幾種雅羅魚亞科魚類基于mtDNA 序列的親緣關系
2014-12-23 11:30:16竇新杰常玉梅梁利群
江蘇農業學報 2014年4期
竇新杰, 常玉梅, 唐 然, 陶 然,3, 梁利群
(1.淡水魚類育種國家地方聯合工程實驗室,農業部淡水水產生物技術與遺傳育種重點實驗室,中國水產科學研究院黑龍江水產研究所,黑龍江 哈爾濱150070;2.上海海洋大學水產與生命學院,上海201306;3.寧波大學海洋學院,浙江 寧波315211)
雅羅魚亞科(Leuciscinae)隸屬于鯉形目(Cyprinoidei)鯉科(Cyprinidae),在中國南北方均有分布,其中北方主要以雅羅魚屬(Leuciscus cuvier)為主[1-2]。雅羅魚屬約有20 種[1],在中國有7 種,其中瓦氏雅羅魚(L.waleckii)主要分布于黑龍江流域包括呼倫湖、黑龍江、松花江、烏蘇里江,此外在達里湖、岱海灤河、黃河、遼河以及俄羅斯境內的黑龍江上游江段也有少量分布;圖們雅羅魚(L.waleckii tumensisMori)分布于圖們江;準噶爾雅羅魚(Leuciscus merzbacheri)分布于準噶爾盆地;高體雅羅魚(L.idus)和貝加爾雅羅魚(L.leuciscus baicalebsis)主要分布在中國新疆額爾齊斯河水系。灘頭雅羅魚(Tribolodon branditDybowski)和珠星雅羅魚(L.haronensisGunther)也主要分布在中國北方綏芬河和圖們江流域[3-4],并且有海邊(半咸水區)溯河產卵現象。這些種類在分布區是具有重要經濟價值的魚類。
雅羅魚亞科的魚類具有對不良環境超強的適應性。如瓦氏雅羅魚能夠在pH 9.6,堿度高達50 mmol/L的高鹽堿湖泊-達里湖中生存[5-8],而珠星雅羅魚能夠在pH3.5 的酸性湖泊-日本宇曾利山湖中存活[9]。那么,對當地環境的適應是否造成不同種群遺傳結構發生變異,以及這種變異是否導致不同種群進化成不同的物種?池炳杰等[10]利用微衛星標記檢測了瓦氏雅羅魚淡水群體和堿水群體的遺傳組成,發現2 個群體出現了明顯的遺傳分化,進一步深入研究發現,除有限的基因交流造成的地域種群分化外,由環境因子(如堿度)誘發的自然選擇作用也是導致不同群體分化的主要進化力量[11]。此外,灘頭雅羅魚和珠星雅羅魚由于生活環境的不同(海水或酸性水),有些學者依據表型和遺傳數據將這2 個種劃分為3 塊魚屬而非雅羅魚屬[12]。劉金亮等[13]利用微衛星標記估算了4 個雅羅魚種5 個不同群體的遺傳距離,發現貝加爾雅羅魚和高體雅羅魚遺傳距離小,聚為一類,并與灘頭雅羅魚的遺傳距離較近;瓦氏雅羅魚的達里湖群體和松花江群體遺傳距離小,聚為另一類。
