斑點叉尾源維氏氣單胞菌對四環(huán)素類抗生素的耐藥性及耐藥基因的檢測
2014-11-05 06:32:56汪開毓陳德芳黃凌遠王浩丞
水生生物學(xué)報 2014年2期
趙 敏 汪開毓, 王 均, 陳德芳 黃凌遠, 王浩丞
(1. 四川農(nóng)業(yè)大學(xué)魚病研究中心, 雅安 625014; 2. 四川農(nóng)業(yè)大學(xué)動物疫病與人類健康四川省重點實驗室, 雅安 625014)
維氏氣單胞菌(Aeromonas veronii)為氣單胞菌屬(Aeromonas)的一種, 亦被稱為維羅納氣單胞菌、凡隆氣單胞菌和維隆氣單胞菌, 存在于水體和淤泥等環(huán)境中[1], 能夠感染斑點叉尾(Ictalurus punctatus)[2,3]、錦鯉(Cyprinus carpio L.)[4]、西伯利亞鱘(Acipenser baerii)[5]、鯡形白鮭(Coregonus clupeaformis)[6]、華鯪(Sinilabeo rendahl)[7]、框鏡鯉(Cyprinus carpio)[8]等多種魚類。其中, 斑點叉尾感染維氏氣單胞菌近幾年才被發(fā)現(xiàn), 其發(fā)病率可達 30%以上, 病魚死亡率在 50%以上, 給養(yǎng)殖戶造成了巨大的經(jīng)濟損失, 也嚴(yán)重制約了斑點叉尾養(yǎng)殖業(yè)的健康發(fā)展[2,9]。
目前, 在魚病的防治過程中, 養(yǎng)殖業(yè)者為減少損失,首先采用的對策就是使用抗菌藥物, 但由于水產(chǎn)病原菌日趨嚴(yán)重的耐藥性, 使得抗菌藥難以達到預(yù)期的治療效果[10—12], 從而又給魚病的防治帶來了巨大的困難。其中,四環(huán)素類藥物為一種廣譜抗生素, 通過干擾細(xì)菌蛋白質(zhì)的合成起到抑菌作用[13], 可用于防治魚類的多種細(xì)菌性疾病, 而因臨床上對該類抗生素的不合理使用, 使得魚類病原菌對此類抗生素的耐藥性變的尤為突出[10—12]。
