非天然磷酰化氨基酸對胰蛋白酶的熒光猝滅效應(yīng)
2014-12-25 02:27:30符芳妮鄧少平陳麗春
食品與生物技術(shù)學(xué)報(bào) 2014年2期
符芳妮 , 張 暉 , 鄧少平 , 陳麗春 *,2
(1.浙江工商大學(xué) 食品與生物工程學(xué)院,浙江 杭州 310012;2.浙江科技學(xué)院 生物與化學(xué)工程學(xué)院,浙江 杭州310023)
胰蛋白酶(Trypsin)是一種廣泛分布于生物體內(nèi)的特異性最強(qiáng)的蛋白水解酶,在食品加工、細(xì)胞培養(yǎng)、臨床醫(yī)學(xué)等眾多領(lǐng)域具有重要-的應(yīng)用[1-2]。在食品加工技術(shù)中,酶法技術(shù)能夠用于食品蛋白質(zhì)的改性[3-4],例如,胰蛋白酶已經(jīng)廣泛用于改善食品物理化學(xué)性質(zhì)、提高食品功能性質(zhì)、延長貨架期、增加營養(yǎng)特性。國內(nèi)外關(guān)于有機(jī)小分子物質(zhì)與蛋白酶的相互作用已有很多研究[5-9]。目前,清華大學(xué)趙玉芬研究小組關(guān)于天然磷酰化氨基酸與生物大分子物質(zhì)的相互作用已有[10],但是,非天然氨基酸磷酰化前后對蛋白質(zhì)(胰蛋白酶)結(jié)構(gòu)影響的應(yīng)用基礎(chǔ)研究還未有報(bào)道,發(fā)現(xiàn)并研究新的有機(jī)物質(zhì)滿足各個領(lǐng)域的需求已成為需要解決的一個問題。
N-磷酰化氨基酸(N-phosphoryl amino acid)是在天然-α-氨基酸的氨基上引入磷酰基團(tuán)[11]。作者在磷酰化氨基酸具有生物活性的基礎(chǔ)上[11],利用熒光光譜法進(jìn)一步研究非天然脂肪酸鏈磷酰化氨基酸與胰蛋白酶的相互作用,以期深入探究非天然脂肪酸鏈磷酰化氨基酸在抑制胰蛋白酶活性方面的作用性質(zhì)。為了研究非天然氨基酸的磷酰化對胰蛋白酶結(jié)構(gòu)的影響,作者自主設(shè)計(jì)并合成了4種側(cè)鏈長度不同的非天然脂肪酸鏈磷酰化氨基酸,用熒光法比較了非天然脂肪酸鏈氨基酸磷酰化前后對胰蛋白酶結(jié)構(gòu)的影響,對濃度、側(cè)鏈長度不同的磷酰化氨基酸對胰蛋白酶熒光猝滅的影響等進(jìn)行研究。為探索和開發(fā)酶活性調(diào)節(jié)物提供了基礎(chǔ),也為蛋白質(zhì)結(jié)構(gòu)與功能的研究提供了思路,有利于進(jìn)一步研究磷酰化氨基酸對胰蛋白酶活性的調(diào)節(jié)作用及可能的機(jī)制。
1 材料與方法
1.1 材料與儀器
試劑:L-2-氨基戊酸、L-2-氨基庚酸、L-2-氨基辛酸、L-2-氨基壬酸:質(zhì)量分?jǐn)?shù)均為98.5%,購于上海濱海瀚鴻生化有限公司;胰蛋白酶:購于吉諾生物醫(yī)藥技術(shù)有限公司;Tris-HCl緩沖溶:杭州匯普化工有限公司;核磁試劑(CDCl3):購于Sigma公司;三乙胺、乙醇、四氯化碳、亞磷酸二異丙酯:均為市售分析純試劑;超純水。
儀器:500AVANCE III型核磁共振波譜儀,德國 Bruker公司;Therm LCQ TM Deca XP plus離子阱質(zhì)譜儀:美國Thermo公司;三維熒光光譜儀(F-7000):日本日立公司;RE-52c旋轉(zhuǎn)蒸發(fā)儀:鞏義市英峪予華儀器廠;DF-101 Z集熱型磁力攪拌器:鄭州長城科工貿(mào)有限公司;制冰機(jī):日本SANYO SIMF140公司;電熱恒溫水槽:上海精宏實(shí)驗(yàn)設(shè)備有限公司;B0500891型去離子超純水儀:美國Millipore公司。
1.2 實(shí)驗(yàn)方法
1.2.1 非天然氨基酸的磷酰化與結(jié)構(gòu)表征 合成方法采用趙玉芬院士研究小組的亞磷酸二異丙酯四氯化碳水相合成法[12-14]。以L-2-氨基戊酸、L-2-氨基庚酸、L-2-氨基辛酸、L-2-氨基壬酸為底物,亞磷酸二異丙酯(DIPPH)為磷酰化試劑。合成路線如下:

4種分子的結(jié)構(gòu)式分別為

產(chǎn)物結(jié)構(gòu)經(jīng)1H NMR鑒定,結(jié)果見表1。

表1 N-二異丙氧磷酰化氨基酸的1H NMR化學(xué)位移Table 1 1H NMR chemical shift of NIPP-AA
經(jīng)ESI--MS鑒定,結(jié)果見圖1。


圖1 ESI--MS分析譜圖Fig.1 Spectra of ESI--MS
4種磷酰化氨基酸的理論相對分子質(zhì)量分別為281.29、309.34、323.37、337.39。 質(zhì)譜圖中顯示的質(zhì)荷比 280.1、308.2、322.2、336.2 分別為產(chǎn)物的分子離子峰,檢測電離源為ESI-,說明產(chǎn)品中有DIPPL-2-NH2-戊酸、庚酸、辛酸、壬酸。同時,圖1雜峰很少,主峰突出,能夠確定合成產(chǎn)物為目標(biāo)產(chǎn)物。
1.2.2 溶液的配制 室溫下取Tris-HCl緩沖溶液,配制0.25%胰蛋白酶溶液。500 V掃描電壓,波長掃描范圍為300~400 nm,發(fā)射與激發(fā)狹縫為5、5 nm,于5 mL樣品池中加入樣品,測其熒光光譜。
室溫下取Tris-HCl緩沖溶液配制相同濃度的胰蛋白酶溶液6個樣品,恒溫水浴槽溫度分別設(shè)為25、30、35、40、45、50 ℃,將 6 個樣品分別放入水浴槽中達(dá)到相應(yīng)的溫度。保持溫度恒定,掃描電壓為500 V,固定激發(fā)波長λex為280 nm,發(fā)射與激發(fā)狹縫為 5、5 nm。
室溫下分別配制3 mmol/L的L-2-氨基戊酸、庚酸、辛酸及壬酸溶液。向3 mmol/L的L-2-氨基戊酸、庚酸、辛酸及壬酸溶液中依次加入相同體積的已配制的胰蛋白酶溶液。以胰蛋白酶溶液為空白作對照測熒光強(qiáng)度。熒光光譜的掃描參數(shù)同上。
室溫下取超純水分別配制3 mmol/L的N-磷酰化L-2-氨基戊酸、庚酸、辛酸及壬酸溶液。向3 mmol/L的N-磷酰化L-2-氨基戊酸、庚酸、辛酸及辛酸溶液中依次加入相同體積的胰蛋白酶溶液。以胰蛋白酶溶液為空白作對照測熒光光譜。熒光光譜的掃描參數(shù)同上。
25 ℃分別配制 1、3、6、8、10、12、15 mmol/L 的磷酰化庚酸溶液,分別加入相同體積的胰蛋白酶溶液,在恒溫水浴槽中保持恒溫。以胰蛋白酶溶液為空白對照測熒光光譜。熒光光譜的掃描參數(shù)同上。
2 結(jié)果與分析
2.1 胰蛋白酶的熒光特性
胰蛋白酶具有熒光特性,圖2為0.25%胰蛋白酶溶液的三維熒光光譜。可以看出,當(dāng)激發(fā)波長λex=280 nm時,在發(fā)射波長λem=335 nm附近有較強(qiáng)的熒光信號。

