、大蒜、生姜、魚汁、青梅汁和蜂蜜等配料混勻后,均勻地涂抹于白菜的表面,再將一些水果切片涂抹在白菜的表面,最后經過乳酸菌等微生物發酵而成[4-7]。近年來的研究表明,辣白菜中存在豐富的乳酸菌資源[8-9],是挖掘有益乳酸菌的寶庫。2012年,Kim等[10]研究發現,明串珠菌、乳酸桿菌、短乳桿菌以及鏈球菌有助于泡菜發酵,植物乳桿菌屬和片球菌屬會使泡菜過度酸化。
本實驗試圖通過傳統方法從采集到的25 份辣白菜樣品中分離出乳酸菌疑似菌株,并結合16S rDNA序列分析技術,分別對其鑒定,初步探明辣白菜中乳酸菌的構成。
1 材料與方法
1.1 材料、培養基與試劑
樣品分別采自沈陽市大東區、東陵區、皇姑區、和平區、鐵西區、于洪區以及蘇家屯區7 個區不同的農貿市場,共計25 份辣白菜樣品,所用樣品均為朝鮮族傳統發酵辣白菜。
培養基采用MRS培養基[11]。
20 mg/mL蛋白酶K、RNA酶、正向引物、反向引物、TaKaRa Ex Taq酶、10×PCR Buffer、dNTP Mix、瓊脂糖、溴化乙錠(ethidium bromide,EB)染色液北京鼎國昌盛生物科技有限公司。
1.2 儀器與設備
LDZX-50KBS立式壓力蒸汽滅菌鍋鄭州南北儀器設備有限公司;無菌操作臺蘇凈集團安泰公司;DNP-9080生化培養箱上海精密實驗設備有限公司;HH-6恒溫水浴鍋常州國華電器有限公司;VS-1渦旋儀北京鼎昊原料科技有限公司;CR-21G離心機(50 mL)天美科技儀器有限公司;Centrifuge-5418離心機(1.5 mL)上海艾研生物科技有限公司;PTC-200梯度基因擴增儀、瓊脂糖凝膠電泳儀美國伯樂公司;UV-3200分光光度計上海美普達儀器有限公司。
1.3 方法
應用層主要是以數據層為基礎,通過調用數據層向上提供的接口,對該層提供的盾構運行數據和特征數據進行對應的加工處理,實現相應的業務邏輯,并向表示層提供接口,將實現的業務邏輯通過表示層可視化表現。如此處于系統中間層的應用層是系統核心,起著承上啟下的作用,系統的功能實現則主要在這層實現。
1.3.1 辣白菜中乳酸菌活菌數的測定
采用平板計數法,分別對25 個辣白菜樣品中的乳酸菌活菌數進行測定[12-13]。
1.3.2 乳酸菌的分離、純化與初步鑒定
將采集的辣白菜汁樣品用滅菌生理鹽水進行梯度稀釋,分別接種到含有2%碳酸鈣的固體MRS培養基中,分別在20、37、45 ℃厭氧培養48 h[14],挑選有鈣圈的菌落,分別劃線分離純化2~3次后,選取單菌落,經革蘭氏染色后鏡檢,同時進行過氧化氫酶實驗,確認純種、革蘭氏陽性、過氧化氫酶陰性的無芽孢菌進行拍照保存[11-17]。
1.3.3 乳酸菌16S rDNA序列鑒定
采用十六烷基三甲基溴化銨(hexadecyltrimethy ammonium bromide,CTAB)法提取對數生長末期的乳酸菌基因組DNA[18-19],采用16S rDNA通用引物進行聚合酶鏈式反應(polymerase chain reaction,PCR)擴增[20-21],正向引物為27F(對應于Escherichia coil 8~27位堿基):5’-AGAGTTTGATCCTGGCTCAG-3’;反向引物為1495R(對應于Escherichia coil 1495~1515位堿基):5′-CTACGGCTACCTTGTTACGA-3′,由上海桑尼生物技術有限公司合成。擴增產物經1.0%的瓊脂糖凝膠電泳檢測,在1 500 bp處有清晰的條帶后測序[22-24],測序由上海桑尼生物技術有限公司進行。
1.3.4 乳酸菌同源性分析
利用BLAST分析工具(http://www.ncbi.nlm.nih.gov/ BLAST)將所分離的乳酸菌16S rDNA序列與GenBank/ EMBL/DDBJ數據庫中已知菌株的對應序列進行比較鑒定,尋找與目的基因序列同源性最高的已知菌種[25]。
2 結果與分析
2.1 辣白菜中乳酸菌計數結果

