外來入侵害蟲棉花粉蚧雄蟲生殖能力研究
2014-12-16 01:57:12陸永躍
環境昆蟲學報 2014年5期
王 超,陳 芳,陸永躍
(華南農業大學昆蟲學系,廣州 510642)
兩性生殖是昆蟲最常見的生殖方式,在穩定的環境中兩性交配對維持其種群繁殖起到重要作用;而在不穩定的環境中,有性生殖可產生遺傳變異,造成基因多樣性,使生物有更大的機會繁衍下去,而不致滅絕(彩萬志等,2001)。許多兩性生殖的昆蟲存在多次交配現象,交配次數對一雌多雄和一雄多雌昆蟲是非常重要的。對于多數昆蟲,雌性的繁殖力受到卵子生產的限制,雌蟲在交配行為中獲得新鮮精子、促性腺激素等有用的物質(Arnqvist et al.,2000;Dunn et al.,2005;Edvardsson,2007;Yamane,2008),通過多次交配可以增加卵子的受精率,因此雌蟲交配次數決定了其繁殖的成功率(Simmons,2005);雄性能夠生產大量的精子,比與之交配的雌性全部卵子的數量更多,雄性的潛在生殖成功率受與它交配的雌性數量限制,為了避免雄蟲精子的浪費(Arnqvist et al.,2005)或是更好的基因交流(Jennions,2000;Slatyer et al.,2011),雄蟲通過多次交配提高自身的適應性,從而達到更高的交配成功率(Arnqvist et al.,2000)。在復雜多變的外界環境下,雌、雄蟲通過多次交配,對后代種群增長和進化具有重要作用。
棉花粉蚧(又名扶桑綿粉蚧)Phenacoccus solenopsis Tinsley 是2008年發現入侵中國大陸的重要害蟲(陸永躍等,2008;武三安和張潤志,2009)。該蟲寄主植物廣泛,而且隨著侵入地區變化有逐步擴大的趨勢,在巴基斯坦摩耳坦地區一處20 km 半徑范圍地點就記錄到寄主53 科154種,包括大田作物、蔬菜、觀賞植物、雜草、灌木及一些小喬木(Arif et al.,2009)。該蟲對棉花為害嚴重。2005年巴基斯坦信德省、旁遮普省、印度等地發現棉花粉蚧侵害少量棉株,隨后很快在18個棉區中的11個大面積發生,在Bt 棉和非Bt棉上均暴發成災,給當地棉花種植業造成巨大經濟損失(Anonymous,2005)。關鑫等(2011)研究了自然條件下棉花粉蚧入侵定殖能力,廣州地區6-8月番茄、棉花田新入侵的棉花粉蚧的最小入侵種群規模為36-40 頭。