動物線粒體DNA(mtDNA)結構相對簡單,在進化過程中與核基因平行進化,屬于母系遺傳,具有進化速率快,不易發生重組等特點,是目前分析群體遺傳結構、種間遺傳分化以及種屬親緣關系鑒定的重要工具[14-15]。本研究選擇mtDNA 的COI、Cytb和Dloop3 個基因,對瓦氏雅羅魚的2 個地理群體(松花江雅羅魚和達里湖雅羅魚)、高體雅羅魚、貝加爾雅羅魚、灘頭雅羅魚和珠星雅羅魚進行遺傳多樣性分析,探究這幾種魚類之間的系統發育關系,為進一步探討雅羅魚亞科系統發育關系以及種屬劃分提供基礎資料。
1 材料與方法
1.1 材料
本研究共收集到雅羅魚亞科隸屬于2 個屬的6個種群樣本(表1)。其中雅羅魚屬包括瓦氏雅羅魚的2 個地理群體(松花江群體,SHJ;達里湖群體,DL)、高體雅羅魚(GT)和貝加爾雅羅魚(BJ);三塊魚屬包括灘頭雅羅魚(TT)和珠星雅羅魚(ZX)。樣本采集地分別是松花江綏濱段、內蒙古達里湖、新疆額爾齊斯河、新疆賽里木湖、黑龍江綏芬河、日本宇曾利山湖(Kaneko 教授惠贈)。所有樣本鰭條于95%酒精中保存備用。
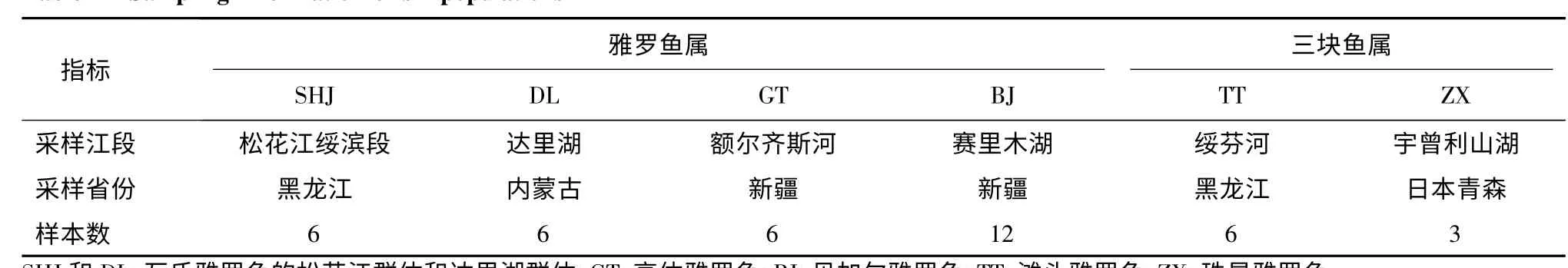
表1 6 個雅羅魚群體的樣本信息Table 1 Sampling information of six populations
1.2 基因組DNA 的提取及PCR 擴增
基因組DNA 的提取方法為常規的酚-氯仿法,具體步驟參照劉金亮等[13]的方法。COI和D-loop引物[16-17]適用于所有樣本,而參照孔慶鵬等的Cytb引物[18]僅適用于雅羅魚屬。因此,根據NCBI 上已公布的灘頭雅羅魚的Cytb序列,利用軟件Primer 5.0 設計1 對引物,引物序列見表2。PCR 反應體系為50 μl,包括35 μl 自制混合Buffer(包括50.0 mmol/L KCl、10.0 mmol/L Tris-HCl、0.10% TritonX-100、1.5 mmol/L MgCl2、0.10% NP-40、0.01%明膠、4 種dNTP),濃度為10.0 mmol/L上、下游引物各0.75 μl,50 ng 的基因組2 μl,TaqDNA 聚合酶1U(Fermentas)、去離子無菌水補至總體積50 μl。PCR反應條件為:94 ℃預變性5 min;35 個循環包括94℃30 s,55 ℃或52 ℃30 s(表2),72 ℃1 min;最后72 ℃延伸10 min。PCR 擴增產物用1.5%的瓊脂糖凝膠電泳檢測后,送至上海立菲生物技術有限公司完成測序。