2006—2011年, 我實驗室先后從四川地區(qū)人工養(yǎng)殖的斑點叉尾病樣中, 分離得到維氏氣單胞菌共 42株。本試驗以這些菌株作為研究對象, 旨在分析四川地區(qū)斑點叉尾源維氏氣單胞菌對四環(huán)素類抗生素的耐藥狀況以及相關(guān)耐藥基因, 為臨床合理用藥以及進一步研究維氏氣單胞菌的耐藥機制提供部分參考依據(jù)。
1 材料與方法
1.1 菌株
1.2 主要試劑
藥敏紙片(四環(huán)素 30 μg/片, 強力霉素 30 μg/片)和MH培養(yǎng)基購自杭州微生物試劑有限公司, 細(xì)菌基因組DNA提取試劑盒(DP302-2)、2×Taq Master Mix和感受態(tài)細(xì)胞 DH5α購自天根生物科技有限公司, 克隆載體pMD19-T、DNA Marker及 DNA純化回收試劑盒購自TaKaRa公司, 氨芐青霉素鈉購自上海生工生物技術(shù)有限公司, 瓊脂糖購自廣州英韋創(chuàng)津生物科技有限公司, 鹽酸四環(huán)素(98.5%)和鹽酸多西環(huán)素(98%)由四川省民生藥業(yè)有限責(zé)任公司惠贈。
1.3 菌株培養(yǎng)及藥物敏感性試驗
在無菌條件下, 取低溫保存的維氏氣單胞菌和質(zhì)控菌株大腸桿菌ATCC25922接種于MH肉湯中, 28℃恒溫?fù)u床120 r/min振蕩培養(yǎng)過夜后, 采用麥?zhǔn)媳葷岱ㄕ{(diào)整菌液濁度為0.5麥?zhǔn)媳葷峁軡岫取H缓? 參照CLSI[13]推薦的紙片擴散法和微量肉湯稀釋法(藥物濃度由 128 μg/mL 2倍稀釋至0.25 μg/mL)對所有菌株進行藥物敏感性分析; 結(jié)果參照2008年版 CLSI抗微生物藥物敏感性試驗執(zhí)行標(biāo)準(zhǔn)進行判定。
1.4 tet基因片段獲得與分析