圖2 25℃時0.25%胰蛋白酶溶液的三維熒光光譜圖Fig.2 Tridimensional fluorescence spectrum of 0.25%trypsin
2.2 溫度對胰蛋白酶熒光發(fā)射光譜的影響
眾所周知,蛋白質(zhì)遇熱會變性,通常蛋白質(zhì)的最大激發(fā)波長在280 nm左右。
表2為胰蛋白酶溶液在溫度分別為25、30、35、40、45、50℃時的熒光強(qiáng)度及發(fā)射波長。胰蛋白酶在25~40℃時,其最大發(fā)射波長并沒有發(fā)生明顯的位移,說明溫度從25℃升高到40℃時,僅僅是胰蛋白酶的熒光強(qiáng)度發(fā)生變化,并沒有引起其構(gòu)型的改變。但是當(dāng)溫度超過40℃時,其發(fā)射波長有稍微的偏移,熒光強(qiáng)度也發(fā)生了變化,所以可以預(yù)測溫度大于40℃時,胰蛋白酶的構(gòu)型會發(fā)生部分的改變。選擇溫度25℃,避免溫度對胰蛋白酶結(jié)構(gòu)的影響。

表2 溫度對胰蛋白酶熒光發(fā)射光譜的影響Table2 Effectoftemperatureon thefluorescence spectrum of trypsin
2.3 非天然脂肪酸鏈氨基酸磷酰化前后與胰蛋白酶的相互作用
圖3顯示了4種相同濃度的非天然脂肪酸鏈氨基酸(即L-2-氨基戊酸、庚酸、辛酸、壬酸)存在時胰蛋白酶溶液的熒光強(qiáng)度值。4種非天然氨基酸本身沒有熒光特性,不會對實(shí)驗(yàn)結(jié)果產(chǎn)生干擾。從圖3可以看到,L-2-氨基戊酸、庚酸、辛酸存在時,胰蛋白酶溶液的熒光強(qiáng)度基本沒有發(fā)生變化,說明這3種非天然氨基酸并不能使胰蛋白酶的熒光強(qiáng)度減弱。L-2-氨基壬酸存在時,胰蛋白酶的最大熒光強(qiáng)度從1 680降低到1 570,但總體而言胰蛋白酶的熒光強(qiáng)度并沒有明顯減弱。并且胰蛋白酶的發(fā)射波長沒有發(fā)生偏移,因此,L-2-氨基戊酸、庚酸、辛酸及壬酸4種非天然氨基酸對胰蛋白酶的熒光基本不存在猝滅效應(yīng),并且隨著氨基酸側(cè)鏈碳原子數(shù)的增加,不會對胰蛋白酶的熒光強(qiáng)度產(chǎn)生影響。

圖3 25℃時4種氨基酸存在時0.25%胰酶的熒光發(fā)射光譜Fig.3 Fluorescence intensity of 0.25%trypsin solution(c=3 mmol/L)in the presence of four non-natural amino acids
為了研究4種非天然氨基酸磷酰化后對胰蛋白酶的作用,考察了相同濃度的4種非天然氨基酸及其衍生的非天然脂肪酸鏈磷酰化氨基酸分別對胰蛋白酶熒光強(qiáng)度的影響,見圖4。4種磷酰化氨基酸本身沒有熒光性,對實(shí)驗(yàn)不存在干擾。圖3與圖4相比,3 mmol/L的L-2-氨基戊酸、庚酸、辛酸、壬酸對胰蛋白酶熒光沒有猝滅效應(yīng),但3 mmol/L的磷酰化氨基酸能夠使胰蛋白酶溶液的熒光強(qiáng)度明顯降低。由此表明,磷酰基的引入顯著改變了非天然氨基酸的分子的性質(zhì)。

