H2O2脅迫下豌豆初生根及抗氧化酶系統對外源Ca2+的響應
2014-11-07 02:08:04劉會杰李勝馬紹英張品南時振振楊曉明
草業學報 2014年6期
劉會杰,李勝*,馬紹英,張品南,時振振,楊曉明
(1.甘肅省干旱生境作物學重點實驗室 甘肅農業大學生命科學技術學院,甘肅 蘭州 730070;2.甘肅省農業科學院作物研究所,甘肅 蘭州 730070)
豌豆(Pisumsativum)屬豆科(Fabaceae)蝶形花亞科(Faboideae)豌豆屬(Pisum)一年生攀緣草本植物[1]。它是糧菜飼兼用作物,小雜糧中主要的食用豆,具有極大的農業潛在價值。豌豆還具有很高的生物固氮能力,有助于富集土壤中的營養肥料減少生產成本,所以對可持續農業的發展起著重要的作用[2]。然而,我國西北地區淡水資源缺乏,土壤鹽漬化日趨嚴重[3-4],并且伴隨著采礦、冶金以及鎘(Cd)處理等工業的發展,我國農業土壤Cd污染問題也不斷加劇。Cd是毒性最強的重金屬元素之一,被植物吸收后大部分富集在根部,從而抑制根系的正常生長[5],限制豆科植物與根瘤菌共生關系的建立,導致作物產量和品質急劇降低,威脅著農業生產和生態環境,因此對鹽漬土及Cd污染土壤的治理已經引起國內外的廣泛重視[6]。
隨著生物膜理論和研究技術的發展,越來越多的研究證實了生物膜在植物逆境脅迫中的重要性。研究表明膜傷害與植物低溫脅迫、水分脅迫、鹽脅迫和環境污染物脅迫(重金屬污染如Cd)有密切關系[7]。王芳等[8]研究指出100 mg/L的Cd處理使膜脂過氧化產物丙二醛(MDA)含量增加,質膜透性增大,Cd又能導致擬南芥(Arabidopsisthaliana)根尖H2O2的積累[9],發生膜脂過氧化傷害,同時鹽脅迫導致黃瓜(Cucumissativus)根系MDA含量和電解質滲漏率增加,生物膜受到傷害[10]。大量實驗表明,各種逆境脅迫對植物的傷害首先表現為生物膜的傷害,而脅迫中產生的羥基自由基(·OH)是造成膜脂過氧化作用的主要因素。植物在逆境脅迫過程中會產生大量的活性氧(ROS)[11-12],主要是H2O2能通過Harber-weiss反應產生更活躍、毒性更強的羥基自由基(·OH)[13],當其產生速度超出細胞抗氧化防御系統的清除能力時,就會引發一系列毒性效應,造成對細胞的“氧化脅迫”,從而引發或加劇膜脂過氧化作用,造成細胞膜系統的損傷,嚴重時導致細胞衰老或死亡。因此可以認為植物細胞內H2O2的累積是植物逆境脅迫過程中發揮作用的重要因素,減緩植物細胞內H2O2積累降低質膜傷害程度是研究植物抗逆性的首要任務。那么施加外源H2O2就相當于對植物細胞進行直接的過氧化脅迫也是對高鹽高鎘脅迫的模擬,對外源H2O2脅迫產生緩解效應的物質則能直接緩解各種逆境脅迫對植物的傷害。