按照天根細(xì)菌基因組 DNA提取試劑盒(DP302-2)的操作說明提取菌體基因組DNA, –20 ℃保存?zhèn)溆谩K沫h(huán)素類耐藥基因的引物序列參照Nawaz, et al.[14]的報道, 詳見表 1。引物由上海生工生物技術(shù)有限公司合成。 以維氏氣單胞菌基因組DNA為模板, 分別用相應(yīng)的引物擴增目標(biāo)基因tet A、tet B、tet C、tet D和tet E; PCR反應(yīng)體系為 50 μL: 2×Taq Master Mix 25 μL, DNA 模板 2.0 μL,上、下游引物各 1.0 μL, 無菌超純水 21 μL。反應(yīng)條件:94℃預(yù)變性2min, 94℃ 20s, 53℃10s, 65℃ 45s共30個循環(huán), 最后 65℃延伸 4min。PCR擴增后的產(chǎn)物在 1%瓊脂糖凝膠中進行電泳。采用DNA純化回收試劑盒回收各個基因的陽性擴增產(chǎn)物(每個基因隨機選取其中的 5個, 不足 5個則全部回收), 相關(guān)操作參照試劑盒說明進行。構(gòu)建pMD19-T-tet重組質(zhì)粒, 然后提取質(zhì)粒, 采用對應(yīng)引物進行PCR鑒定, 將鑒定為陽性的質(zhì)粒送往公司進行測序。測序結(jié)果使用DNAMAN軟件進行序列間相似性分析, 再利用 NCBI中的 Blastn系統(tǒng)進行序列的相似性搜索, 并使用Clustal X 2.0和Mega 4.0軟件對tet基因構(gòu)建系統(tǒng)發(fā)育樹。
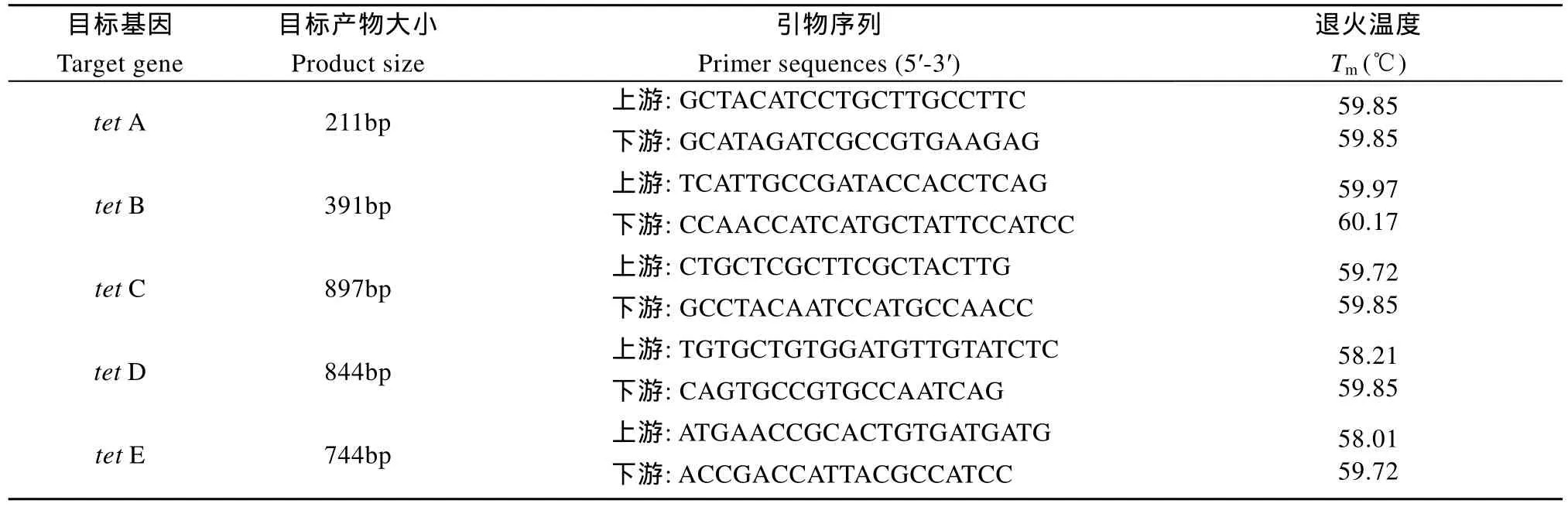
表1 四環(huán)素耐藥基因擴增引物Tab. 1 Sequences of the primers used in the study
2 結(jié)果
2.1 藥物敏感性測試結(jié)果
在本試驗中同時采用了紙片擴散法和微量肉湯稀釋法來測定維氏氣單胞菌的藥物敏感性, 參照吳俊偉[16]所描述的方法, 以紙片擴散法中抑菌圈的大小作為評價維氏氣單胞菌藥物敏感性的主要依據(jù), 在進行耐藥水平最終結(jié)果判定的時候, 對抑菌圈顯示為中介, 而 MIC值較大的菌株判定為耐藥, 對抑菌圈遠大于標(biāo)準(zhǔn)而 MIC值偏大的菌株判定為敏感。最終得出42株維氏氣單胞菌對四環(huán)素類的藥物敏感性結(jié)果(表2、表3)。