表1 辣白菜中乳酸菌計數結果Table 1 Lactic acid bacterial counts in kimchi
由表1可知,25 個辣白菜樣品中乳酸菌菌落總數大約在3.30×108~2.56×1010CFU/mL之間,平均為8.08×109CFU/mL。說明辣白菜樣品中乳酸菌含量較高,對于辣白菜的發酵可能發揮著重要的作用。
2.2 辣白菜中乳酸菌的分離與鑒定
2.2.1 辣白菜中乳酸菌的分離與初步鑒定
乳酸菌的形態學特征包括菌落形狀、菌落顏色和菌落透明度等,從25 個辣白菜樣品中初步分離出81 株疑似乳酸菌的菌株,經過革蘭氏染色和過氧化氫酶反應,81 株菌革蘭氏染色均為陽性,過氧化氫酶反應均為陰性。其中,革蘭氏染色結果為:桿菌34 株,球菌47 株。81 株菌經初步鑒定均為乳酸菌。部分代表型乳酸菌菌株的革蘭氏染色鏡檢照片如圖1所示。


圖1 部分代表型乳酸菌的革蘭氏染色圖片Fig.1 Gram staining photos of some representativeLactobacillusstrains
2.2.2 乳酸菌16S rDNA序列鑒定和同源性分析結果

表2 乳酸菌16S rDNA序列同源性分析結果Table 2 Homological analysis of lactic acid bacteria by 16S rDNA sequence
由表2可知,從沈陽各地采集的25 份朝鮮族家庭制作的辣白菜樣品中分離到的81 株乳酸菌疑似菌株,經鑒定均為乳酸菌,除菌株20HM1-2、37TX2-2、37HX2-1、37SJ1-6、20HX3-1和37HX7-1這6 株菌外,其余75 株菌與標準菌株的同源性均在99%以上,其中,56 株菌與標準菌株的同源性達到100%。將上述結果進行歸納后可得到表3。
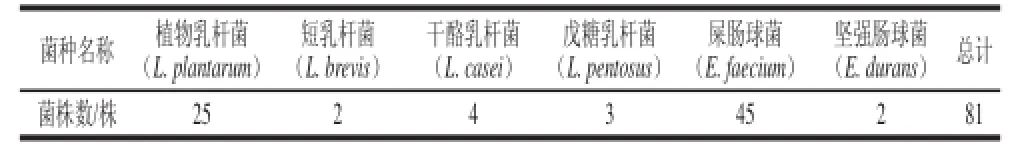
表3 辣白菜中乳酸菌的分布結果Table 3 Distribution of lactic acid bacteria in kimchi
由表3可知,25 份樣品中分離出的81 株乳酸菌疑似菌株,經16S rDNA序列測定和同源性分析鑒定,分屬2 個屬6 個種,分別為E. faecium 45 株,占分離菌株總數的55.6%;L. plantarum 25 株,占分離菌株總數的30.86%;L. casei 4 株,占分離菌株總數的4.94%;L. pentosus 3 株,占分離菌株總數的3.70%;L. brevis 2 株,占分離菌株總數的2.5%;E. durans 2 株,占分離菌株總數的2.5%。E. faecium和L. plantarum是本次分離菌株中數量最多的菌種,同時大多數樣品中均分離出了這兩種乳酸菌,因此,可以推斷L. plantarum和E. faecium是本次采集的25 份辣白菜樣品中的乳酸菌優勢菌種。
2.2.3 辣白菜中乳酸菌的分離溫度分布分析
從沈陽25 份辣白菜樣品中共分離出81 株乳酸菌,結合表2對這81株乳酸菌的分離溫度進行了對比,結果見表4。

表4 辣白菜中乳酸菌的分離溫度分布結果Table 4 Temperature distribution of lactic acid bacteria in kimchi
由表4可知,20 ℃培養條件下共分離出24 株乳酸菌,占所分離乳酸菌菌株總數的29.63%;37 ℃培養條件下共分離出42 株乳酸菌,占所分離乳酸菌菌株總數的51.85%;45 ℃培養條件下共分離出15 株乳酸菌,占所分離乳酸菌菌株總數的18.52%。不同屬種乳酸菌有著不同的最適生長溫度,本次采集的辣白菜樣品中的乳酸菌大多數最適生長溫度在37 ℃左右。而在20、37、45 ℃培養條件下均分離到了L. plantarum和E. faecium這兩種菌,說明這兩種菌對溫度的適應范圍比較廣,并且對溫度的耐受性較強,屬于辣白菜樣品中的乳酸菌優勢菌種。屬于低溫型菌種的L. brevis只在20 ℃培養條件下分離得到。屬于中低溫型菌種的L. casei和L. pentosus則在20 ℃和37 ℃培養條件下分離到。由此可知,在不同培養溫度下,分離得到的菌群有一定差異,同一類群菌株在不同溫度培養條件下生長情況也不相同,以上結果可為以后乳酸菌的分離鑒定提供參考。