本試驗初期發現外源H2O2脅迫抑制了豌豆初生根的正常向地性生長并導致水平彎曲,機理尚不明確,猜想可能是高濃度H2O2對豌豆根系生物膜產生了氧化迫害。Demidchik等[14]研究發現外源H2O2處理可導致擬南芥根部Ca2+瞬間大量涌入。張春平等[15]指出外源Ca2+可提高紫蘇(Perillafrutescens)幼苗葉片中抗氧化物酶的活性,減輕氧化損傷,并顯著緩解鹽脅迫對紫蘇生長的抑制作用,從而提高植株的耐鹽性。也有研究表明外源加入Ca2+可拮抗Cd對毛狀根生長的抑制或毒害,減少毛狀根對Cd的吸收和吸附[16]。但關于外源Ca2+對H2O2脅迫下豌豆初生根生長的相關研究報道甚少。本研究以豌豆“隴豌一號”為材料,在高濃度H2O2脅迫下施加不同濃度的外源Ca2+,測定了外源Ca2+對H2O2脅迫下的豌豆初生根膜脂過氧化指標及相關的抗氧化酶系統變化,探討不同濃度Ca2+對H2O2脅迫下初生根膜脂過氧化的緩解效應,尋求Ca2+對H2O2脅迫的緩解調節機制,以期為抗逆境脅迫的調控方法提供理論依據,同時為鹽漬土及鎘污染土壤的改良提供方法。
1 材料與方法
1.1 試驗材料
試驗于2013年4-7月在甘肅農業大學植物細胞工程實驗室進行。供試材料為早熟、矮莖、半無葉型豌豆新品種“隴豌一號”,由甘肅省農業科學院作物所提供。H2O2和Ca2+的供體分別為30% H2O2和CaCl2,2種供體均為國產分析純(天津光復精細化工研究所)。
1.2 試驗處理
挑選籽粒飽滿、均勻一致的豌豆種子,消毒后用蒸餾水沖洗3~4次,然后用吸水紙吸干。將處理好的種子放入直徑為15 cm且底部鋪有紗布的大培養皿中,于培養箱中(26±2)℃下進行培養,光照時間為24 h,光照強度為2000 lx。每個培養皿60粒種子,3次重復,每12 h更換處理液,處理液濃度如表1,CK組只加蒸餾水。以上所有的豌豆種子均水平放置在培養皿的底部,即胚根的方向與培養皿的底面相平行。脅迫所用的H2O2濃度為預試驗得出的80 mmol/L的適宜濃度。
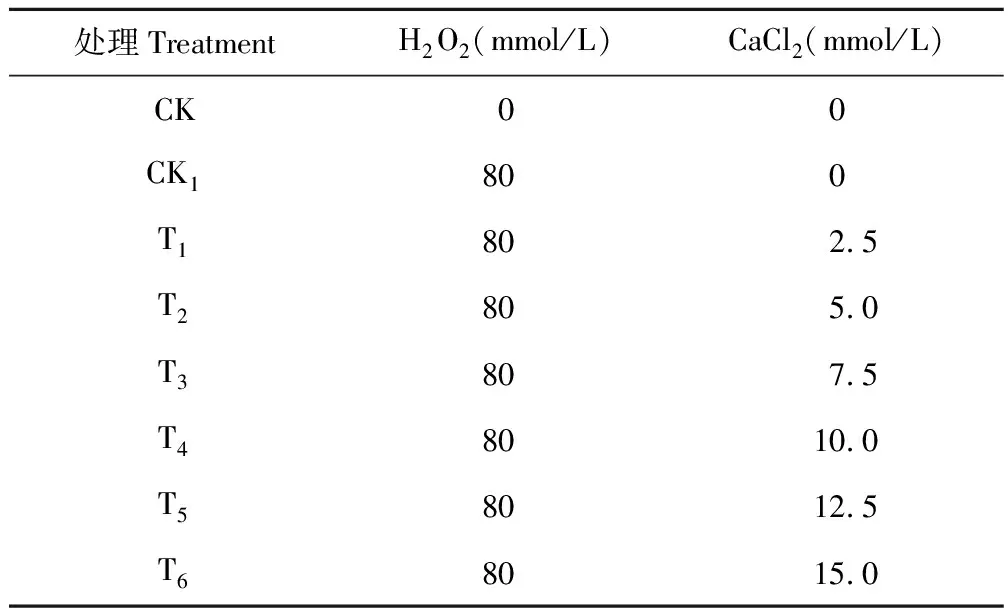
表1 試驗處理Table 1 Treatments of the experiment
1.3 幼根相關生理指標的測定
豌豆種子萌發72 h后拍照,進行數據統計后將幼苗從培養皿中取出,根部用蒸餾水清洗,測定不同處理下初生根的各項生理指標。
用TTC法測定初生根根系活力[17],雙酶法[18]測定初生根內源H2O2含量。丙二醛(MDA)含量參照Velikova等[19]的硫代巴比妥酸(TBA)法檢測。相對電導率的測定參照李合生[20]的方法。