2.2 PCR擴增結(jié)果
2.3 tet基因的克隆與分析
經(jīng)測序, tetA、tetB、tetC和tetE基因片段大小分別為 211、391、891和 744 bp, 與預(yù)期結(jié)果相符。使用DNAMAN軟件分別對5個tetA、2個tetB、4個tetC和2個 tetE基因片段進行多序列比對分析, 結(jié)果顯示其一致性均為100%。然后將上述4類基因片段序列在NCBI中的 Blastn系統(tǒng)進行序列同源性分析, 結(jié)果顯示: tetA、tetB、tetC和tetE基因片段分別與GenBank中登陸的多個tet基因的同源性為100%。

圖1 tetA基因系統(tǒng)發(fā)育樹Fig. 1 Phylogenetic tree of tetA gene
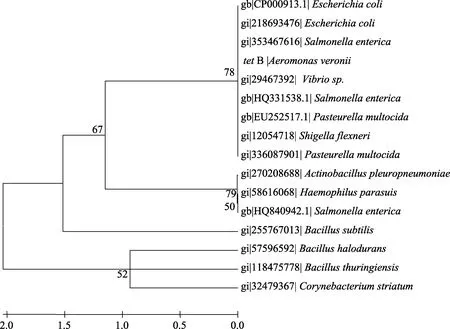
圖2 tetB基因系統(tǒng)發(fā)育樹Fig. 2 Phylogenetic tree of tetB gene
在GenBank中分別下載多個tet A、tet B、tet C和tet E基因用于構(gòu)建系統(tǒng)發(fā)育樹(圖1-4)。結(jié)果顯示tetA基因與存在于點狀氣單胞菌(Aeromonas punctata)pFBAOT6質(zhì)粒(GenBank登錄號: 51492513)中的tetA基因, 大腸桿菌O69 (Escherichia coli serotype O69)質(zhì)粒介導(dǎo)的tetA基因(GeneBank登錄號: 227434002)、大腸桿菌Q19中的tetA基因(GeneBank登錄號: 393195318)、肺炎克雷伯菌(Klebsiella pneumoniae)pKPI-6質(zhì)粒(GenBank登錄號:394557614)中的tetA基因、蠟樣芽胞桿菌(Bacillus cereus)中的 tetA基因(GenBank登錄號: 