表2 COI、Cytb 和D-loop 的PCR 引物信息Table 2 Information of PCR primers for three regions of mtDNA
1.3 序列處理及分析
利用Clustalx1.8 和Bioedit7.0 分別對COI、Cytb和D-loopmtDNA 基因序列進行多重比對、校正。利用MEGA5.1 計算序列4 種核苷酸組成比例、堿基轉換∕堿基顛換值(R),基于Kimura 雙參數模型計算種間遺傳距離,用鄰接法(Neighbor-joining method,NJ 法)構建系統發育進化樹,Bootstrap (重復1 000 次)檢驗系統樹各分支的置信度。利用DNAsp5.0 計算多態位點數(S)、單倍型數(H)、單倍型多樣性(Hd)、核苷酸多樣(Pi)以及平均核苷酸差異數(K)。利用Arlequin3.1 進行種內及種間的分子方差分析(AMOVA)。
2 結果
2.1 COI 基因多樣性
序列經比對校正后,得到COI部分的有效序列長度為635 bp。COI基因在6 個種群中共分析了39 條有效序列。核苷酸組成分析結果顯示,腺嘌呤(A )、胸腺嘧啶(T)、鳥嘌呤(G)、胞嘧啶(C)4 種堿基的平 均 含 量 依 次 為24.53%、28.13%、19.05% 和18.19%,A + T 含量(52.70%)略高于G + C 含量(47.3%),堿基轉換與堿基顛換比(R)為3.394。單倍型分析顯示,所有樣本共有16 個單倍型,其中DL和SHJ 共享單倍型且數量為2 個;GT 和ZX 各獨享1個單倍型(表3)。遺傳多樣性參數統計顯示,貝加爾雅羅魚(BJ)和灘頭雅羅魚(TT)的遺傳多樣性最高,其次為瓦氏雅羅魚的DL 和SHJ 群體,高體雅羅魚(GT)和珠星雅羅魚(ZX)未檢測到遺傳變異(表3)。
2.2 Cytb 和D-loop 基因多樣性
由于Cytb和D-loop分析的樣本相同,因此將這2個基因序列整合為1 個長度為1 095 bp 的片斷(Cytb-D-loop),共分析了39 條有效序列。核苷酸組成分析結果顯示,A、T、G 和C 4 種堿基的平均含量分別為27.93%、31.21%、15.90% 和24.96%,A + T 含 量(59.14%)明顯高于G+C 含量(40.86%),堿基轉換與堿基顛換比(R)為3.27。所有樣本共檢測到34 個單倍型,其中貝加爾雅羅魚(BJ)享有單倍型數最多為11 個;珠星雅羅魚(ZX)享有的單倍型數最少為3個,其余種群的單倍型數為4 ~6 個(表4)。遺傳多樣性分析結果顯示,6 個種群的各項遺傳多樣性參數普遍較高,其中,貝加爾湖雅羅魚(BJ)和高體雅羅魚(GT)各項參數值比較接近;瓦氏雅羅魚的DL 和SHJ群體的遺傳多樣性相近,而灘頭雅羅魚(TT)與珠星雅羅魚(ZX)數值差異大(表4)。