酶液的制備:分別取各處理的初生根液氮研磨后,加入5 mL 50 mmol/L預冷的磷酸緩沖液(pH 7.8),轉入離心管中在4℃下離心20 min,上清液即為待測酶液。
超氧化物歧化酶(SOD)采用氮藍四唑(NBT)法檢測[21],以抑制NBT光化還原50%所需的酶量為1個酶活力單位(U)。過氧化氫酶(CAT)活性檢測采用鉬酸銨比色法進行測定[22]。借鑒陸曉民等[23]的方法測定過氧化物酶(POD)活性,以每分鐘OD值變化0.01為1個酶活性單位(U)。抗壞血酸過氧化物酶(APX)的測定參照Mishra等[24]的方法。每種指標每個處理重復3次。
1.4 數據處理
用Excel軟件處理數據和繪圖,SPSS 19.0軟件進行統計分析,用Duncan’s新復極差法進行差異顯著性檢驗(P<0.05)。
2 結果與分析
2.1 不同處理對豌豆初生根生長的影響
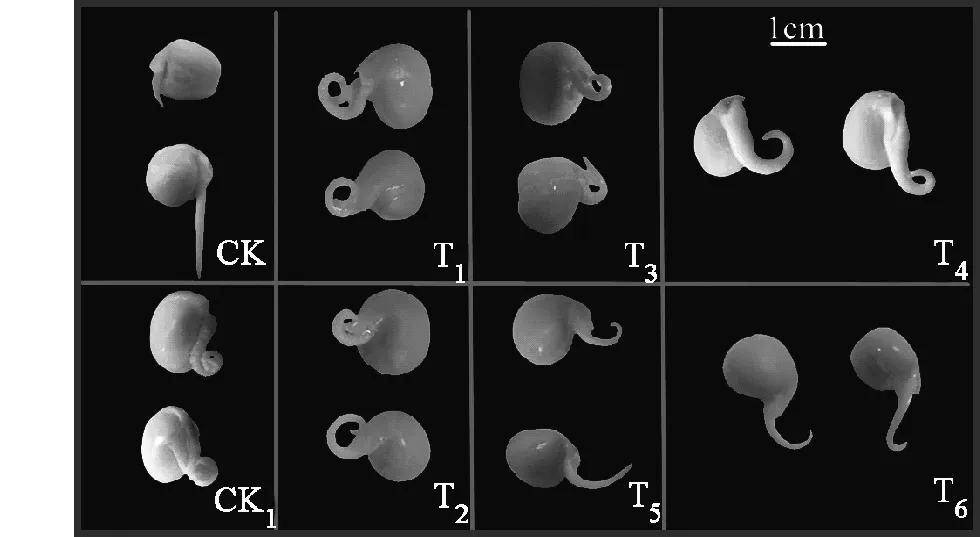
圖1 不同處理對豌豆初生根發生水平彎曲的效應Fig.1 Effect of different treatments on horizontal bending of pea primary roots
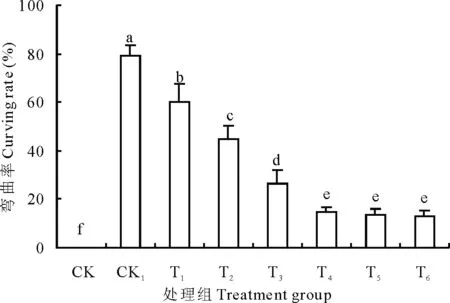
圖2 不同處理對豌豆初生根彎曲率的影響Fig.2 Effects of different treatments on curving rate of pea primary roots 不同小寫字母代表差異顯著(P<0.05),下同。Different small letters indicate statistically significant differences according to test (P<0.05). The same below.
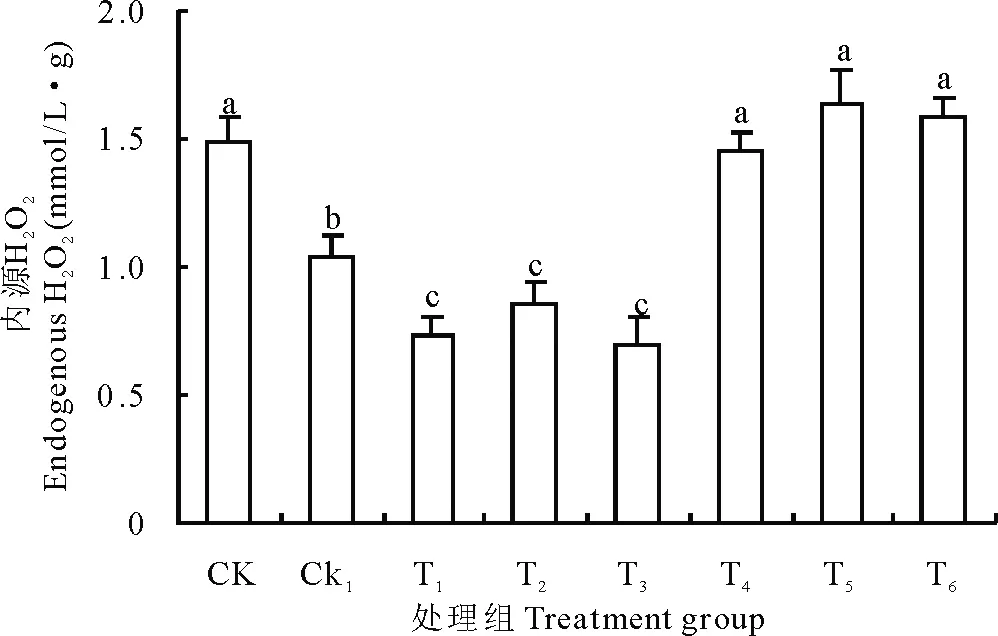
圖3 不同處理對豌豆初生根內源H2O2的影響Fig.3 Effects of different treatments on endogenous H2O2 of pea primary roots
將清洗干凈的豌豆種子水平放置在盛有蒸餾水的培養皿中培養24 h時,豌豆種子開始萌發,胚根開始突破種皮,由于重力作用,豌豆的初生根開始沿著重力的方向發生向地性彎曲(圖1,CK)。然而,如果將豌豆種子同樣水平放置在盛有H2O2的培養皿中,則向地性生長完全受到抑制,而發生水平方向的彎曲,這種彎曲的方式,不同于向地性的生長。試驗中用高濃度(80 mmol/L)H2O2處理的豌豆種子,初生根72 h后水平彎曲弧度大于360度(圖1,CK1)。若在施加外源H2O2(80 mmol/L)的前提下同時加入不同濃度的Ca2+,這種水平彎曲的現象將會得到緩解,并且隨著Ca2+濃度的增加緩解效應越明顯(圖1)。