棉花粉蚧耐饑力較強,這也有助于它在野外建立種群(鄭婷等,2011)。對不同溫度下、不同寄主植物上發育歷期和實驗種群的研究結果表明,在適宜條件下棉花粉蚧種群趨勢指數可高達190 以上(黃芳等,2011;Lu et al.,2011;王瑩瑩等,2012;關鑫等,2012a;關鑫等,2012b;Prasad et al.,2012)。關于該蟲的生殖方式,部分認為是營兼性孤雌生殖(Fand et al.,2010;Sahito et al.,2010;Vennila et al.,2010;魏婷等,2010),但也有報道認為只有兩性生殖(Arif et al.,2012;Mamoon-ur-Rashid et al.,2012;Prasad et al.,2012)。筆者對廣東、廣西、海南、福建、上海、浙江、安徽、湖北等我國大陸11個入侵種群生殖形式進行了觀察,發現上述棉花粉蚧種群在無雄蟲情況下雌蟲不能產仔,這說明這些入侵種群僅為兩性生殖。棉花粉蚧有明顯的雌雄二型現象(Anonymous,2008;黃玲等,2011),雌成蟲個體較大,行動遲緩,雄蟲具翅、小、活動迅速,因此,交配行為主要依靠雄蟲進行(胡鳳等,2011;朱藝勇等,2011;關鑫和陸永躍,2012)。在棉花粉蚧兩性生殖種群中,雄蟲的交配能力、生殖能力對雌蟲的繁殖和種群的增長均是十分重要的。本文采用單頭雄蟲與不同數量雌蟲配比的實驗方法,研究了一頭雄蟲能滿足多少頭雌蟲的生殖需求、產生下一代種群數量和相對生殖效率等生殖生物學問題,為揭示該蟲入侵種群定殖、發生、發展等過程中雄蟲的作用提供了依據。
1 材料與方法
1.1 實驗材料
供試蟲源:棉花粉蚧采于廣州市珠江新城街道兩旁扶桑,在室內以扶桑為寄主植物進行飼養。
供試植物:扶桑幼苗種植于華南農業大學昆蟲學系實驗室,不施用任何化學藥劑滅蟲。
人工氣候箱:RXZ 型智能人工氣候箱(溫度誤差為±1℃,寧波江南儀器廠制造),設置溫度27±1℃,光周期14L∶10D,相對濕度為70±5%。
1.2 實驗方法
將棉花粉蚧一齡若蟲挑入盛有新鮮扶桑葉片的培養皿(Φ=9 cm),并置于人工氣候箱里,至二齡末時將雌、雄蟲分離飼養。取剛進入成蟲期健康的成蟲,按照性比(雌成蟲:雄成蟲)為1∶1-7∶1 比例混合后放入盛有新鮮扶桑葉片的培養皿(Φ=9 cm)中,每3 天更換一次葉片。記錄不同性比例條件下,棉花粉蚧產仔雌成蟲數量、單雌產仔量和后代性比。每個性比設30個重復。