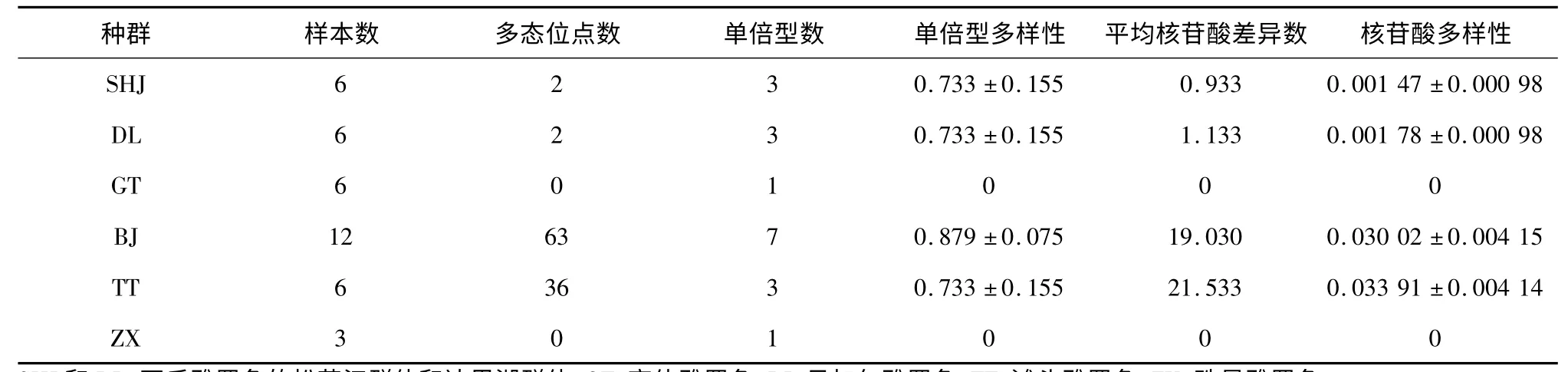
表3 6 個雅羅魚種群COI 基因的遺傳多樣性Table 3 Genetic diversity of six populations based on COI gene
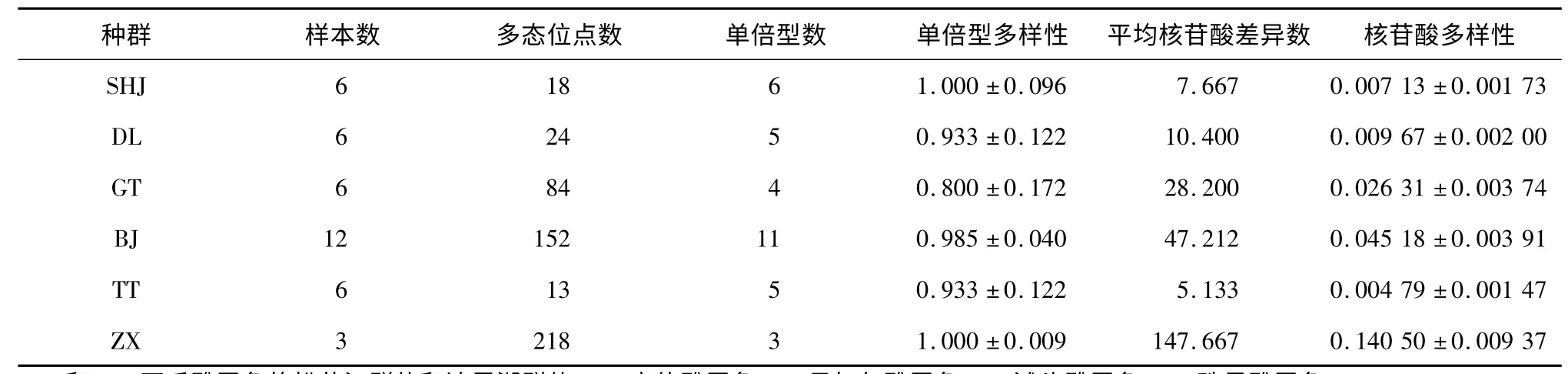
表4 6 個雅羅魚種群Cytb-D-loop 基因的遺傳多樣性Table 4 Genetic diversity of Cytb-D-loop gene of six populations
2.3 群體遺傳結構分析
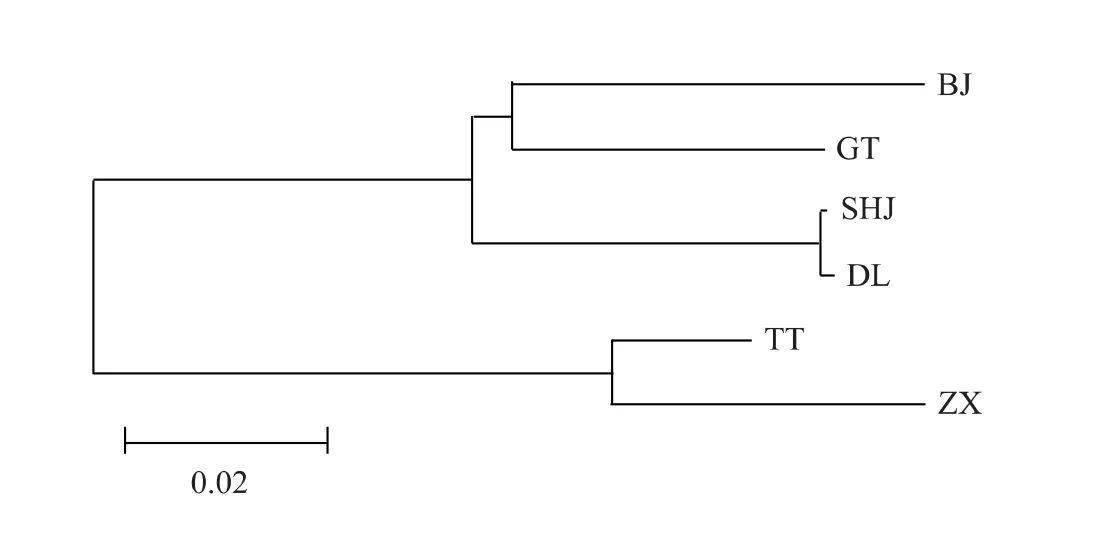
圖1 基于COI 基因的種間遺傳距離構建的系統發育樹Fig.1 Phylogenetic tree constructed with COI gene mtDNA
以Kimura 為雙參數模型計算6 個雅羅魚種群間的遺傳距離。基于COI基因的遺傳距離顯示,6個種群聚為2 大分支,其中SHJ、DL、GT 和BJ 親緣關系較近,聚為一支,而TT 和ZX 聚為另一支(圖1),該聚類支持6 個種群分屬2 個屬,即雅羅魚屬和三塊魚屬。在雅羅魚屬的4 個種群中,瓦氏雅羅魚的2 個群體的遺傳距離最近為0.002,GT 與BJ 之間的距離為0.072,分別與SHJ 和DL 的遺傳距離介于0.068 ~0.082;三塊魚屬中的TT 和ZX 2 個物種的遺傳距離為0.045,與另外4 個種群間的距離介于0.137 ~0.161(表5)。基于Cytb-D-loop基因的親緣關系聚類結果顯示,ZX 單獨聚為一支,且與另外5 個種群間的遺傳距離較遠,介于0.170 ~0.283(表5);除此之外,其他種群的聚類結果與COI相似(圖2)。
種群遺傳分化指數分析顯示,瓦氏雅羅魚種內的2 個地理群體在COI和Cytb-D-loop2 個mtDNA區段均沒有出現明顯的遺傳分化(P>0.05),而不同種間相比,6 個種群在COI和Cytb-D-loop區段出現了不同程度的遺傳分化(P<0.05)(表5)。對6個種群的遺傳結構按照種屬關系進行2 個層次的分子生物學方差分析。其中,按照雅羅魚屬(SHJ、DL、GT、BJ)和三塊魚屬(TT、ZX)2 個屬劃分,組間遺傳結構有明顯分歧(P<0.05);按照瓦氏雅羅魚(SHJ、DL)、高體雅羅魚、貝加爾雅羅魚、灘頭雅羅魚和珠星雅羅魚5 個種劃分,組間差異不顯著(P>0.05)。