試驗中以彎曲大于90度以上的初生根數量占初生根總數的百分比計算初生根的彎曲率。結合圖2更能說明Ca2+可以緩解外源H2O2誘導的豌豆初生根水平彎曲的現象。從圖2中可知,在CK1處理下,72 h后初生根彎曲率高達79.13%,隨著外源Ca2+濃度的增加,初生根的彎曲率顯著降低。Ca2+濃度增加到10 mmol/L時(T4),初生根彎曲率降低為14.74%,是外源H2O2脅迫下(CK1,79.13%)的1/5。在Ca2+濃度大于10 mmol/L后緩解趨勢減弱,差異不顯著。
2.2 不同處理對豌豆初生根內源H2O2的影響
在正常的狀態下,清除系統會保持植物細胞內H2O2產生和清除的一種動態平衡。但是,當遇到環境的脅迫時如鹽、干旱、低溫等,H2O2的含量會快速升高[25]。為了研究外源H2O2處理后,是否引起了豌豆初生根細胞內H2O2水平的變化,采用雙酶法定量檢測了 H2O2含量。結果發現,與CK組相比,CK1處理內源的H2O2含量顯著降低,即CK1初生根的內源H2O2含量(1.034 mmol/L·g)明顯低于CK(1.4913 mmol/L·g)。但是豌豆種子浸泡在施加了外源Ca2+的培養皿中72 h后,因施加濃度的不同變化也各有差異。當施加的外源Ca2+濃度從2.5 mmol/L到7.5 mmol/L的處理中初生根的內源H2O2水平無顯著差異,但顯著低于其他處理。當Ca2+濃度大于10 mmol/L時,初生根內源H2O2含量則顯著高于CK1和低濃度Ca2+處理,較CK差異不顯著。試驗結果說明,外施Ca2+能緩解H2O2脅迫下造成的內源H2O2含量的降低,當外施Ca2+達到一定濃度時,初生根內源H2O2的產生可恢復至正常生長水平(圖3)。
2.3 不同處理對豌豆初生根根系活力的影響
眾所周知,在正常的植物生長代謝中,H2O2作為代謝的副產物會源源不斷地產生,因此內源H2O2的產生與植物組織器官的代謝和生長活力緊密相關。有些實驗表明,當植物在遭受外界因子的脅迫或者刺激時,內源H2O2水平會迅速的升高,轉換成信號進行細胞調節[26]。為什么在本試驗中外源H2O2處理后內源H2O2水平會降低,而在施加一定高濃度的外源Ca2+后又重新升高呢,為了闡明這一問題,用TTC法測定了初生根的根活力。根的活力是一種能夠反映植物生長代謝旺盛程度的一個重要指標。無色的TTC在接受細胞中線粒體上脫氫酶傳遞的電子后轉變為粉紅色的TF[27-28]。因此根的活力與初生根的TF染色程度或者TF的含量呈正相關。
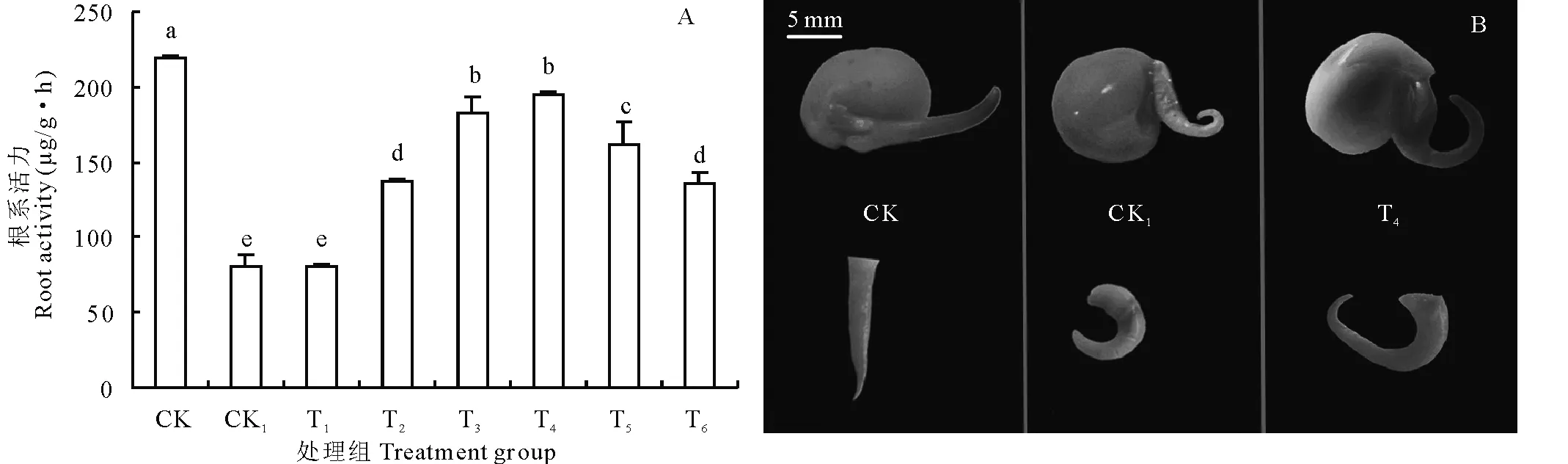
