RNA干擾抑制EZH2基因表達(dá)對(duì)人膠質(zhì)瘤細(xì)胞凋亡及對(duì)卡莫司汀敏感性的影響
2014-12-02 04:33:52周章明楚勝華
山東醫(yī)藥 2014年34期
周章明,楚勝華
(1成都大學(xué)附屬都江堰醫(yī)療中心,四川都江堰611830;2上海交通大學(xué)附屬第三醫(yī)院)
放化療是目前常見(jiàn)的腫瘤術(shù)后治療手段。逃避凋亡是腫瘤發(fā)生、發(fā)展和產(chǎn)生藥物抵抗的重要機(jī)制[1]。誘導(dǎo)腫瘤細(xì)胞凋亡并增強(qiáng)其對(duì)化療藥物的敏感性是目前腫瘤治療的研究熱點(diǎn)。果蠅zeste基因增強(qiáng)子人類同源物2(EZH2)是多疏基因(PcG)蛋白家族的重要成員,與腫瘤細(xì)胞凋亡并在促進(jìn)癌細(xì)胞生長(zhǎng)、侵襲、轉(zhuǎn)移中起重要作用[2]。卡莫司汀可以通過(guò)血腦屏障,該藥物及其代謝物可通過(guò)烷化作用與核酸交鏈,并有可能因改變蛋白質(zhì)而產(chǎn)生抗腫瘤作用,為膠質(zhì)瘤化療的重要藥物之一[3-5]。2012年5月~2013年6月,我們用RNA干擾技術(shù)抑制人膠質(zhì)瘤U251細(xì)胞中EZH2基因的表達(dá),并檢測(cè)干擾后U251細(xì)胞對(duì)卡莫司汀的敏感性。現(xiàn)報(bào)告如下。
1 材料與方法
1.1 細(xì)胞培養(yǎng)及分組 人膠質(zhì)瘤U251細(xì)胞購(gòu)于中國(guó)科學(xué)院細(xì)胞所。細(xì)胞培養(yǎng)于含有10%新生小牛血清的RPMI-1640培養(yǎng)基中,內(nèi)加青霉素1×105U/mL,鏈霉素 100 mg/mL,置于 5%CO2、飽和濕度、37℃恒溫培養(yǎng)箱內(nèi)培養(yǎng)。分為空白對(duì)照組、脂質(zhì)體組、陰性對(duì)照組和pRNAT-EZH2組。
1.2 小干擾RNA(siRNA)真核表達(dá)載體的構(gòu)建具有互補(bǔ)序列的能夠編碼短發(fā)卡RNA(shRNA)的雙鏈寡核苷酸模板DNA由上海康成公司合成。其轉(zhuǎn)錄產(chǎn)物所形成的siRNA的作用靶點(diǎn)為人EZH2 mRNA。干擾序列為:AAGACTCTGAATGCAGTTGCT;在兩端分別懸垂Sal I、Xba I的酶切位點(diǎn),3'端酶切位點(diǎn)之前加入終止信號(hào)TTTT;陰性對(duì)照是雙鏈寡核苷酸轉(zhuǎn)錄產(chǎn)物所形成的siRNA,此序列不與任何人類基因序列同源。
1.3 細(xì)胞轉(zhuǎn)染 用Lipofectin-2000TM(美國(guó)Invitrogene公司)介導(dǎo)進(jìn)行轉(zhuǎn)染。空白對(duì)照組不轉(zhuǎn)染。脂質(zhì)體組轉(zhuǎn)染液中僅含脂質(zhì)體無(wú)siRNA。陰性對(duì)照組轉(zhuǎn)染液加入空質(zhì)粒pRNAT-Negative。pRNAT-EZH2組轉(zhuǎn)染液加入重組pRNAT-EZH2質(zhì)粒。
1.4 U251細(xì)胞中EZH2 mRNA檢測(cè)方法 收集轉(zhuǎn)染24、48、72 h各組細(xì)胞,用 Trizol(美國(guó) Invitrogene公司)提取細(xì)胞總RNA,采用RT-PCR法檢測(cè)EZH2 mRNA,操作按試劑盒說(shuō)明書(shū)進(jìn)行。EZH2引物序列:上游 5'-GCCAGACTGGGAAGAAATCTG-3',下游3'-TGTGCTGGAAAATCCAAGTCA-5',產(chǎn)物大小為86 bp。β-actin引物序列:上游 5'-ATCTGGCACCAAACACCTTCTACAATGAGCTGCG-3',下游5'-CGTCATACTCCTGCTTGCTGATCCACATCTGC-3',產(chǎn)物為167 bp。以EZH2與β-actin條帶的積分吸光度比值表示EZH2 mRNA的相對(duì)表達(dá)量。
1.5 卡莫司汀處理后各組細(xì)胞凋亡檢測(cè)方法 將轉(zhuǎn)染后48 h的細(xì)胞以2×105個(gè)/mL的密度接種在6孔培養(yǎng)板中,分別加入含卡莫司汀 10、25、50 μmol/L的完全培養(yǎng)基,每組設(shè)3個(gè)復(fù)孔,培養(yǎng)24、48 h后小心吸去培養(yǎng)液,加入0.5 mL細(xì)胞固定液,室溫下固定10 min。吸去固定液,用PBS洗兩遍,洗滌時(shí)用搖床晃動(dòng),每次3 min,吸盡液體。加入0.5 mL Hoechst 33258染色液,染色5 min。PBS洗兩遍,每次3 min,吸盡液體。滴一滴抗熒光淬滅封片液于載玻片上,蓋上貼有細(xì)胞的蓋玻片,盡量避免汽泡,使細(xì)胞接觸封片液。用激光共聚焦顯微鏡觀察細(xì)胞凋亡情況,激發(fā)波長(zhǎng)352 nm,發(fā)射波長(zhǎng)461 nm。