1.3 數據分析
對實驗獲得的棉花粉蚧交配成功率、產仔雌蟲數量、總產仔量、單雌產仔量、子代性比、子代中雌蟲數量等數據進行方差分析及差異顯著性檢驗。相關指標按以下含義和公式確定。交配成功率(%)=(成功交配并產生子代的雌蟲數量/供試雌蟲數量)×100%;產仔雌蟲數量是指交配后產仔雌蟲的頭數;總產仔量指同一雌雄性比試驗中與1 頭雄蟲交配的所有雌蟲的產仔量之和;單雌產仔量(頭)=總產仔量/產仔雌蟲數量;子代性比=子代中雌蟲數量/子代中雄蟲數量;子代中雌蟲數量(頭)=總產仔量×(子代性比/(子代性比+1))。數據的分析與處理在SAS 9.0 上進行,采用Excel 軟件進行繪圖。
2 結果與分析
2.1 單頭雄蟲與雌蟲的交配成功率和產仔雌蟲數量
當棉花粉蚧1 頭雄蟲與多頭雌蟲共存時,1-3 頭雌蟲時雄蟲的交配成功率最高,為85.4%-94.4%;隨著雌蟲數量增大,雄蟲交配成功率逐漸降低,4-7 頭雌蟲時分別為80.7%、77.7%、69.5%、66.2%(表1)。
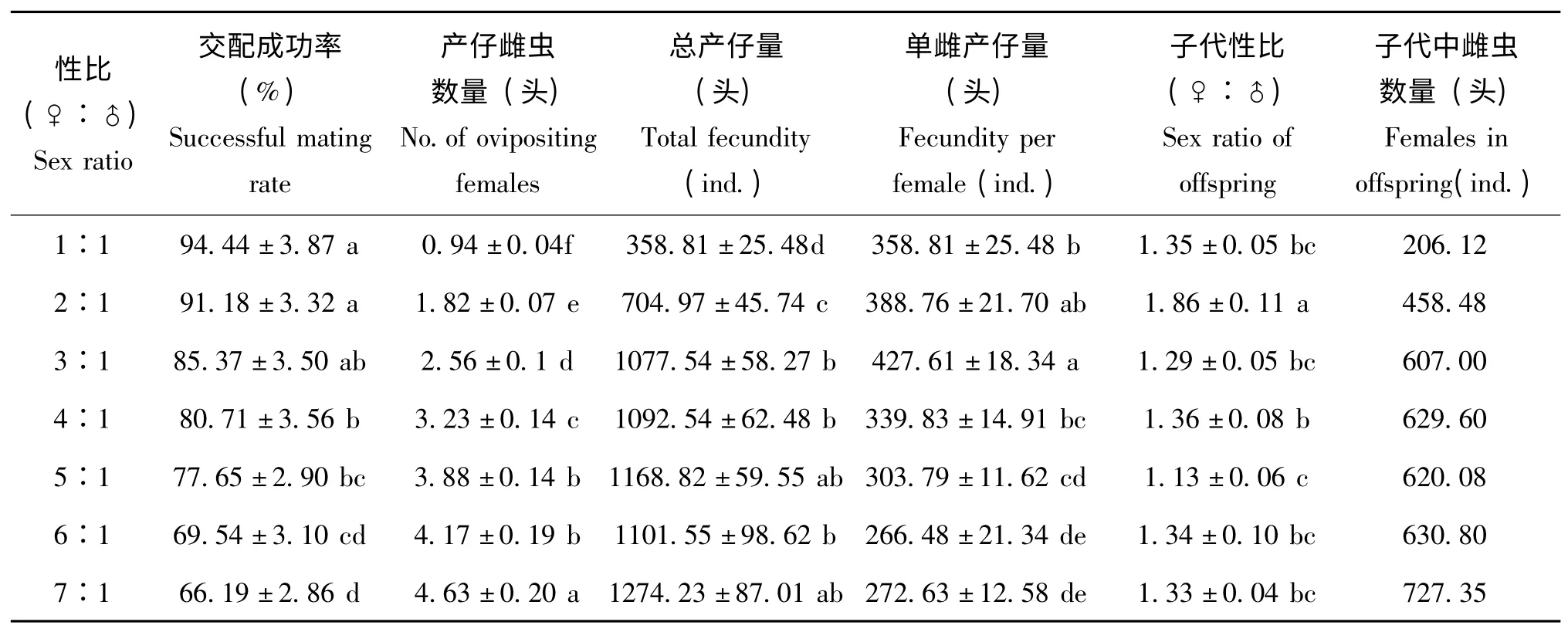
表1 不同性比條件下棉花粉蚧成蟲主要繁殖生物學參數Table 1 Parameters of reproductive biology for cotton mealybug P.solenopsis at different sex ratio
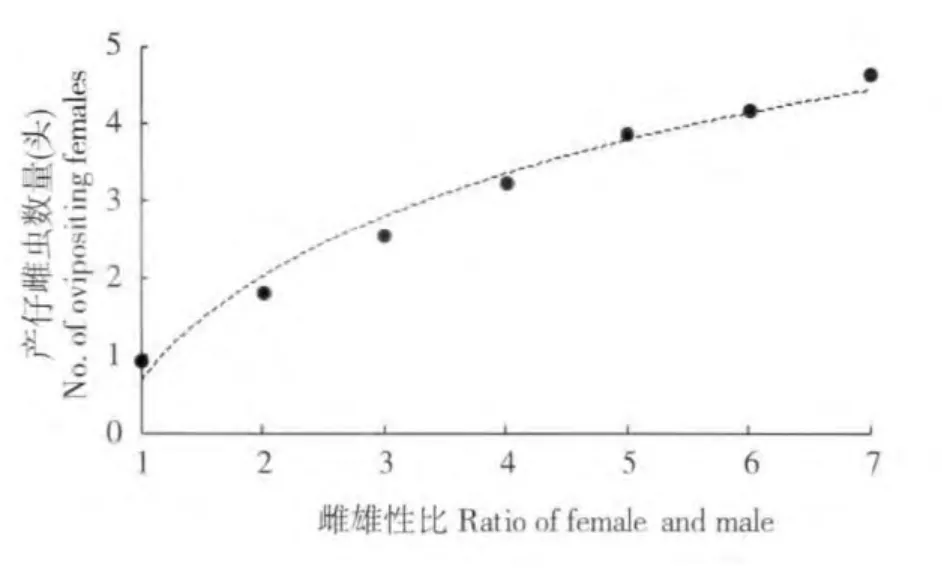
圖1 不同雌雄性比下棉花粉蚧產仔雌蟲數量增長趨勢Fig.1 Dynamic of ovipositing females number for cotton mealybug at different sex ratio
隨著雌蟲比例增大,與1 頭雄蟲成功交配并產仔的雌蟲數量也是逐漸增大的(表1)。1-7 頭雌蟲時產仔雌蟲數量分別為0.94、1.82、2.56、3.23、3.88、4.17、4.63 頭。對產仔雌蟲數量與雌雄性比的關系進行分析,發現兩者之間符合以下邏輯斯蒂曲線方程:Ff=5.2159EXP[-2.5304EXP(-0.421739N)],(R=0.9989,P=0.0001)(圖1)。式中Ff為產仔雌蟲數量(頭),N 為雌雄性比。應用該方程計算出1 頭雄蟲最多能成功交配并滿足5.22 頭雌蟲生殖后代的需求。