圖4 不同處理對豌豆初生根根系活力的影響Fig.4 Effects of different treatments on root activity of pea primary roots A:不同處理下初生根根系活力值;B:不同處理下初生根TTC法染色效果。 A:Values of primary root activity under different treatments; B:Dyeing effect of TTC on pea primary root under different treatments.
檢測結果如圖4所示,外源H2O2降低豌豆初生根的根系活力,而同時施加外源Ca2+時根系活力又隨著Ca2+濃度的增加而增強,體現了其緩解效應,如圖4A,而染色反應也印證了這一結果(圖4B)。從圖4A可看出,正常生長情況下(CK)初生根的根系活力達到219.225 μg/(g·h),是H2O2處理63.429 μg/(g·h)的3.45倍。當Ca2+濃度為2.5 mmol/L時豌豆初生根根系活力并沒有得到顯著提高,隨著外源Ca2+濃度的增加根系活力逐漸恢復,當Ca2+濃度增大至10 mmol/L時(T4)初生根的根系活力達到194.849 μg/(g·h),但與T3相比并不明顯,也顯著低于CK。當Ca2+濃度大于10 mmol/L時,根系活力又顯著降低,在Ca2+濃度為15 mmol/L (T6)時根系活力降低為135.779 μg/(g·h),雖然高于CK1,但顯著低于CK、T3和T4。
圖4B中可以觀察到,CK培養的豌豆種子初生根活力最強,整個初生根幾乎都被染成了TF的粉紅色。當用H2O2處理72 h后,初生根的根系活力明顯下降,即染色非常淺或者染不上色(圖4B,CK1)。在施加高濃度的Ca2+后,初生根又能夠被TTC染上色,這表明根系活力得到恢復,但結合圖4A的數據分析仍明顯低于CK。在植物的正常生長代謝中,H2O2作為信號因子不斷地產生,參與植物組織器官的代謝。本試驗結果表明,正常處理的豌豆初生根生長過程中,根的活力強,代謝旺盛,H2O2作為信號因子產生參與細胞的調控,因此內源的H2O2含量高(圖3);反之,外源H2O2處理后,初生根活力明顯受到抑制,因而使初生根自身內源H2O2產生含量降低,恢復后根的活力又明顯地反彈。TTC染色的實驗結果與定量檢測的結果一致(圖4B)。
2.4 不同處理對豌豆初生根膜脂過氧化和質膜透性的影響
膜脂過氧化產物 MDA 含量及相對膜透性是細胞膜受損傷程度的重要指標。由圖5可知,不論是在單獨H2O2處理下或者同時施加Ca2+處理,培養72 h后的豌豆初生根中MDA含量和相對電導率均比同期CK組升高,但是外施Ca2+降低了MDA含量和相對電導率的增加幅度。