242277425)聚為一簇;tetB基因與來自福氏志賀氏菌(Shigella flexneri)中質(zhì)粒介導(dǎo)的 tetB基因(GenBank登錄號: 12054718)、大腸桿菌pTC1質(zhì)粒中的tetB基因(GenBank登錄號: CP000913.1)、多殺巴斯的桿菌(Pasteurella multocida)中的 tetB基因(GenBank登錄號分別為: 336087901、EU252517.1)、沙門氏菌(Salmonella enterica)質(zhì)粒介導(dǎo)的 tetB基因(GenBank登錄號:353467616、HQ331538.1)等聚為一簇;tetC基因與大腸桿菌中的 tetC基因(GenBank登錄號分別為: 190887129、190887131、190887127、190887125)、沙門氏菌中的tetC基因(GenBank登錄號: GU987054.1)以及plPO2T質(zhì)粒中的 tetC基因(GenBank登錄號:21735067)聚為一簇; tetE基因則與來自殺鮭氣單胞菌(Aeromonas salmonicida)中的 tetE基因(GenBank登錄號分別為: 113129387、145301211)聚為一簇。
3 討論
在本研究中, 采用了紙片擴散法和微量肉湯稀釋法對 42株維氏氣單胞菌進行了藥物敏感性測試, 兩種方法所得出的結(jié)果并不完全一致, 為了能夠確定最終的結(jié)果, 因此參考吳俊偉[15]所提出的方法, 以紙片擴散法中抑菌圈的大小作為評價菌株耐藥性的主要依據(jù), 參考MIC值進行評判, 對抑菌圈中介, 而MIC值較大的菌株判定為耐藥, 對抑菌圈遠大于標(biāo)準(zhǔn)而MIC值偏大的菌株判定為敏感。最終得出受試菌株對四環(huán)素耐藥的共有28株, 耐藥率為66.67%; 對強力霉素耐藥的共有24株, 耐藥率為57.14%,明顯低于四環(huán)素。但從敏感菌株來看,對四環(huán)素和強力霉素敏感的菌株均為10株, 敏感率為23.80%, 兩者沒有差別, 這說明了四川地區(qū)的維氏氣單胞菌對于四環(huán)素和強力霉素的耐藥性已經(jīng)達到了非常嚴(yán)重的水平。