2.2 與單頭雄蟲交配后雌蟲產生后代數量和雄蟲的交配效率
隨著雌雄性比的增大,與單頭雄蟲交配后雌蟲總產仔量不斷增加,1∶1-3∶1 時呈快速上升趨勢,總產仔量分別為358.8 頭、705.0 頭、1077.5 頭,大于3∶1 后上升趨勢減緩,性比為7∶1時總產仔量達到1274.2 頭(表1)。雌蟲總產仔量與雌雄性比呈顯著正相關,建立了描述總產仔量與雌雄性比關系的S 型增長曲線為:Ft=1524.589e-1.3922/N(F=97.61,R=0.9876,P=0.0002)。式中Ft為雌蟲總產仔量(頭),N 為雌雄性比。通過該方程計算、獲得了與單頭雄蟲交配后雌蟲總產仔最大可達到1525 頭左右,即為1 頭雄蟲平均可能的最大生殖后代數量。在子代中,性比(♀∶♂)變化無明顯的規律,以2∶1時最大(1.86),5∶1 時最小(1.13)。根據該表中可看出,1∶1-7∶1 時與1 頭雄蟲交配的所有雌蟲所產生的子代中雌蟲數量由206.1 頭增大到727.4 頭,建立了Fof=840.1124e-1.2599/N(F=97.72,R=0.9754,P=0.0002),式中Fof為子代中雌蟲數量(頭),N 為雌雄性比。由該模型可知,與1 頭雄蟲交配后雌蟲總產仔最大可能產生840 頭雌蟲。
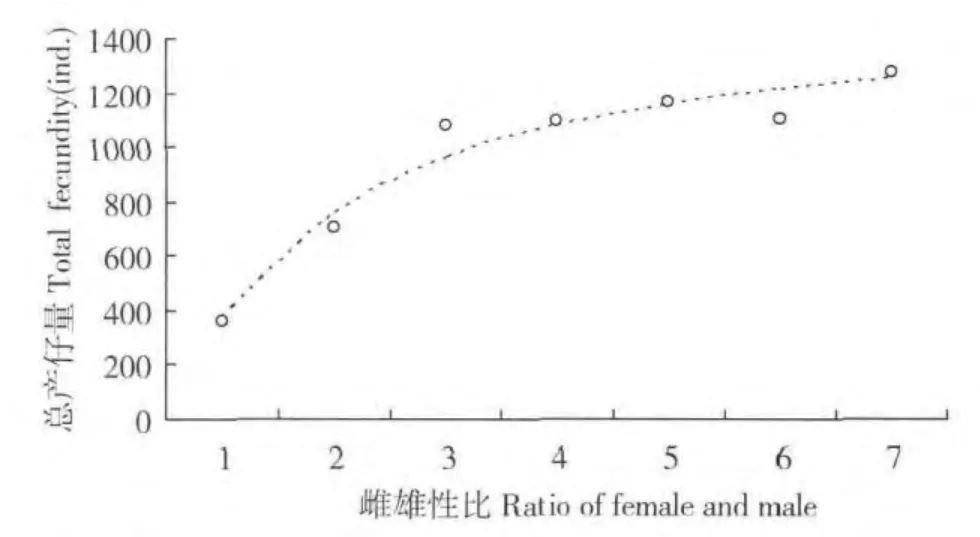
圖2 不同雌雄性比下棉花粉蚧雌蟲總產仔量變化動態Fig.2 Dynamic of total fecundity of cotton mealybug females at different sex ratio
不同雌雄性比時,與雄蟲交配成功后單雌平均產仔量是變化的,1∶1-3∶1 時呈上升趨勢,3∶1時達到最大值,為427.6 頭;4∶1-7∶1 范圍呈下降趨勢,6∶1、7∶1(♀∶♂)時最小,分別為266.5 頭、272.6 頭(表1)。這表明,不同雄蟲資源條件下雌雄蟲單次成功交配的效率是不同的,以雌雄比3∶1 時效率最高,1∶1 時下降到84%,而6∶1-7∶1 時下降到62%-63%。
3 結論與討論