圖5A顯示,CK1處理豌豆初生根MDA含量較之CK提高了97.40%,同時施加外源Ca2+后初生根MDA含量與CK1相比顯著降低,但仍高于CK組,且差異顯著。在T5處理下即Ca2+濃度為12.5 mmol/L時初生根MDA含量降低至與CK組無顯著差異,降低至CK1組的61.36%,為0.6732 μmol/(L·g),Ca2+濃度增加至15 mmol/L時,MDA含量又有所升高,為0.8086 μmol/(L·g)。
從圖5B可以看出,各處理組相對膜透性的變化趨勢與MDA含量的變化趨勢相似,CK1的相對膜透性(31.94%)是CK (14.16%)的2.25倍,且與低濃度Ca2+處理T1無顯著差異。之后,隨著外源Ca2+濃度的增加相對膜透性逐漸降低,但仍高于CK,T5與T6組則差異不顯著。
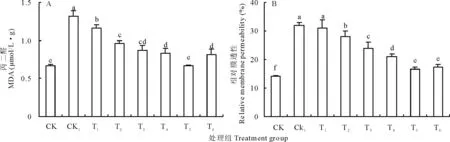
圖5 不同處理對豌豆初生根MDA(A)和相對膜透性(B)的影響Fig.5 Effects of different treatments on MDA (A) and relative membrane permeability (B) of pea primary roots
2.5 不同處理對豌豆初生根抗氧化物酶活性的影響
本試驗研究發現,與CK相比,CK1使豌豆初生根內的POD (圖6A)、SOD(圖6B)和CAT(圖6C)的活性顯著降低,APX活性(圖6D)則與CK無顯著差異。H2O2脅迫下,POD、SOD和CAT的活性均隨著外施Ca2+濃度的增大顯著增加,且在T4達到最大值后隨之降低,APX活性則在T4、T5和T6處理無顯著差異。
如圖6所示,CK1的POD和CAT活性顯著低于CK。CK1豌豆初生根的POD活性降低為CK組的55.19%(圖6A),CAT的活性降低為CK的37.29%,降低幅度比POD更大(圖6C)。同時施加Ca2+處理后,POD和CAT的活性顯著提高,在T4處理下均達最大值,分別為51.946 U/(mg·min)和1.9739 mmol/(L·g·min),是對應CK1的3.22和2.99倍。T4處理下POD活性相對于CK組的增加幅度比CAT更大,T4組POD活性比CK增加了78.28%,CAT活性則增加了13.87%,T5和T6處理下POD和CAT活性均有明顯降低,但都高于CK1(圖6A,C)。
SOD是一種重要的抗氧化酶類,能保護植物細胞受到氧化傷害。如圖6B所示,CK1組SOD活性比CK組降低了9.828%,隨著施加外源Ca2+濃度的增加,SOD活性隨之顯著增加,在T4處理下活性達到865.174 U/g FW,比CK和CK1分別高出101.06和172.026 U/g,均差異顯著。T5、T6處理下,酶活性降低,但是仍高于CK1,與CK組相比差異不大。