3 結論與討論
國外對韓國泡菜做了較多研究,研究發現其中蘊含著豐富的乳酸菌資源。而國內對于四川泡菜的研究較多,主要集中于對四川泡菜中乳酸菌多樣性分析、特定功能乳酸菌的篩選等,國內外學者均成功從泡菜中分離鑒定出了不同種類的乳酸菌,表明乳酸菌是自然發酵泡菜中的優勢菌群,并在其發酵過程中起著重要作用。同時不同地區和生存環境采集的樣品中,微生物種類、群落結構和生物學特性均有差異。我國東北居住著大量朝鮮族居民,至今仍保留著采用傳統方法制作和食用辣白菜等自然發酵泡菜食品的習慣,再加上我國東北特有的地理、氣候和環境特點,我國東北辣白菜現已成為我國一種獨特的傳統發酵泡菜制品。通過采集沈陽25 份辣白菜樣品,并從中分離鑒定出81 株乳酸菌。其中不但分離到了L. plantarum、L. brevis、L. pentosus、L. casei以及E. faecium等國內外泡菜研究中發現的常見菌種,同時也分離到了2株E. durans,這是其他文獻中鮮有報道的。
我國東北傳統發酵辣白菜中蘊含著豐富多樣的乳酸菌資源,是我國乳酸菌研究和資源開發的一個寶庫。與國內外類似發酵泡菜食品相比,我國東北辣白菜中的乳酸菌種類、分布和組成又有獨特的地區地域特點,可進一步從中挖掘和開發一些新的或特有的乳酸菌優良菌株。
[1] 何擴, 張秀媛, 李育峰, 等. 泡菜優質乳酸茵的篩選及其產酸性能鑒定[J]. 中國調味品, 2012, 37(11): 48-49.
[2] PARK E J, CHUN J, CHA C J, et al. Bacterial community analysis during fermentation of ten representative kinds of kimchi with barcoded pyrosequencing[J]. Food Microbiology, 2012, 30(1): 197-204.
[3] 李淑媛, 張磊. 長貨架期東北辣白菜的研究[J]. 中國食品添加劑, 2014(7): 94-97.
[4] 劉林. 泡菜的制作[J]. 湖南農業, 2002(13): 19-20.
[5] 崔鳳月, 王愈. 辣白菜工藝特色及改進措施的初步探討[J]. 中國釀造, 2011, 30(10): 161-163.
[6] 許雅楠, 池承燈, 姚閩娜. 四川泡菜的制作工藝及風味形成原理[J].農產品加工: 學刊, 2014(7): 31-32.
[7] 南相云, 李璐, 路新國. 韓國泡菜的制作工藝及營養價值[J]. 揚州大學烹飪學報, 2010, 27(2): 46-48.
[8] 李青青, 陳啟和, 何國慶, 等. 我國傳統食品中乳酸菌資源的開發[J].食品科學, 2009, 30(23): 516-520.
[9] 李冬梅, 白月娥, 李玲, 等. SDS-PAGE電泳法分析朝鮮族辣白菜中乳酸菌分布[J]. 食品科技, 2012, 37(11): 10-13.
[10] KIM J, BANG J, LARRY R, et al. Controlled fermentation of kimchi using naturally occurring antimicrobial agents[J]. Food Microbiology, 2012, 32(1): 20-31.
[11] 凌代文, 東秀珠. 乳酸細菌分類鑒定及試驗方法[M]. 北京: 中國輕工業出版社, 1999: 1-25.
[12] 楊瑞, 張偉, 陳煉紅,等. 發酵條件對泡菜發酵過程中微生物菌系的影響[J]. 食品與發酵工業, 2005, 31(3): 90-92.