圖4 25℃0.25%胰蛋白酶溶液在4種非天然氨基酸分子及4種N-磷酰化氨基酸存在時的熒光強(qiáng)度值Fig.4 Fluorescence intensity of 0.25%trypsin solution in the presence of different molecules at 25℃
圖5為0.25%胰蛋白酶的熒光強(qiáng)度隨4種相同濃度、不同側(cè)鏈長度的N-磷酰化氨基酸的變化圖。從圖5可以看出,4種磷酰化氨基酸側(cè)鏈長度不同,對熒光強(qiáng)度的猝滅效應(yīng)也不同。隨著磷酰氨基酸側(cè)鏈碳原子數(shù)目的增加,胰蛋白酶溶液的熒光強(qiáng)度逐漸降低。空白的胰蛋白酶的熒光強(qiáng)度為2 549,熒光猝滅后的熒光強(qiáng)度則分別變?yōu)? 149、2 078、1 897、1 682。從而說明,磷酰化氨基酸的側(cè)鏈長度能夠?qū)σ鹊鞍酌傅臒晒鈴?qiáng)度產(chǎn)生影響。
2.4 不同濃度的磷酰化庚酸與胰蛋白酶的熒光猝滅作用
圖6(a)中,胰蛋白酶溶液的熒光強(qiáng)度隨著N-磷酰化庚酸的濃度的增大而不斷降低。N-磷酰化庚酸對胰蛋白酶溶液有明顯的熒光猝滅效應(yīng)。由于4種N-磷酰化氨基酸只是側(cè)鏈長度不一樣的氨基酸,其他結(jié)構(gòu)均相同,因此可知,其它N-磷酰化戊酸、辛酸、壬酸對胰蛋白酶的猝滅效應(yīng)隨其濃度的增加具有同樣的趨勢。

圖5 25℃時4種鏈長不同的N-磷酰化氨基酸存在時0.25%胰酶的熒光發(fā)射光譜Fig.5 Fluorescence intensity of 0.25%trypsin solution in the presence of four new N-phosphoryl amino acids at 25℃
從圖6(b)可以看出,胰蛋白酶溶液的熒光最大發(fā)射光譜在加入N-磷酰化庚酸溶液后沒有發(fā)生變化,即其最大發(fā)射光譜沒有隨著猝滅劑濃度的增加而改變,一直在335 nm左右,僅僅是其最大熒光強(qiáng)度逐漸減小。其發(fā)射峰及峰型基本不變,表明其存在相互作用,由此可以說明,25℃時在N-磷酰化庚酸猝滅劑存在時,胰蛋白酶的構(gòu)型并沒有發(fā)生變化,是熒光猝滅劑與胰蛋白酶之間的相互作用猝滅了胰蛋白酶的熒光。


圖6 25℃時不同濃度N-磷酰化庚酸與胰蛋白酶的熒光強(qiáng)度和發(fā)射光譜Fig.6 Fluorescence spectra oftrypsin solution as function of different concentration of DIPP-L-2-Amino heptanoic acid at 25℃
猝滅過程實(shí)際上是與發(fā)光過程相互競爭而縮短發(fā)光分子激發(fā)態(tài)壽命的過程 ,猝滅過程可能發(fā)生于猝滅劑與熒光物質(zhì)的激發(fā)態(tài)之間的相互作用,也可能發(fā)生于猝滅劑與熒光物質(zhì)的基態(tài)分子之間的相互作用。前一過程稱為動態(tài)猝滅,后一種稱為靜態(tài)猝滅[15]。為了解釋這個猝滅過程,按照動態(tài)猝滅的Stern-Volmer方程[16]處理,即:

F0、F分別表示未加入化合物和加入化合物后胰蛋白酶的熒光強(qiáng)度,ksv是動態(tài)猝滅常數(shù),kq表示雙分子猝滅過程中的速率常數(shù),τ0為猝滅劑不存在時生物大分子的平均壽命。
25~40℃時胰蛋白酶的分子結(jié)構(gòu)不變,以F0/F對c[DIPP-L-heptanoicacid]作圖,作出25℃不同濃度的N-磷酰化庚酸對胰蛋白酶熒光猝滅的Stern-Volmer圖,見圖7。
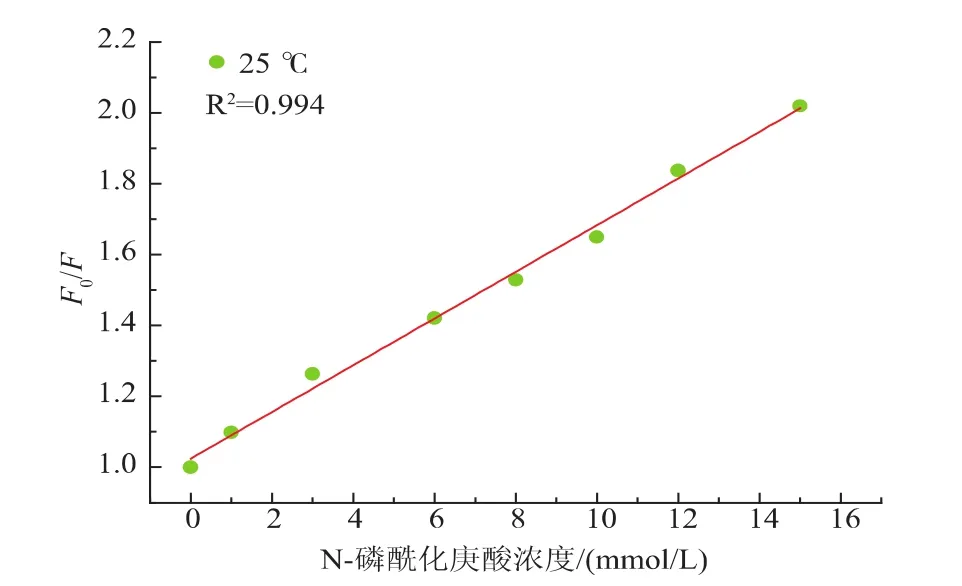
圖7 25℃時N-磷酰化庚酸與胰蛋白酶相互作用的Stern-Volmer曲線Fig.7 Stern-Volmer plot of DIPP-L-2-Amino heptanoic acid quenching effect on trypsin solution at 25℃
從圖7可以看出,25℃時F0/F與磷酰化氨基酸的濃度呈良好的線性關(guān)系,線性相關(guān)系數(shù)R2=0.994。進(jìn)一步說明了胰蛋白酶的熒光完全可以酰化庚酸所猝滅。結(jié)果表明,磷酰化氨基酸對胰蛋白酶熒光的猝滅是靜態(tài)猝滅,磷酰化氨基酸與胰蛋白酶在基態(tài)由于親和力的作用形成配合物降低了酶的熒光強(qiáng)度,從而能夠?qū)σ鹊鞍酌府a(chǎn)生熒光猝滅。
3 結(jié)語
作者設(shè)計(jì)了側(cè)鏈長度不同的非天然脂肪酸鏈磷酰化氨基酸,熒光法研究了非天然磷酰化氨基酸與胰蛋白酶的相互作用,熒光光譜結(jié)果表明,磷酰化氨基酸對胰蛋白酶能夠產(chǎn)生熒光猝滅;非天然L-型氨基酸不能對胰蛋白酶產(chǎn)生熒光猝滅,并且隨著磷酰化氨基酸側(cè)鏈的增長,猝滅效應(yīng)越明顯。同時,隨著磷酰化庚酸的濃度增大,熒光強(qiáng)度不斷降低,Stern-Volmer曲線表明,胰蛋白酶溶液的熒光能夠完全被猝滅。L-型氨基酸的磷酰化能夠改變氨基酸的性質(zhì),使其對胰蛋白酶更具有親和力,Stern-Volmer曲線表明,胰蛋白酶溶液的熒光可以完全被猝滅。胰蛋白酶與非天然脂肪酸鏈磷酰化氨基酸之間相互作用的研究有利于探索和開發(fā)酶活性調(diào)節(jié)物,也為蛋白質(zhì)結(jié)構(gòu)與功能的研究提供了思路,能夠改善胰蛋白酶在食品、生物技術(shù)等領(lǐng)域的應(yīng)用,但磷酰化氨基酸對胰蛋白酶的調(diào)節(jié)作用機(jī)制有待進(jìn)一步研究。
[1]HU Lianzhe,HAN Shuang,Saima Parveen,et al.Highly sensitive fluorescent detection of trypsin based on BSA-stabilize gold nanoclusters[J].Biosensors and Bioelectronics,2012,32:297-299.
[2]程偉,陳紅兵,高金燕,等.酶改性對食物蛋白質(zhì)過敏原性的影響[J].食品科學(xué),2010,31(23):391-394.CHENG Wei,CHEN Hongbin,GAO Jinyan,et al.A review of the effect of enzymatic modification on the allergenicity of food proteins[J].Food Science,2010,31(23):391-94.(in Chinese)
[3]Da Ren,Pipes Gary D,Liu Dingjiang,et al.An improved trypsin digestion method minimize digestion-induced modification on proteins[J].Anayltical Biochemistry,2009,392:12-21.
[4]FANG Meijuan,LUO Shuna,WANG Heqing,et al.The effect of phosphoryl oxygen on the intermolecular action of alanin and lysozyme[J].Acta Phys-Chim Sin,2005,21(9):1042-1045.
[5]XUE Wangxin,ZHANG Guanxin,ZHANG Deqing,et al.A new label-free continuous fluorometric assay for trypsin and inhibitor screening with tetraphenylethene compunds[J].Ogranic Letters,2010,12(10):2274-2277.