在革蘭氏陰性菌中, 細(xì)菌對四環(huán)素類藥物的耐藥機制主要有兩種, 一種是核糖體保護蛋白, 該蛋白能保護核糖體免受四環(huán)素類藥物的作用從而產(chǎn)生耐藥[16]; 另一種是主動外排作用, 通過外排蛋白將四環(huán)素排出胞外,細(xì)胞內(nèi)的藥物濃度因此降低, 核糖體因此受到保護, 從而產(chǎn)生了耐藥性, 這是最早被研究的一種耐藥機制[17],也是本研究中被證明存在于維氏氣單胞菌中的一種耐藥機制。外排蛋白屬于主要異化超家族(Major facilitatorsuperfamily, MFS)中的一員[18]。目前, 已有超過 20種不同的四環(huán)素外排蛋白被發(fā)現(xiàn), 這些蛋白被分為 6個不同的類群[17], tetA、tetB、tetC、tetD和tetE基因所編碼的外排蛋白屬于該6個類群中的第1類群[19], 均發(fā)現(xiàn)于革蘭氏陰性菌中[17,19]; 該類群蛋白具有12個跨膜α-螺旋, 其四環(huán)素抗性蛋白具有 41%—78%的氨基酸同源性, 而它們的四環(huán)素阻抑蛋白具有 37%—88%的氨基酸同源性[20]。在氣單胞菌中, tetA、tetB、tetC、tetD和tetE基因已經(jīng)先后被證實存在于該屬中, 并且發(fā)現(xiàn)tetE基因在該5種耐藥基因中檢出率最高[21—24], 說明該基因是介導(dǎo)氣單胞菌分離株對四環(huán)素類藥物耐藥的優(yōu)勢基因。Nawaz, et al.[14]對分離自鯰魚的 81株耐四環(huán)素的維氏氣單胞菌進行了相關(guān)研究, 發(fā)現(xiàn)tetA、tetB、tetC、tetD和 tetE檢出率分別為 3.70%(3/81)、28.40%(23/81)、2.47%(2/81)、2.47%(2/81)、90.12%(73/81), 其中 tetE基因的檢出率顯著高于其他 4個基因, 為其優(yōu)勢基因, 符合上述觀點。