從本研究結果可看出,即使是1∶1 比例時雄蟲也不會100%與雌蟲成功交配、授精,約有5%-6%比例個體(包括雌蟲、雄蟲)不會交配、不能成功交配或者交配后無法成功授精,因此導致部分雌蟲不產生后代。♀∶♂在1∶1-7∶1 范圍內,1 頭雄蟲一般可以滿足4-5 頭雌蟲生殖的需要,平均最大為5.22 頭。雄蟲進入成蟲期后,不取食,壽命明顯短于雌蟲,為確保最大限度的種群增長,雄蟲與盡可能多的雌蟲進行交配,從而促進種群增長。在不同性比條件下,雌雄蟲多次交配會提高交配的成功率,避免雄蟲精子的浪費,使粉蚧雌蟲受精囊中精子數量增多,受精卵數量也隨之增多,這使得種群基因在群體內高度雜合化,對種的生存和后代優勢的保持是十分有利的。在雄蟲數量有限的條件下,雄蟲也會選擇部分雌蟲進行交配。在前三次交配時雄蟲可以給雌蟲提供較多的精子,但隨著交配次數增加,精子不斷消耗,隨后與雄蟲交配的雌蟲獲取精子量將越來越少,直至雄蟲精子貯存數量不足或完全耗盡。例如,當雌雄性比大于3∶1 時,成功交配后的雌蟲單雌產仔量逐漸減小,所有與雄蟲交配后的雌蟲的總產仔量逐漸趨于穩定,即雄蟲交配所產生的后代總數接近或者達到了最大值,這個數量約為1525 頭。在一定性比范圍內,雄性比例小的種群后代數量將更大。例如,♀∶♂3∶1-6∶1 時所產生的子代中雌蟲數量是相近的,約為607-631 頭。
本研究表明,雌雄性別比對棉花粉蚧種群增長有著重要影響。作為外來入侵害蟲,研究棉花粉蚧雌雄性比在其入侵種群定殖、發生、發展等過程中的作用,可以為進一步了解種群的增長規律奠定基礎。通過分析棉花粉蚧產仔雌蟲數量、總產仔量、單雌產仔量、子代性比、子代中雌蟲數量與雌雄性比關系,建立定量化模型,從而預測出不同雌雄性比條件下該蟲種群的增長規律和趨勢。根據野外調查的棉花粉蚧種群數量、雌雄性比等數據,可以分析和預測種群的增長狀態,對棉花粉蚧入侵地區的風險分析和管理具有指導意義。
References)
Anonymous.Weekly Pest Scouting Reports[N].Directorate General,Pest Warning and Quality Control of Pesticides Punjab,Lahore,Pakistan,Official Reports for the months August and September,2005.