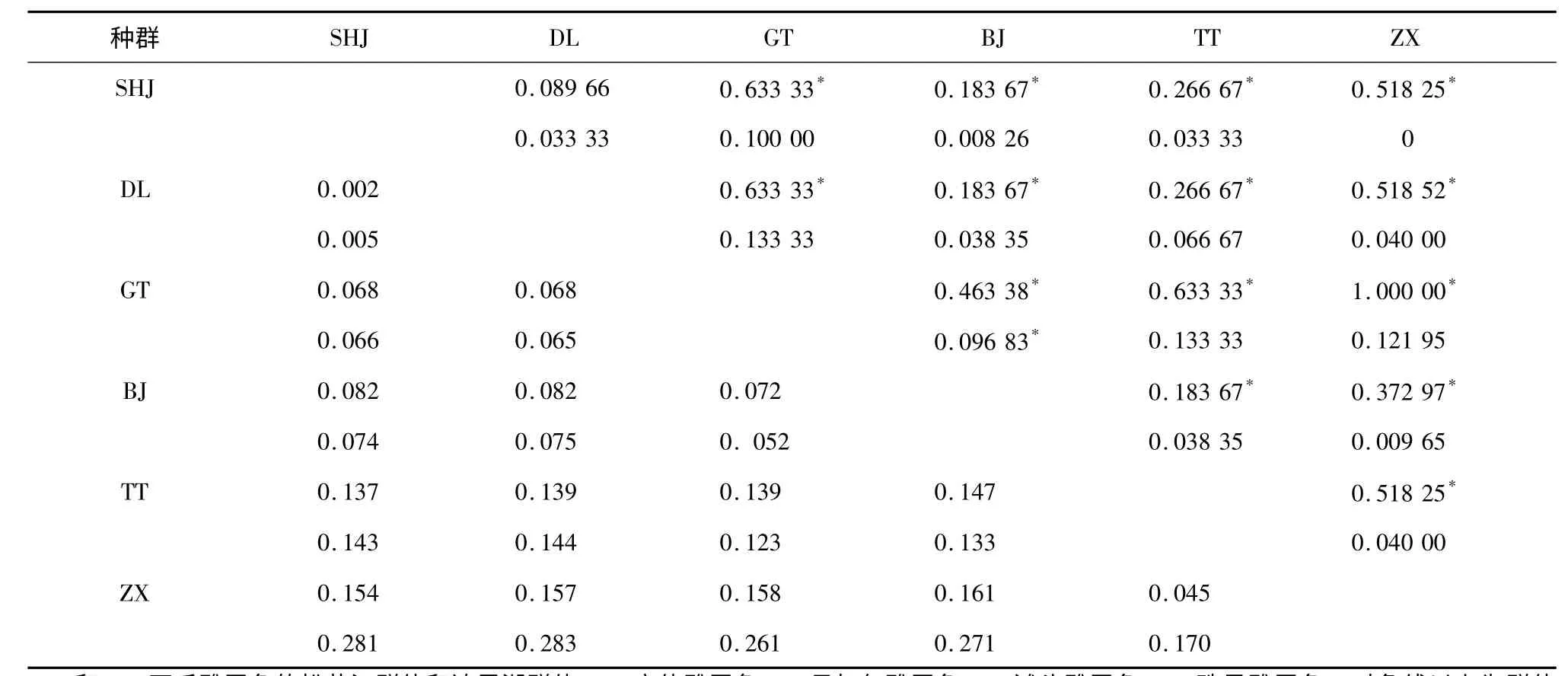
表5 基于COI 和Cytb-D-loop 基因的6 個雅羅魚種群間的遺傳分化指數和遺傳距離Table 5 The genetic differentiation index and genetic distance among six poputations based on COI and Cytb-D-loop genes
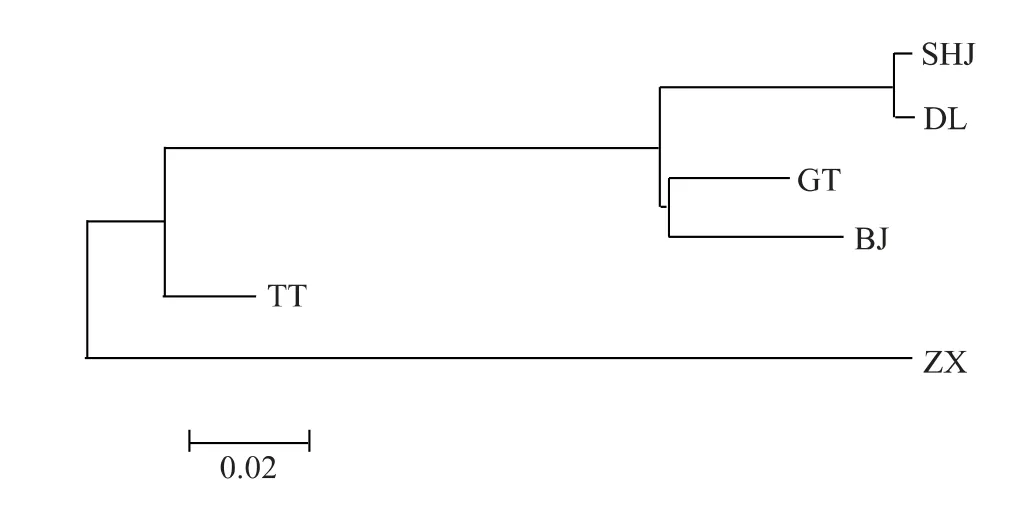
圖2 基于Cytb-D-loop 基因的種間遺傳距離構建的系統發育樹Fig.2 Phylogenetic tree constructed with Cytb-D-loop gene mtDNA
2.4 種群特異位點的鑒定
對6 個種群所特有的多態位點數進行統計,沒有找到能夠區分瓦氏雅羅魚2 個地理群體的特異位點,因此按照5 個種群進行統計,在COI和Cytb-Dloop2 個區段均發現了多個種群特異位點,其中,珠星雅羅魚擁有的特異位點數最多(53 個),依次為瓦氏雅羅魚(47 個)、貝加爾雅羅魚(31 個)、灘頭雅羅魚(27 個)和高體雅羅魚(22 個)(表6)。

表6 基于COI 和Cytb-D-loop 基因的6 個雅羅魚種群的特異位點數Table 6 The specific loci on COI 和Cytb-D-loop genes of mtDNA differentiating six populations
3 討論
3.1 群體的遺傳多樣性
基于6 個種群的COI和Cytb-D-loop基因核苷酸組成分析,結果顯示4 種堿基含量不均一,而且A+T 的比例均高于G+C 的比例,與脊椎動物mtDNA 的特點一致[20]。mtDNA 2 個基因區段的堿基轉換與堿基顛換比(R)依次為3.94 和3.27,這一結果符合脊椎動物線粒體基因組進化時堿基轉換高于堿基顛換的規律;有研究認為當R值小于2 時基因的序列突變達到飽和,在構建系統樹的過程中受進化噪音的影響易出現錯誤信息[21],本研究中的R值均大于2,可進行構建系統樹。從遺傳多樣性上來看,6 個種群在Cytb-D-loop區段的多樣性水平均較高,而在COI區域變異較大,有的種群甚至沒有出現遺傳變異。有報道認為,由于不編碼蛋白質,D-loop區域受進化選擇的影響小,進化速度快,在不同物種或同一物種的不同群體表現出較高的遺傳多樣性[22],這與我們的研究結果是一致的。