1.6 卡莫司汀處理后各組細(xì)胞增殖抑制情況檢測(cè)方法 采用MTT法。取pRNAT-EZH2組、陰性對(duì)照組、空白對(duì)照組的對(duì)數(shù)生長(zhǎng)期細(xì)胞接種于96孔培養(yǎng)板內(nèi),每孔加入細(xì)胞1×104個(gè),24 h后加入含卡莫司汀10、25、50 μmol/L 的完全培養(yǎng)基,每組設(shè)3 個(gè)復(fù)孔,繼續(xù)作用24 h,每孔加入20 μL MTT(5 mg/mL)繼續(xù)培養(yǎng)4 h,吸棄去培養(yǎng)液,加入DMSO 150 μL,酶標(biāo)儀測(cè)定570 nm處吸光度(A)值。取3孔平均A值。計(jì)算細(xì)胞增殖抑制率。
1.7 Caspase-3檢測(cè)方法 取 pRNAT-EZH2組、陰性對(duì)照組、空白對(duì)照組的對(duì)數(shù)生長(zhǎng)期細(xì)胞,24 h后加入含卡莫司汀 0、10、25、50μmol/L 的完全培養(yǎng)基,加入細(xì)胞裂解液裂解,用分光光度計(jì)檢測(cè)Caspase-3水平。操作按試劑盒說(shuō)明進(jìn)行。
1.8 統(tǒng)計(jì)學(xué)方法 采用SPSS11.5統(tǒng)計(jì)軟件。計(jì)量資料以±s表示,組間比采用方差分析。P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 EZH2 mRNA 轉(zhuǎn)染72 h,空白對(duì)照組、脂質(zhì)體組、陰性對(duì)照組、pRNAT-EZH2組 U251細(xì)胞EZH2 mRNA相對(duì)表達(dá)量分別為 0.597±0.046、0.603 ±0.031、0.428 ± 0.047、0.151 ± 0.008。pRNAT-EZH2組與空白對(duì)照組相比,P<0.05,空白對(duì)照組與陰性對(duì)照組及脂質(zhì)體組相比,P均>0.05。pRNAT-EZH2組轉(zhuǎn)染24、48、72h時(shí) EZH2 mRNA 表達(dá)抑制率分別為58.19%、73.37%和74.58%。
2.2 細(xì)胞凋亡情況 空白對(duì)照組及陰性對(duì)照組細(xì)胞經(jīng)Hoechst 33258染色后,激光共聚焦顯微鏡下觀察見(jiàn)細(xì)胞均勻藍(lán)染,脂質(zhì)體組和pRNAT-EZH2組細(xì)胞則檢測(cè)出明顯的細(xì)胞凋亡,細(xì)胞核或細(xì)胞質(zhì)內(nèi)可見(jiàn)濃染致密的顆粒狀藍(lán)色熒光及明顯的熒光碎片,pRNAT-EZH2組凋亡細(xì)胞最多。
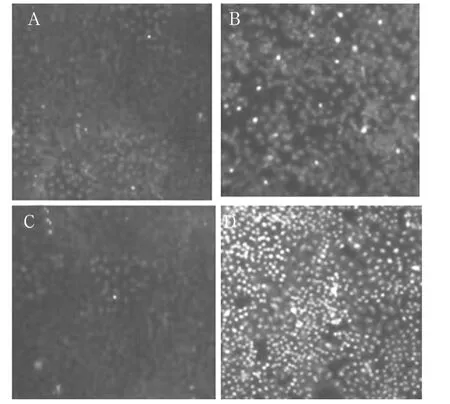
圖1 各組細(xì)胞凋亡情況激光共聚焦顯微鏡觀察結(jié)果
2.3 細(xì)胞增殖抑制率和Caspase-3相對(duì)表達(dá)量pRNAT-EZH2轉(zhuǎn)染后,U251細(xì)胞增殖抑制率和Caspase-3相對(duì)表達(dá)量明顯高于空白對(duì)照組和陰性對(duì)照組(P均<0.01),而空白對(duì)照組和陰性對(duì)照組的生長(zhǎng)抑制率差異無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05)。詳見(jiàn)表1。
表1 不同濃度卡莫司汀處理后各組U251細(xì)胞增殖抑制率和Caspase-3相對(duì)表達(dá)量(±s)

表1 不同濃度卡莫司汀處理后各組U251細(xì)胞增殖抑制率和Caspase-3相對(duì)表達(dá)量(±s)
注:與空白對(duì)照組和陰性對(duì)照組相比,*P<0.05
組別 n 細(xì)胞增殖抑制率(%)Caspase-3相對(duì)表達(dá)量空白對(duì)照組3 0 μmol/L 1.00 ±0.05 10 μmol/L 2.21 ±0.32 1.59 ±0.27 25 μmol/L 17.43 ±3.51 3.97 ±0.36 50 μmol/L 42.32 ±4.27 7.85 ±0.17陰性對(duì)照組 3 0 μmol/L 1.01 ±0.06 10 μmol/L 2.39 ±1.01 1.48 ±0.32 25 μmol/L 19.28 ±3.59 3.89 ±0.14 50 μmol/L 45.12 ±4.39 7.64 ±0.27 pRNAT-EZH2組 3 0 μmol/L 1.93 ±0.04*10 μmol/L 15.34 ±1.72* 2.17 ±0.03*25 μmol/L 36.49 ±4.13* 4.91 ±0.06*50 μmol/L 67.16 ±4.57* 10.12 ±0.03*