Anonymous.Annual report of the Central Cotton Research Institute,Multan.In:Pakistan Central Cotton Committee,Ministry of Food[J].Agriculture and Livestock,Islamabad,Pakistan,2008,75-76.
Arif MI,Rafiq M,Wazir S,et al.Studies on cotton mealybug,Phenacoccus solenopsis(Pseudococcidae:Homoptera),and its natural enemies in Punjab[J].Pakistan International,Journal of Agriculture and Biology,2012,14(4):557-562.
Arif MI,Rafiq M,Ghaffar A.Host plants of cotton mealybug(Phenacoccus solenopsis):a new menace to cotton agroecosystem of Punjab,Pakistan[J].International Journal of Agriculture and Biology,2009,11(2):163-167.
Arnqvist G,Nilsson T.The evolution of polyandry:multiple mating and female fitness in insects[J].Animal Behaviour,2000,60(2):145-164.
Cai WZ,Pang XF,Hua BZ,et al.General Entomology[M],Beijing:China Agricultural University Press,2001[彩萬志,龐雄飛,花保禎,等.普通昆蟲學[M].北京:中國農業大學出版社,2001]
Dunn DW,Sumner JP,Goulson D.The benefits of multiple mating to female seaweed flies,Coelopa frigida(Diptera:Coelpidae)[J].Behavioral Ecology and Sociobiology,2005,58(2):128-135.
Edvardsson M.Female Callosobruchus maculatus mate when they are thirsty:resource-rich ejaculates as mating effort in a beetle[J].Animal Behaviour,2007,74(2):183-188.
Fand BB,Gautam RD,Chander Subhash S,et al.Life table analysis of the mealybug,Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)under laboratory conditions[J].Journal of Entomological Research,2010,34(2):175-179.
Guan X,Lu YY,Zeng L.Study on developmental durations and fecundity of Phenacoccus solenopsis Tinsley on four species of hosts[J].Agricultural Science & Technology,2012,13(2):408-411.
Guan X,Lu YY,Zeng L,et al.Life Tables of Phenacoccus solenopsis Tinsley experimental population at different temperatures[J].Agricultural Science & Technology,2012,13(4):792-797,814.
Guan X,Zeng L,Lu YY.Potential of invasion and colonization for the cotton mealybug Phenacoccus solenopsis Tinsley populations in cotton and tomato fields in Guangzhou,China[J].Journal of Biosafety,2011,20(3):192-197.[關鑫,曾玲,陸永躍.廣州地區自然條件下扶桑綿粉蚧入侵定殖能力研究[J].生物安全學報,2011,20(3):192-197]
Guan X,Lu YY.Study on body length,body width and morphological characteristics of Phenacoccus solenopsis Tinsley fed on cotton[J].Journal of Anhui Agricultural Sciences,2012,40(1):261-262,298.[關鑫,陸永躍.棉花上扶桑綿粉蚧各蟲態體長、體寬與形態特征[J].安徽農業科學,2012,40(1):261-262,298]
Hu F,Liu H,Xiao TG,et al.Analysis the main factor of cotton mealybug produce male[J].Crop Research,2011,25(1):63-65.[胡鳳,劉慧,肖鐵光,等.扶桑綿粉蚧雄蟲產生的主要影響因子分析[J].作物研究,2011,25(1):63-65]
Huang L,Liu H,Ou GC,et al.Preliminary studies on biology of Phenacoccus solenopsis Tinsley,a cotton pest[J].Crop Research,2011,25(3):245-248.[黃玲,劉慧,歐高財,等.扶桑綿粉蚧部分生物學特性研究[J].作物研究,2011,25(3):245-248]
Huang F,Zhang PJ,Zhang JM,et al.Effects of three host plants on the development and reproduction of Phenacoccus solenopsis[J].Plant Protection,2011,37(4):58-62.[黃芳,張蓬軍,章金明,等.三種寄主植物對扶桑綿粉蚧發育和繁殖的影響[J].植物保護,2011,37(4):58-62]
Jennions MD,Petrie M.Why do females mate multiply?A review of the genetic benefits[J].Biological Reviews,2000,75(1):21-64.