[13] 楊潔彬, 郭興華. 乳酸菌: 生物學基礎及應用[M]. 北京: 中國輕工業出版社, 1991: 1-3.
[14] 尚天翠. 溫度及pH條件對乳酸菌生長影響的研究[J]. 伊犁師范學院學報: 自然科學版, 2011(3): 33-35.
[15] HOLT J G, SNEATH P H. Bergey’s manual of systematic bacteriology (vol. 2)[M]. 4th ed. Baltimore: Williams & Wilkins, 1986: 1209-1245.
[16] 張豪, 章超樺, 曹文紅, 等. 傳統魚露發酵液中優勢乳酸菌的分離、純化與初步鑒定[J]. 食品工業科技, 2013, 34(24): 186-188.
[17] 高娃. 四川部分地區泡菜中乳酸菌的分離鑒定[D]. 呼和浩特: 內蒙古農業大學, 2010: 10-11.
[18] 奧斯伯F, 布倫特R, 金斯頓R E, 等. 精編分子生物學實驗指南[M].顏子穎, 王海林, 譯. 北京: 科學出版社, 1998: 37-39.
[19] 馮廣達, 陳美標, 羊宋貞, 等. 用于PCR擴增的細菌DNA提取方法比較[J]. 華南農業大學學報, 2013, 34(3): 440-442.
[20] 烏日娜. 內蒙古傳統酸馬奶中乳桿菌的分離鑒定及16S rDNA序列同源性分析[D]. 呼和浩特: 內蒙古農業大學, 2005: 14-15.
[21] SCARPELLINI M, MORA D, COLOMBO S, et al. Development of genus/species-specific PCR analysis for identification of Carnobacterium strains[J]. Current Microbiology, 2002, 45(1): 24-29.
[22] 張潔, 徐桂花, 尤麗琴. 16S rDNA序列分析法鑒定乳酸菌[J]. 農產品加工: 創新版, 2009(4): 47-49.
[23] WU R, WANG L P, MENGHE B, et al. Isolation and preliminary probiotic selection of Lactobacilli from koumiss in Inner Mongolia[J]. Journal of Basic Microbiology, 2009, 49(3): 318-326.
[24] 武俊瑞, 李欣, 李曉忱, 等. 自然發酵酸菜汁中乳桿菌的分離鑒定[J].食品科學, 2012, 33(15): 191-194.
[25] 李鐵. GenBank數據庫檢索及其應用: Entrez檢索功能[J]. 中華醫學圖書情報雜志, 2008, 17(5): 49-51.
Isolation and Identification of Lactic Acid Bacteria from Kimchi
MENG Lingshuai1, ZHANG Ying1, ZOU Tingting1, WU Rina1,2, YUE Xiqing1, WU Junrui1,*
(1. College of Food Science, Shenyang Agricultural University, Shenyang 110866, China; 2. State Key Laboratory of Food Science and Technology, School of Food Science and Technology, Jiangnan University, Wuxi 214122, China)
Lactic acid bacteria were isolated and identified from 25 Korean traditional fermented kimchi samples from 7 districts of Shenyang. Totally 81 strains suspected of being lactic acid bacteria were isolated successfully, and 34 of the isolated strains were identifi ed as bacilli and 47 were identifi ed as cocci. These 81 strains were further identifi ed by 16S rDNA sequence analysis. The results showed that all these 81 strains were confi rmed as lactic acid bacteria from 2 genus and 6 species, including 45 strains of Enterococcus faecium, 25 strains of Lactobacillus plantarum, 4 strains of Lactobacillus casei, 3 strains of Lactobacillus pentosus, 2 strains of Lactobacillus brevis, and 2 strains of Enterococcus durans. These results can provide a basis for further studies on lactic acid bacteria from kimchi in northeast China.
kimchi; Lactobacillus; 16S rDNA; homology analysis
TS201.3
A
10.7506/spkx1002-6630-201511025
2014-11-25
國家自然科學基金面上項目(31470538;31370502);中國博士后科學基金資助項目(2014M560395);
遼寧省農業領域青年科技創新人才培養資助計劃項目(2014048);江蘇省博士后科研資助計劃項目(1402071C);沈陽農業大學“天柱山英才支持計劃”項目
孟令帥(1989—),男,碩士研究生,研究方向為食品生物技術。E-mail:15040260380@163.com
*通信作者:武俊瑞(1977—),男,副教授,博士,研究方向為食品生物技術。E-mail:junruiwu@126.com