3 討論
膠質(zhì)瘤具有高侵襲性,局部復(fù)發(fā)轉(zhuǎn)移快,并對(duì)已知的化療藥物基本耐藥。術(shù)后化療在膠質(zhì)瘤的綜合治療中占有重要地位,多藥耐藥是影響治療效果的重要原因[6,7]。化學(xué)藥物所致細(xì)胞毒作用的終末效應(yīng)均為誘導(dǎo)細(xì)胞凋亡[8],激活細(xì)胞凋亡的能力是抗腫瘤藥物療效的主要檢測(cè)標(biāo)準(zhǔn)[9],由于腫瘤細(xì)胞常存在凋亡信號(hào)轉(zhuǎn)導(dǎo)途徑缺陷,導(dǎo)致許多腫瘤對(duì)化療藥物產(chǎn)生耐受[10]。因此通過(guò)沉默癌基因及活化凋亡信號(hào)關(guān)鍵分子而促進(jìn)細(xì)胞凋亡,為提高腫瘤化療敏感性提供了新思路。
EZH2是凋亡抑制蛋白家族中一類抑制凋亡、調(diào)節(jié)細(xì)胞分化的細(xì)胞因子[2]。目前已有研究發(fā)現(xiàn)在前列腺癌、胃腺、直腸癌等高度惡性轉(zhuǎn)移性腫瘤中EZH2 過(guò)度表達(dá)[2,11~14]。RNA 干擾技術(shù)是通過(guò)小的雙鏈RNA阻斷體內(nèi)特定基因的表達(dá),促使mRNA降解,誘使細(xì)胞表現(xiàn)出特定基因缺失表達(dá)的技術(shù),是目前沉默某些致病基因功能的高效及特異性手段。本研究構(gòu)建了針對(duì)EZH2基因的siRNA真核表達(dá)載體,并用其轉(zhuǎn)染人膠質(zhì)瘤U251細(xì)胞,結(jié)果顯示pRNAT-EZH2轉(zhuǎn)染U251細(xì)胞后顯著抑制了EZH2 mRNA表達(dá),提示本研究構(gòu)建的siRNA真核表達(dá)載體的確能夠在細(xì)胞內(nèi)持續(xù)地表達(dá)siRNA,高效、特異的抑制EZH2基因的表達(dá)。
進(jìn)一步研究發(fā)現(xiàn)空白對(duì)照組及陰性對(duì)照組細(xì)胞經(jīng)Hoechst 33258染色后,激光共聚焦顯微鏡下呈現(xiàn)的細(xì)胞均勻藍(lán)染,而卡莫司汀組及卡莫司汀加pRNAT-EZH2組細(xì)胞則檢測(cè)出明顯的細(xì)胞凋亡,細(xì)胞核或細(xì)胞質(zhì)內(nèi)可見(jiàn)濃染致密的顆粒狀藍(lán)色熒光及明顯的熒光碎片,其中可見(jiàn)卡莫司汀加pRNAT-EZH2組細(xì)胞凋亡最多。可以證實(shí)轉(zhuǎn)染EZH2 siRNA促進(jìn)了膠質(zhì)瘤細(xì)胞凋亡,在與卡莫司汀聯(lián)合作用下,凋亡更加明顯。
本研究結(jié)果顯示,穩(wěn)定轉(zhuǎn)染EZH2 siRNA真核表達(dá)載體后卡莫司汀對(duì)細(xì)胞生長(zhǎng)抑制作用增加,pRNAT-EZH2轉(zhuǎn)染U251細(xì)胞后增殖抑制率明顯高于空白對(duì)照組和陰性對(duì)照組,卡莫司汀加pRNATEZH2組細(xì)胞增殖抑制尤為明顯。Caspase家族在介導(dǎo)細(xì)胞凋亡的過(guò)程中起著非常重要的作用,其中Caspase-3為關(guān)鍵的執(zhí)行分子,它在凋亡信號(hào)傳導(dǎo)的許多途徑中發(fā)揮功能[15]。穩(wěn)定轉(zhuǎn)染 EZH2 siRNA載體后Caspase-3活性明顯升高,以卡莫司汀加pRNAT-EZH2組細(xì)胞升高最為顯著,表明用RNA干擾技術(shù)抑制EZH2表達(dá)造成腫瘤細(xì)胞對(duì)卡莫司汀的耐受性降低。
過(guò)多表達(dá)EZH2可能并非激活腫瘤細(xì)胞化療藥物的耐受性和抗凋亡能力所必需的因素,但抑制EZH2表達(dá)可能作為有效的調(diào)控信號(hào),提高腫瘤細(xì)胞對(duì)卡莫司汀化療敏感性,導(dǎo)致腫瘤細(xì)胞凋亡。本研究中筆者通過(guò)應(yīng)用RNA干擾技術(shù)成功增加了人膠質(zhì)瘤細(xì)胞對(duì)卡莫司汀的敏感性,應(yīng)引起重視。
[1]Bradley D,Rees J.Updates in the management of high-grade glioma[J].J Neurol,2014,261(4):651-654.