Lu YY,Zeng L,Wang L,et al.Precaution of solenopsis mealybug Phenacoccus solenopsis Tinsley[J].Journal of Environmental Entomology,2008,30(4):386-387.[陸永躍,曾玲,王琳,等.警惕一種危險性綿粉蚧入侵中國[J].環境與昆蟲學報,2008,30(4):386-387]
Lu YY,Guan X,Zeng L.Effect of temperature on the development of the mealybug,Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)[J].Scientific Research and Essays,2011,6(31):6459-6464.
Mamoonur-Rashid M,Khattak MK,Abdullah K.Residual toxicity and biological effects of neem(Azadirachta indica)oil against cotton mealybug,Phenacoccus solenopsis Tinsley(Sternorrhyncha:Pseudococcidae)[J].Pak.J.Zool,2012,44(3):837-843.
Prasad YG,Prabhakar M,Sreedevi G,et al.Effect of temperature on development,survival and reproduction of the mealybug,Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)on cotton[J].Crop Protection,2012,39:81-88.
Sahito HA,Abro GH,Khuhro RD,et al.Biological and morphological studies of cotton mealybug Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)development under laboratory environment[J].Pakistan,The Entomological Society of Karachi,2010,25(2):131.
Simmons LW.The evolution of polyandry:sperm competition,sperm selection,and offspring viability[J].Annual Review of Ecology,Evolution,and Systematics,2005,36:125-146.
Slatyer RA,Mautz BS,Backwell PR,et al.Estimating genetic benefits of polyandry from experimental studies:a meta ‐ analysis[J].Biological Reviews,2011,87(1):1-33.
Vennila S,Deshmukh AJ,Pinjarkar D,et al.Biology of the mealybug,Phenacoccus solenopsis on cotton in the laboratory[J].Journal of Insect Science,2010,10(115):1-9.
Wang YY,Xu ZH,Zhang LL,et al.Developmental duration and life table of the laboratory population of Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)fed on pumpkin at different temperatures[J].Acta Entomologica Sinica,2012,55(1):77-83.[王瑩瑩,徐志宏,張莉麗,等.南瓜寄主上扶桑綿粉蚧不同溫度下的發育歷期和實驗種群生命表[J].昆蟲學報,2012,55(1):77-83]
Wei T,Liu H,Ou GC,et al.Preliminary studies on behavior of Phenacoccus solenopsis Tinsley[J].Crop Research,2010,24(4):358-360.[魏婷,劉慧,歐高財,等.扶桑綿粉蚧部分行為學的初步研究[J].作物研究,2010,24(4):358-360]
Wu SA,Zhang RZ.A new invasive pest,Phenacoccus solenopsis,threatening seriously to cotton production[J].Chinese Bulletin of Entomology,2009,46(1):159-162.[武三安,張潤志.威脅棉花生產的外來入侵新害蟲-扶桑綿粉蚧[J].昆蟲知識,2009,46(1):159-162]
Yamane T,Miyatake T.Strategic ejaculation and level of polyandry inCallosobruchus chinensis(Coleoptera:Bruchidae)[J].Journal of Ethology,2008,26(2):225-231.
Zheng T,Cui XH,Wang T,et al.Effects of starvation on survival rate and oviposition of the invasive mealybug Phenacoccus solenopsis(Hemiptera:Pseudococcidae)[J].Journal of Biosafety,2011,20(3)3:239-242.[鄭婷,崔旭紅,汪婷,等.饑餓對扶桑綿粉蚧存活率和產卵量的影響[J].生物安全學報,2011,20(3):239-242]
Zhu YY,Huang F,Lv YB.Bionomics of mealybug Phenacoccus solenopsis Tinsley(Hemiptera:Pseudococcidae)on cotton[J].Acta Entomologica Sinica,2011,54(2):246-252.[朱藝勇,黃芳,呂要斌.扶桑綿粉蚧生物學特性研究[J].昆蟲學報,2011,54(2):246-252]