3.2 群體遺傳分化及親緣關系
從分組的分子生物學方差分析結果可以看出,將6 個種群分為同一個組,結果顯示組內差異明顯,說明6 個種群可能具有不同的種屬來源;按照雅羅魚屬(SHJ、DL、GT 和BJ)和三塊魚屬(TT、ZX)分組,結果顯示組間差異顯著,說明這6 個種群可能來源于不同的屬;按照瓦氏雅羅魚(SHJ、DL)、高體雅羅魚(GT)、貝加爾雅羅魚(BJ)、灘頭雅羅魚(TT)和珠星雅羅魚(ZX)5 個種劃分,結果顯示組間差異不顯著,說明這5 個種親緣關系較近。
具體來看,DL 和SHJ 2 個地理群體的遺傳距離最近,在COI和Cytb-D-loop區段上的遺傳距離依次為0.002 和0.005,在系統進化樹上始終聚在一起,此外,在本研究分析的2 個mtDNA 區段內沒有發現能夠區分這2 個地理群體的特異位點,但是作為同一個種,卻發現很多共有且有別于其他4 個種群的種內特異位點,說明DL 和SHJ 2 個群體并沒有因為棲息環境的巨大差異而出現明顯的群體分化,應歸屬為瓦氏雅羅魚種的2 個地理群體。池炳杰等[10]和Chang 等[11]利用SSR 標記檢測發現,達里湖水系和黑龍江水系的瓦氏雅羅魚種群由于有限的基因交流和對當地環境的適應性,2 個水系的群體出現了明顯的遺傳分化。但是否將這2 個水系的群體劃分為2 個亞種,論據尚不充分。
貝加爾雅羅魚和高體雅羅魚在COI和Cytb-Dloop基因區段上遺傳距離分別為0.072 和0.052,種群遺傳分化檢驗發現,BJ 和GT 種群在COI和Cytb-D-loop出現了明顯的遺傳分化,而且在2 個基因區段分別發現了多個種內特異位點,應劃分為2 個不同的種,這與胡文閣等[23]的研究結果是一致的,但是系統進化樹支持將這兩個種群聚為一支,說明這2 個種群親緣關系較近。本研究中基于mtDNA 的聚類分析顯示,瓦氏雅羅魚(SHJ、DL)、高體雅羅魚(GT)、貝加爾雅羅魚(BJ)聚為一個大的分支,陳星玉[24]的研究發現,貝加爾雅羅魚在骨骼結構上與瓦氏雅羅魚接近。因此推斷瓦氏雅羅魚、高體雅羅魚和貝加爾雅羅魚具有屬內水平的種間親緣關系。