[2]Yamaguchi H,Hung MC.Regulation and Role of EZH2 in Cancer[J].Cancer Res Treat,2014,46(3):209-222.
[3]Mack F,Schufer N,Kebir S,et al.Carmustine(BCNU)plus Teniposide(VM26)in Recurrent Malignant Glioma[J].Oncology,2014,86(5-6):369-372.
[4]Ishikawa E,Yamamoto T,Satomi K,et al.Intraoperative pathological diagnosis in 205 glioma patients in the pre-BCNU wafer era:retrospective analysis with intraoperative implantation of BCNU wafers in mind[J].Brain Tumor Pathol,2014,31(3):156-161.
[5]Yi N,Oh B,Kim HA,et al.Combined delivery of BCNU and VEGF siRNA using amphiphilic peptides for glioblastoma[J].J Drug Target,2014,22(2):156-164.
[6]Weller M,Pfister SM,Wick W,et al.Molecular neuro-oncology in clinical practice:a new horizon[J].Lancet Oncol,2013,14(9):370-379.
[7]Kim S,Jo S,Lee H,et al.Lobarstin enhances chemosensitivity in human glioblastoma T98G cells[J].Anticancer Res,2013,33(12):5445-5451.
[8]Schmitt CA.Senescence,apoptosis and therapy-cutting the lifelines of cancer[J].Nat Rev Cancer,2003,32:286-295.
[9]Chen N,Chen CC,Lau LF.Adhesion of human skin fibroblasts to EZH2 is mediated through integrin α6β1 and cell surface heparan sulfate proteoglycans[J].J Biol Chem,2000,275(3):24953-24961.
[10]Qiu S,Huang D,Yin D,et al.Suppression of tumorigenicity by microRNA-138 through inhibition of EZH2-CDK4/6-pRb-E2F1 signal loop in glioblastoma multiforme[J].Biochim Biophys Acta,2013,1832(10):1697-1707.
[11]McCabe MT,Creasy CL.EZH2 as a potential target in cancer therapy[J].Epigenomics,2014,6(3):341-51.
[12]Debeb BG,Gong Y,Atkinson RL,et al.EZH2 expression correlates with locoregional recurrence after radiation in inflammatory breast cancer[J].J Exp Clin Cancer Res,2014,33(1):58.
[13]Wee ZN,Li Z,Lee PL,et al.EZH2-mediated inactivation of IFN-γ-JAK-STAT1 signaling is an effective therapeutic target in MYC-driven prostate cancer[J].Cell Rep,2014,8(1):204-16.
[14]Cavalli G.Molecular biology.EZH2 goes solo.Science.2012;338(6113):1430-1431.
[15]D??rr JR,Yu Y,Milanovic M,Beuster G,et,al.Synthetic lethal metabolic targeting of cellular senescence in cancer therapy[J].Nature,2013,501(7467):421-425.