然而,Jacobs, et al.[25]在對37株分離自南非水產(chǎn)養(yǎng)殖環(huán)境中的氣單胞菌的研究中, 以及 Kim, et al.[26]對8株分離自韓國的耐四環(huán)素殺鮭氣單胞菌的研究中都發(fā)現(xiàn)tetA基因在這些菌株中檢出率最高。在本研究中, 同樣發(fā)現(xiàn)四環(huán)素耐藥的菌株 tetA基因檢出率最高, 且明顯高于其他 4個基因, 而 tetE基因僅在 2株菌中被檢測到, 與最初的研究結(jié)果存在差異; 這說明在維氏氣單胞菌中, 四環(huán)素類耐藥基因的流行存在地域性差異。
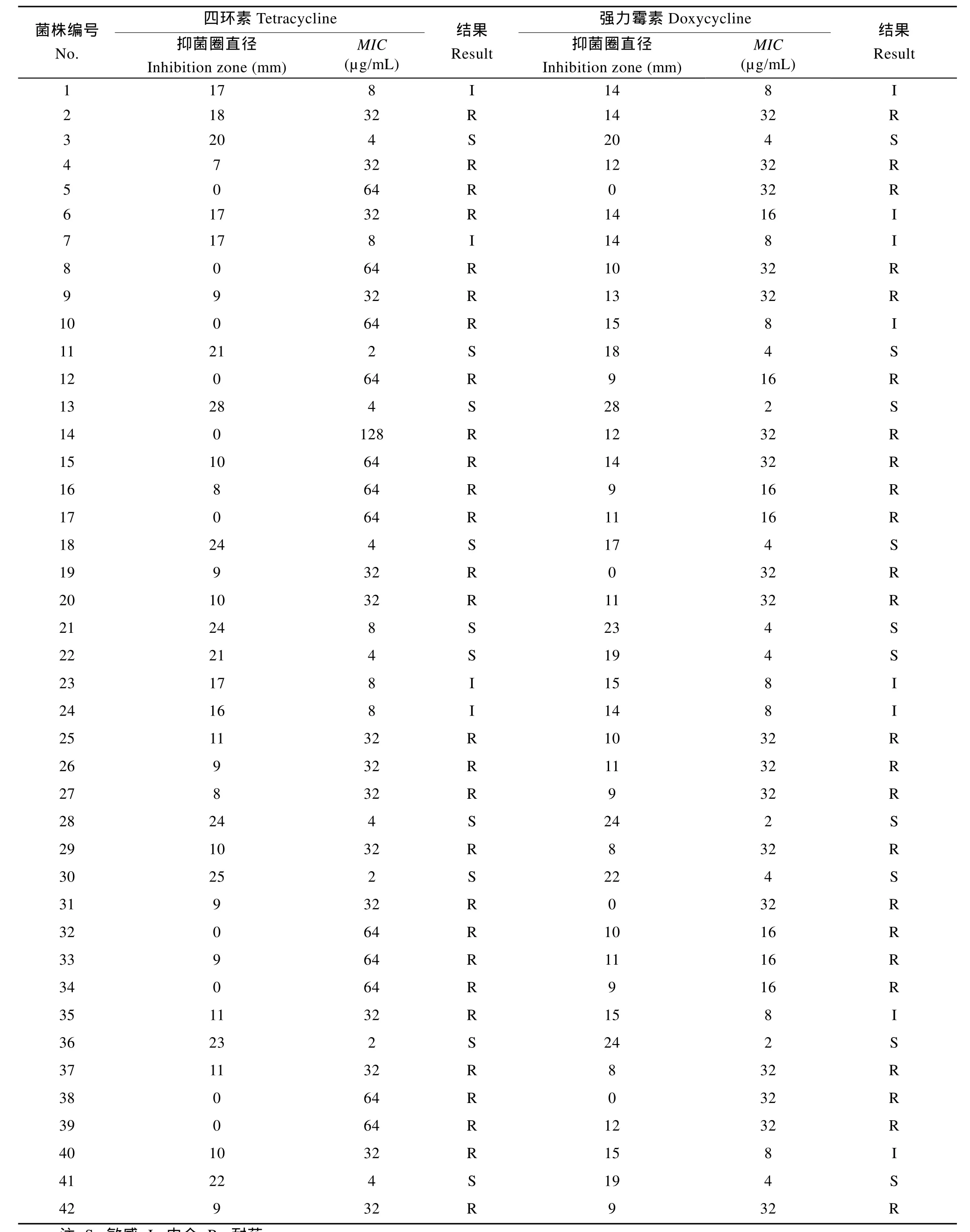
表2 42株維氏氣單胞菌對四環(huán)素類抗生素的敏感性試驗結(jié)果Tab. 2 Susceptibility to tetracycline antibiotics of 42 Aeromonas veronii strains
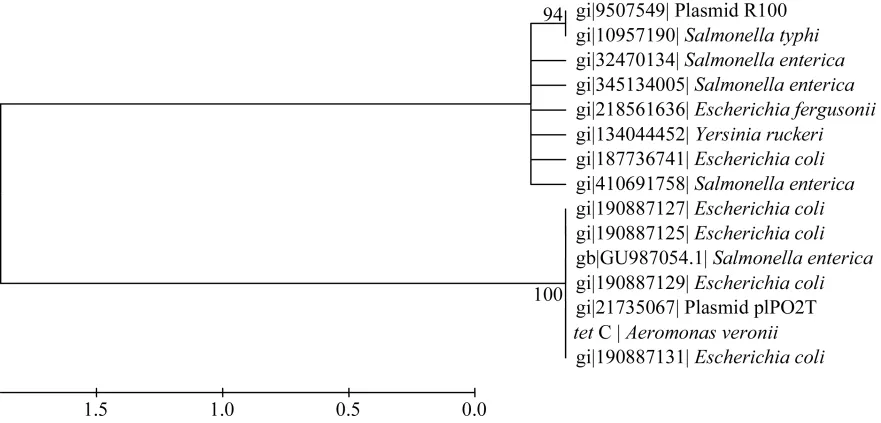
圖3 tetC基因系統(tǒng)發(fā)育樹Fig. 3 Phylogenetic tree of tetC gene
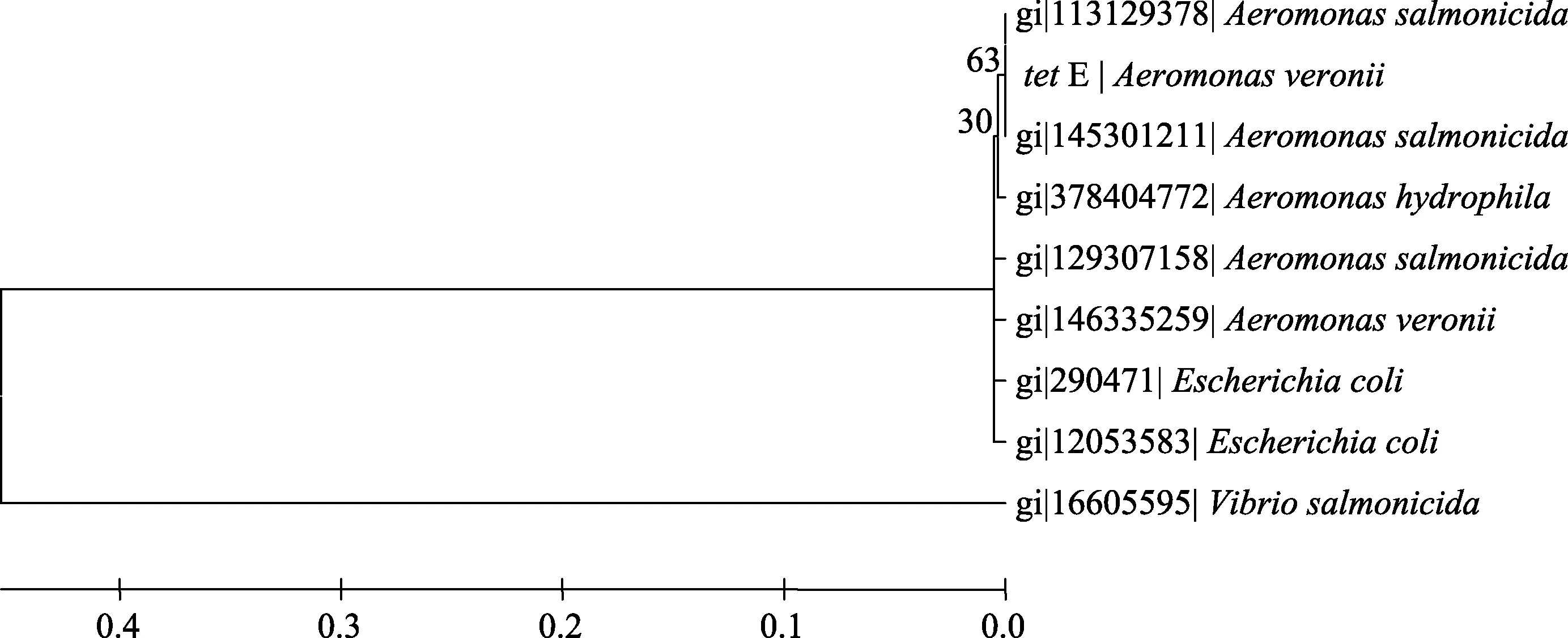
圖4 tetE基因系統(tǒng)發(fā)育樹Fig. 4 Phylogenetic tree of tetE gene
同時, 本研究還發(fā)現(xiàn)在 16和 29號菌株中同時檢測到tetA和tetC, 在8號菌株中同時檢測到tetA、tetB和tetC; 關(guān)于一株菌中同時檢測出幾種四環(huán)素類耐藥基因的情況, 在Nawaz, et al.[15], Schmidt, et al.[23]以及 ?í?ek A, et al.[25]各自的研究中已有報道。此外, 我們的研究還發(fā)現(xiàn)在耐藥菌株4、10、12中并未檢測到上述5種耐藥基因, 說明在維氏氣單胞菌中可能還存在其他的四環(huán)素耐藥機制, 有待進一步研究。