COI相對于Cytb和D-loop最為保守,進化速度最慢,更適合遠緣物種進化研究[25]。本究中灘頭雅羅魚和珠星雅羅魚在COI區域與另外4 個種群的遺傳距離為0.137 ~0.147 和0.154 ~0.161。張鳳英等[26]在比較2 種鯛屬魚類線粒體COI基因時發現,屬間遺傳距離介于0.138 ~0.182;Costa 等[27]在對澳大利亞122 個魚屬的COI條形碼研究中發現,屬間最大的遺傳距離不超過0.206。根據遺傳距離和系統聚類分析結果以及分子生物學方差分析檢驗結果來看,TT 和ZX 種群與其他種群為屬間親緣關系,這與生物學分類的結果是一致的,即灘頭雅羅魚和珠星雅羅魚應劃分為雅羅魚亞科的三塊魚屬。
根據黃小彧等[28]基于COI界定的種間劃分依據(0.05 ~0.30)來看,TT 和ZX 種群在COI的遺傳距離僅為0.045,不支持種間關系。但是二者在COI區域存在明顯的遺傳分歧,且在2 個mtDNA 區段均能找到多個種內特異位點,而同為種內關系的DL和SHJ 群體在2 個基因區段遺傳分歧不明顯,也沒有群體特異位點,說明灘頭雅羅魚和珠星雅羅魚的遺傳結構存在顯著差異;此外,陳金平[12]在探究綏芬河三塊魚屬的分類中發現,灘頭雅羅魚和珠星雅羅魚在磷片組成和骨骼結構上也存在明顯差異;此外,本研究采集的這2 個種群地理隔離比較遠,棲息環境差異大(海水、酸湖)。因此,結合前人和我們的研究結果,將灘頭雅羅魚和珠星雅羅魚劃分為三塊魚屬的2 個不同的種比較合理。
總之,基于遺傳距離、系統進化樹及種內特異位點的研究結果,我們認為SHJ 和DL 2 個地理群體親緣關系近,同屬于瓦氏雅羅魚;高體雅羅魚和貝加爾雅羅魚親緣關系近,且與瓦氏雅羅魚存在屬內親緣關系,同屬于雅羅魚亞科的雅羅魚屬;灘頭雅羅魚和珠星雅羅魚親緣關系近,且與雅羅魚屬的4 個種群存在屬間親緣關系,同屬于雅羅魚亞科的三塊魚屬。
[1] 陳宜瑜.中國動物志,硬骨魚綱,鯉形目(中)[M].北京:科學出版社,1998.
[2] HU W G,DUAN Z Y,WANG J F,et al.Divergence and systematical evolution of three leuciscus species in xinjiang based on mitochondrial dna control region sequences[J].遺傳學報,2004,31(9):970-975.
[3] 戴定遠,張玉玲,黃浩明.圖們江灘頭魚的生物學研究[J].動物學報,1982,28(3):283-292.
[4] 馬 波,陳金平,董崇智.綏芬河三塊魚和珠星三塊魚種群的生化遺傳變異及親緣關系[J].中國水產科學,2005,12(6):688-693.
[5] 李志明,劉海濤,馮偉業,等.達里湖和崗根湖東北雅羅魚和鯽四種同工酶的比較研究[J].淡水漁業.2008,38(5):26-29.