APX是清除細胞內H2O2的主要酶類。如圖6D所示,在CK1處理下APX活性并沒有像其他抗氧化酶類一樣降低,卻與CK無顯著差異。但施加外源Ca2+后酶活性又有顯著的提高,當Ca2+濃度增加到10 mmol/L (T4)達到穩定,與T5、T6無顯著差異。
3 討論
3.1 外源Ca2+對H2O2脅迫下豌豆初生根生長的緩解效應
大量研究表明,施加低濃度外源H2O2對植物的生長發育有著積極的作用,例如能夠促進種子萌發、增強很多植物的抗干旱、抗低溫、耐鹽等能力[29-31]。本試驗中發現,對豌豆外施高濃度(80 mmol/L)H2O2能抑制初生根正常向地性生長并發生水平彎曲現象(圖1,CK1)。眾所周知,向地性生長與細胞內的H2O2信號有關,也許是外源的H2O2擾亂了細胞內的氧化還原平衡,加劇了膜脂過氧化作用,造成細胞膜系統的傷害,從而導致了一系列的反應,使對重力的刺激信號無法向下傳遞。試驗結果表明,外源施加Ca2+能明顯的緩解H2O2對豌豆初生根的傷害,使其恢復正常生長。
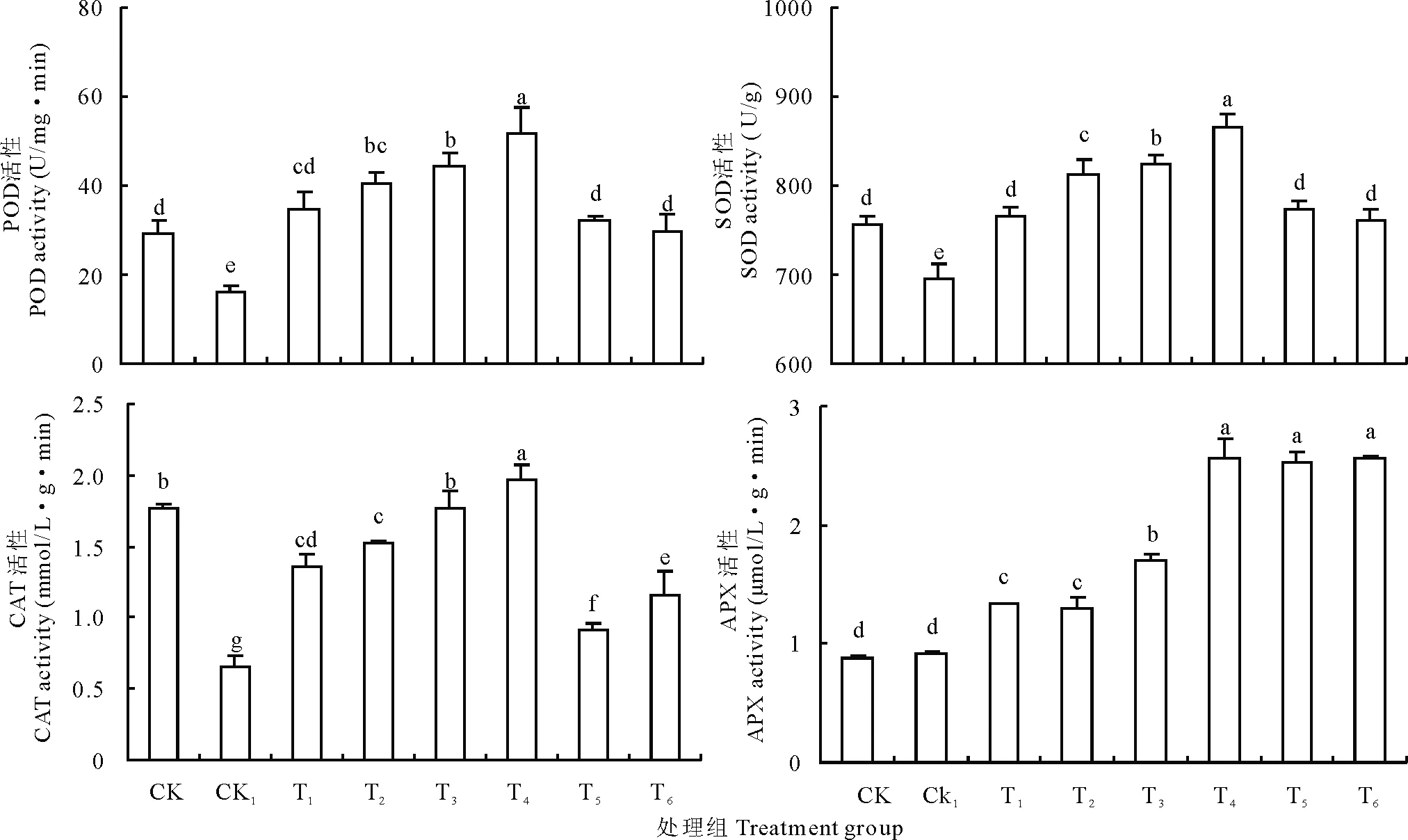
圖6 不同處理對豌豆初生根POD(A),SOD(B),CAT(C)和APX(D)活性的影響Fig.6 Effects of different treatments on POD(A), SOD (B), CAT(C) and APX(D) activity of pea primary roots
根系活力是根生長或者代謝旺盛程度的一個重要指標,根系活力的測定發現,初生根的活力也明顯受到了外施H2O2的抑制(圖4),這表明根細胞的正常代謝也受到了外源的H2O2抑制。施加外源Ca2+能夠有效地緩解H2O2誘導的水平彎曲生長,使根系生長趨于正常,根系活力得到恢復(圖4)。
一般認為,外源H2O2是一種特殊的化學物質,對植物的根產生脅迫。當植物受到逆境脅迫后,內源的H2O2水平會明顯的升高從而產生信號。但本研究結果與這一結論恰恰相反,在檢測了細胞內的H2O2水平后發現,H2O2脅迫下初生根內源H2O2(CK1)水平明顯低于正常處理下(CK)初生根內源H2O2水平(圖3)。因此認為,面對持續的外源H2O2處理,豌豆的種子或胚發生了自發的調節和反饋的抑制,抑制了自身的生長和代謝。植物根系通過呼吸作用產生的H2O2是其主要來源之一,本試驗中H2O2脅迫使初生根代謝降低,呼吸作用減弱,產生的H2O2也隨之減少。施加一定濃度的外源Ca2+后根系活力及代謝得以恢復,內源H2O2含量也隨之升高。