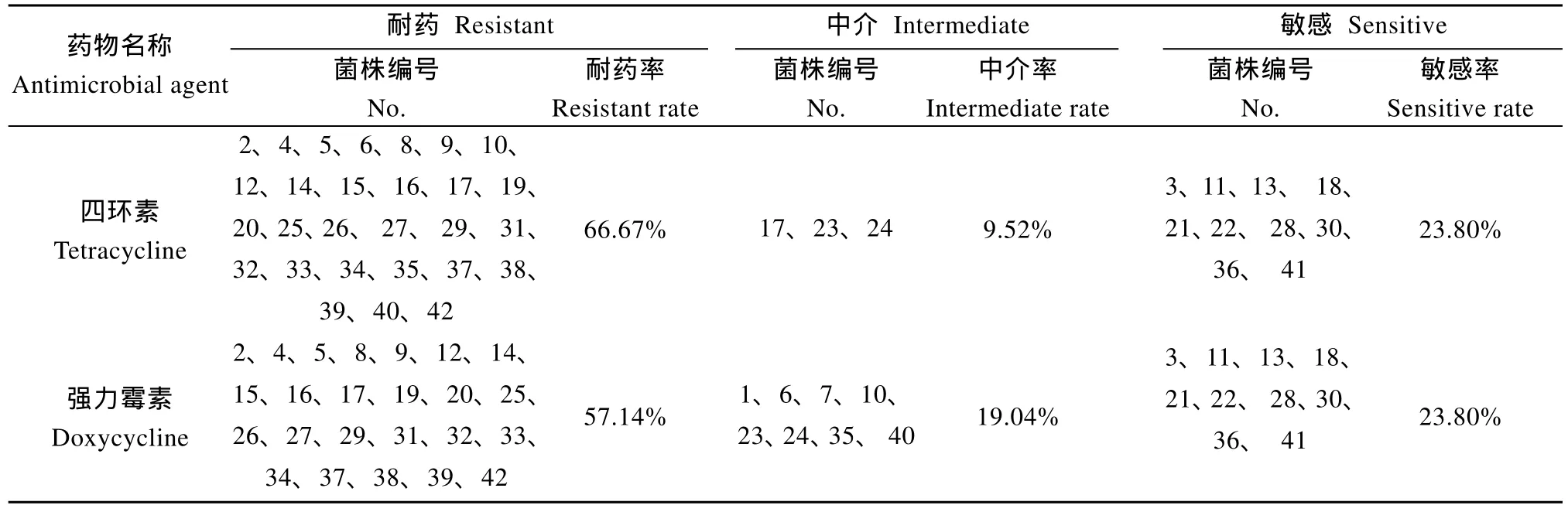
表3 42株維氏氣單胞菌對四環(huán)素類抗生素的敏感性試驗結(jié)果Tab. 3 Susceptibility to tetracycline antibiotics of 42 Aeromonas veronii strains
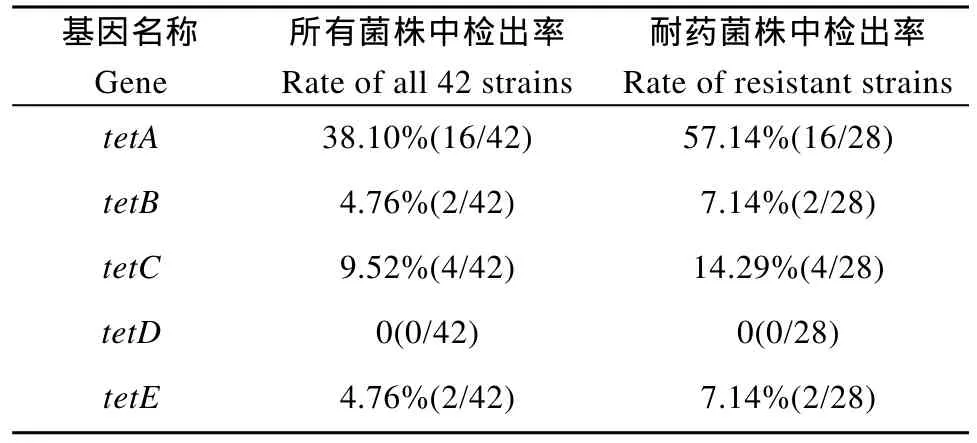
表4 各基因的檢出率Tab. 4 The positive rate of the gene
在本研究中, 首次在國內(nèi)維氏氣單胞菌分離株中檢測到tetA、tetB、tetC和tetE基因, 為進一步研究我國境內(nèi)的維氏氣單胞菌的耐藥機制提供了一定的理論依據(jù);同時, 本研究的結(jié)果也突出了目前我國水產(chǎn)病原菌耐藥性的嚴(yán)重性。因此, 如何防止水產(chǎn)病原菌耐藥性的產(chǎn)生和傳播, 以及在魚病的防治過程中如何應(yīng)對因病原菌耐藥性而造成的困難值得廣大學(xué)者共同關(guān)注。
猜你喜歡
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
現(xiàn)代臨床醫(yī)學(xué)(2022年3期)2022-06-06 07:59:40
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
昆明醫(yī)科大學(xué)學(xué)報(2022年1期)2022-02-28 07:43:40
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科學(xué)大眾(2020年12期)2020-08-13 03:22:22
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