[6] 孟和平,彭本初,王保文,等.達里湖東北雅羅魚受精卵在不同水體中的孵化試驗[J].內蒙古農業科技,2007 (4):74-75.
[7] 齊景偉,安曉萍,孟和平,等.達里湖瓦氏雅羅魚資源現狀及合理利用探討[J].水生態學雜志,2011(1):71-77.
[8] 張建明,張 玉,劉海濤.達里諾爾湖和崗更湖瓦氏雅羅魚同工酶的研究[J].水生態學雜志,2010(6):79-86.
[9] HIRATA T,KANEKO T,ONO T,et al.Mechanism of acid adaptation of a fish living in a pH 3.5 lake[J].American Journal of Physiology-Regulatory,Integrative and Comparative Physiology,2003,284(5):1199-1212.
[10] 池炳杰,常玉梅,閆學春,等.瓦氏雅羅魚達里湖群體和烏蘇里江群體的遺傳多樣性和遺傳結構分析[J].中國水產科學,2010(3):2 -17.
[11] CHANG Y M,TANG R,SUN X W,et al.Genetic analysis of population differentiation and adaptation in Leuciscus waleckii[J].Genetica,2013,141(10-12):417-429.
[12] 陳金平.綏芬河三塊魚屬(Tribolodon)魚類的分類學討論[J].水產學雜志,2001(2):021.
[13] 劉金亮,常玉梅,徐麗華,等.東北雅羅魚微衛星分子標記的篩選及特征分析[J].華北農學報,2011,26(1):87-93.
[14] ZARDOYA R,MEYER A.Phlogenetic performance of mitochondrial protein coding genes in resolving relationships among vertebrates[J].Mol Biol Ecol,1996,13(7):933-942.
[15] PERDICES A,SAYANDA D,COELHO M.Mitochondrial diversity of Opsariichthys bidens (Teleostei Cyprinidae)in three Chinese drainages[J].Mol Phylogenet Fvol,2005,37:920-927.
[16] JOHN B A,KUMAR C P,KHAN S A,et al.Efficiency of universal barcode gene (Coxi)on morphologically cryptic mugilidae fishes delineation [J].Trends in Applied Sciences Research,2011,6(9):1028-1036.
[17] 劉紅艷,熊 飛,楊 東,等.基于細胞色素b基因的鱇浪白魚野生群體和養殖群體遺傳多樣性分析[J].華中農業大學學報,2011,30(1):94-98.
[18] 孔慶鵬,羅 靜,黃順友,等.從線粒體細胞色素b 探討長臀屬三個種分類與進化的關系[J].遺傳,2000,22(6):379-384.
[19] 祁得林,晁 燕,郭松長,等.黃河裸裂尻魚五種群mtDNA 控制區的遺傳結構[J].動物學報,2008,54(6):972-980.
[20] KNIGHT A,MINDELL D P.Substitution bias,weighting of DNA sequence evolution,and the phylogenetic position of Fea's viper[J].Systematic Biology,1993,42(1):18-31.
[21] 張源真,王 偉,姜志強,等.基于16S rRNA 部分序列探討部分鳚亞目魚類的分子系統進化關系[J].Marine Sciences,
2012,36(9):89.
[22] 李 娜,陳少波,謝起浪,等.閩浙地區香魚線粒體Cyt b基因和D-loop區序列多態性分析[J].遺傳,2008,30(7):919-925.
[23] 胡文閣,段子淵,王金富,等.新疆3 種雅羅魚線粒體DNA 細胞色素b 序列的差異與系統進化[J].動物學雜志,2005,4(3):6-11.
[24] 陳星玉.中國雅羅魚亞科的骨骼系統及其分類學意義(鯉形目:鯉科)[J].動物學報,1987,12(3):311-322.
[25] WARD R D,ZEMLAK T S,INNES B H,et al.DNA barcoding Australia's fish species[J].Philosophical Transactions of the Royal Society B:Biological Sciences,2005,360(1462):1847-1857.
[26] 張鳳英,馬凌波,施兆鴻,等.兩種鯛屬魚類線粒體COI基因片段序列的比較[J].上海水產大學學報,2006,15(4):403-408.
[27] COSTA F O,DEWAARD J R,BOUTILLIER J,et al.Biological identifications through DNA barcodes:the case of the Crustacea[J].Canadian Journal of Fisheries and Aquatic Sciences,2007,64(2):272-295.
[28] 黃小彧,章 群,馬 奔,等.中華倒刺鲃線粒體細胞色素b基因的遺傳多樣性分析[J].廣東農業科學,2011,38(24):1-4.