3.2 外源Ca2+對H2O2脅迫下豌豆初生根膜脂過氧化和抗氧化酶系統的緩解
有研究表明高鹽脅迫使植物的生長受到嚴重的阻礙,生物膜受到傷害[32]。王賀正等[33]指出水稻結實期隨水分脅迫加劇,細胞膜脂過氧化加劇。目前有關鋁誘導大豆的氧化脅迫也有報道,結果顯示高鋁脅迫使大豆根系質膜透性增大,使植株抵抗逆境的能力下降受到傷害[34]。當豌豆初生根受到脅迫時,首先受到傷害的是細胞膜,這很大程度上是通過破壞細胞生物膜的生理功能引起的,主要表現在細胞膜結構的破壞和功能的紊亂以及膜透性發生變化,而細胞膜的破壞將加速整個細胞的傷害。而當植物體受到逆境脅迫時,體內的MDA含量會升高,并且氧化程度越高,含量越高。
本研究中豌豆初生根的相對膜透性及MDA含量在不同的處理下均有不同程度的變化。H2O2處理導致初生根MDA含量的積累,相對膜透性增加。施加外源Ca2+后結果顯示,外施Ca2+有效減緩了初生根中MDA的升高,同時降低了細胞膜的相對透性,對細胞膜具有較好的修復和保護作用,從而減輕了外源H2O2對豌豆初生根細胞膜系統的傷害。
在遭受長時間各種逆境脅迫后植物的抗氧化系統包括SOD、 CAT、APX、ASC、GSH、GPX、POD等一些抗氧化的酶或者抗氧化劑均會降低或者減少。本試驗檢測了 CAT、SOD、POD和APX的活性,與大多數的研究結果一致,發現在外源H2O2處理下4種酶活性均明顯的降低(圖6)。柳斌等[35]曾報道施加外源鈣能緩解NaCl對苜蓿(Medicagosativa)幼苗根系的傷害。10 mmol/L的Ca2+能有效防護鹽脅迫導致的氧化損傷,抑制脂質過氧化作用[36]。同時,外源Ca2+能通過維持高的總抗氧化能力和抗氧化物質含量等途徑來增強黑藻對Cd脅迫的抗性[37]。本研究結果表明,外源Ca2+對CAT、SOD、POD和APX等酶活性具有明顯的促進作用,外施Ca2+后4種抗氧化酶活性持續升高。這個結果解釋了為什么內源H2O2水平會降低,同時也證明強大的清除系統緩解了逆境脅迫下產生的過量ROS造成的氧化脅迫,降低初生根的膜脂過氧化程度,保護初生根不被傷害。
4 結論
直接施加外源H2O2使豌豆初生根內MDA含量增加,相對膜透性增強,抗氧化物酶活性降低,對初生根造成膜脂過氧化傷害從而影響根系的正常生長。而同時施加10 mmol/L外源Ca2+能有效地緩解H2O2對豌豆初生根造成的氧化脅迫,減輕對細胞膜的傷害,降低質膜透性及MDA含量,增強初生根系抗氧化物酶活性,因此施加外源Ca2+能有效地緩解逆境脅迫對植物造成的氧化傷害。同時對農業生產中鹽漬及污染土壤的改良除了調整作物品種和改變耕作制度的方法外,外源施加一定濃度的Ca2+也是一種有效的技術手段。