[6]強(qiáng)黎明,董雪茹,呂名秀,等.N-磷酰化肽酯及小肽與溶菌酶相互作用的ESI-MS研究[J].化學(xué)學(xué)報(bào),2009,67(22):2607-2612.QIANG Liming,DONG Xueru,LU Mingxiu,et al.Study on the interaction between N-phosphoryl dipeptides (or methyl esters)and lysozyme by ESI-MS[J].Acta Chimica Sinica,2009,67(22):2607-2612.(in Chinese)
[7]楊冉,陳曉嵐,李萍,等.熒光法研究三種黃酮小分子與溶菌酶的相互作用[J].高等學(xué)校化學(xué)學(xué)報(bào),2006,27(9):1673-1676.YANG Ran,CHEN Xiaolan,LI Ping,et al.Study on the interactions between three flavones and lysozyme by fluorescence method[J].Chemical Journal of Chinese Universities,2006,27(9):1673-1676.(in Chinese)
[8]查雋,何華,劉鐵兵,等.熒光光譜法研究共存碳納米管對牛血清白蛋白與加替沙星相互作用的影響[J].光譜學(xué)與光譜分析,2011,31(1):149-153.CHA Jun,HE Hua,LIU Tiebin,et al.Studies on the interaction of gatifloxacin with bovine serum albumin in the presence of carbon nanotubes by fluorescence spectroscopy[J].Spectroscopy and Spectral Analysis,2011,31(1):149-153.(in Chinese)
[9]CUI Xiaodong,WANG Zhuanhua,LI Yuying,et al.Buckwheat trypsin inhibitor enters Hep G2 cells by clathrin-dependent endocytosis[J].Food Chemistry,2013,141:2625-2633.
[10]李永芳.N-磷酰化氨基酸與生物大分子的相互作用[D].北京:清華大學(xué),1995.
[11]趙玉芬,張建臣,徐軍,等.磷酰化氨基酸的特性及應(yīng)用前景[J].鄭州大學(xué)學(xué)報(bào),2003,35(2):89-91.ZHAO Yufen,ZHANG Jianchen,XU Jun,et al.Properties and applications of N-phosphoryl amino acids[J].Journal of Zhenzhou University,2003,5(2):89-91.(in Chinese)
[12]CHEN Zhongzhou,TONG Yufeng,CHEN Shuibing,et al.Orientation of the peptide formation of N-phosphoryl amino acid in solution[J].Chinese Science Bulletin,2002(47):1866-1870.
[13]LIU Huachen,DONG Aijun,TAN Chunyan,et al.The design,synthesis,and anti-tumor mechanism study of N-phosphoryl amino acid modified resveratrol analogues[J].Bioorganic & Medicinal Chemistry,2008,16:10013-10021.
[14]JI Gaijiao,XUE Chubiao,ZENG Jianing,et al.Synthesis of N-(Diiopropyloxyphoryl) amino acids and peptides[J].Synthesis,1988,6(11):444-448.
[15]CHEN Zhongxiu,SU Xiaoxia,DENG Shaoping.Molecular recognition of melamine by vesicles spontaneously formed from orotic acid derived bolaamphiphiles[J].Journa of Physical Chemistry B,2011,115:1798-1806.
[16]陳曉嵐,于斐,屈凌波,等.熒光及ESI質(zhì)譜法研究溶菌酶與磷酰化黃酮的相互作用[J].化學(xué)學(xué)報(bào),2004,62(2):188-193.CHEN Xiaolan,YU Fei,QU Lingbo,et al.Study on the interaction of lysozyme and diethyl flavon-7-yl phosphate by ESI-MS and fluorescence method[J].Acta Chimica Sinica,2004,62(2):188-19.(in Chinese)
