核糖體蛋白L22對果蠅發育的影響及機制
2014-08-15 03:25:20李紅艷
河北大學學報(自然科學版) 2014年1期
李紅艷
(西藏民族學院 醫學院,陜西 咸陽 712082)
核糖體蛋白在核糖體組裝和蛋白質合成中有重要作用.人類中一些核糖體蛋白的異常表達常引起一些疾病的發生,如Diamond-Blackfan貧血癥[1]、特納綜合征[2]、聽力喪失[3]和癌癥[4].研究發現,果蠅中一些核糖體蛋白的突變可引起幼蟲發育遲緩,剛毛短小、稀疏,生殖力和生活力低下,并隱性致死等特征,這些表型被稱為Minute表型.目前對其解釋為:某一核糖體蛋白基因突變后,引起細胞生長和增殖率減少[5-6];編碼核糖體蛋白的基因發生突變,導致核糖體合成受損,引起蛋白合成速率減少[7].另外,一些核糖體蛋白在細胞周期、凋亡、DNA損傷修復等過程中也有著重要作用,但這些作用和該核糖體蛋白在核糖體組裝和蛋白質合成中的作用無關.
L22是核糖體60 S大亞基的組成成分,除了自身參與核糖體組裝和蛋白質合成外,并與其他多亞基核糖核蛋白顆粒一起發揮相應的生物學功能[8].另外,L22通過與病毒調節蛋白如ICP-4和ICP-22相互作用[9],調節病毒早期基因EAP的轉錄[10].L22誘導p53表達,選擇性阻止αβT細胞發育[11].果蠅Kc細胞刪除L22后,通過與組蛋白H1相互作用抑制基因的轉錄[12].但有關L22在果蠅發育中的作用及機制還不清楚.本研究證實,L22對于果蠅正常發育是不可缺少的,且主要是通過細胞凋亡和細胞周期阻滯的方式發揮作用的.
1 材料和方法
1.1 L22 RNAi轉基因果蠅的獲取和飼養
以果蠅L22 cDNA編碼區為模板,進行PCR擴增(上游引物5-caagcggccgcggtgaacaacctgggcaa-3;下游引物5-ccggaattcgcatcgtcgtcctcatcgtc-3).將擴增產物通過Not I和EcoR酶切位點克隆到SympUAST-w載體中,稱為L22-SympUAST-w.通過顯微注射法將L22-SympUAST-w質粒注射到果蠅w1118胚胎中,經多步篩選最終獲得L22RNAi轉基因果蠅.為了驅動L22在果蠅中特異性干擾,實驗中用到的其他商品化果蠅品系有Hsp70-Gal4,act5c-Gal4,ey-Gal4和 vg-Gal4.果蠅飼養在18 ℃/25 ℃玉米粉-蔗糖培養基上.
1.2 果蠅幼蟲體長測量
L22RNAi雌蠅與Hsp70-Gal4雄蠅在25 ℃下雜交,間隔1 h產卵,1 h后將果蠅轉移到新的培養基下使其再次產卵,依次循環.產卵24 h后,將幼蟲在37 ℃熱激30 min.之后每隔24 h隨機收集1次幼蟲,每組收集至少10只,至對照組發育到蛹期.使用MZ16體視顯微鏡對幼蟲拍照.利用MetaMorph軟件統計幼蟲體長.
1.3 吖啶橙染色
在PBS緩沖液中解剖L22RNAi與vg-Gal4雜交后的三期幼蟲翅膀discs.解剖好后立即放到4 ℃果蠅細胞培養基中,使其維持正常的生理環境.之后轉移到載玻片上,并用100 μL的培養基洗1次.室溫下,以1 μg/mL 吖啶橙(sigma)染色5 min后,即刻在熒光鏡下收集數據.
1.4 果蠅S2細胞中干擾L22
PCR擴增果蠅L22編碼序列(上游引物[含有1個T7啟動子]:5-taatacgactcactatagggagaatggctcctaccgccaagacc-3;下游引物5-taatacgactcactatagggagattactcggcatcgtcgtcctc-3).PCR產物純化后作為模板,再通過MEGAscrip T7 (Ambion)試劑盒生成單鏈RNA(ssRNAs).ssRNAs在65 ℃中退火,孵育30 min,自然降溫至室溫即生成雙鏈RNA(dsRNAs).經乙醇沉淀,重懸在一定量的無RNA酶水中.再經凝膠電泳對其定量,-80 ℃保存備用.
S2細胞培養在Schneider 果蠅基本培養基(Gibco)中,補加終質量濃度為100 g/L的胎牛血清(Gibco),160 μg/mL 青霉素,250 μg/mL 鏈霉素和4 mmol/LL-谷氨酰胺(Sigma),25 ℃培養.依照之前的方法[12]在S2細胞中對L22 進行干擾.用無血清培養基將S2 細胞以1×106個/mL密度種植在35 cm 培養瓶中(Corning).室溫,1 mL無血清培養基加終質量濃度為5 μg/mL L22 dsRNA撫育60 min.隨后補加含有血清的正常培養基2 mL繼續培養.同時以GFP dsRNA作為對照組,處理對照組細胞.在處理后的第4和第8天收集細胞,用作實驗.
1.5 RT-PCR
根據試劑盒(Qiagen)要求提取收集到的細胞總RNA.并對提取的RNA進行DNase(TaKaRa)處理,以防止基因組DNA污染.以提取的RNA為模板,用M-MLV 反轉錄酶(Invitrogen)和oligo-dT 引物合成cDNA.PCR擴增條件(25 μL 反應體系):95 ℃預變性5 min;94 ℃, 30 s,60 ℃,30 s,72 ℃,45 s,30 個循環;72 ℃延伸8 min.同時實驗中為了進一步排除基因組污染的干擾,一并檢測了每一組RNA 樣品在沒有加反轉錄酶的情況下參照基因的表達.用到的引物見表1.

表1 RT-PCR引物
1.6 統計分析
所有實驗重復在3次以上.數據使用Excel 軟件進行統計分析.組間樣本均數比較采用t檢驗,以P<0.05為有統計學意義.
2 結果
2.1 減少L22表達延遲果蠅幼蟲發育
實驗首先用act5c-Gal4驅動L22表達廣泛減少,以初步了解L22在果蠅發育中的作用.結果果蠅死在胚胎期,而對照組發育正常.同時用Hsp70-Gal4驅動子誘導性驅動L22沉默(方法見材料部分).L22RNAi/Hsp70-Gal4從一期到三期幼蟲均發育延遲,持續10~12 d,最后死在三期幼蟲期;對照組發育正常,在幼蟲期持續6~7 d,之后發育成蛹和成蟲.在早48 h,L22RNAi/Hsp70-Gal4生長與對照組比沒有明顯變化,而72 h后,較對照組體長明顯變小(圖1 a,b).由此說明L22是果蠅早期發育所必須的.
2.2 減少L22表達引起果蠅眼和翅膀生長缺陷
實驗進一步用特異性驅動L22在眼睛和翅膀中表達沉默的ey-Gal4和vg-Gal4驅動子.果蠅復眼由一定數目的單眼和特異性視網膜細胞精確排列組成.復眼已成為一個很好的研究細胞增殖和細胞存活的系統[13].統計了500只L22RNAi/ey-Gal4果蠅,其表型可分為3類:第1類(占17%)與正常對照組比沒有顯著差異.第2類(占6%)眼表型與對照組相比較小,可以從蛹中正常孵育出來(圖2a).而第3類(占77%)死在蛹期,解剖發現這些果蠅身體其他部位發育完整,只是缺少頭部結構.所有L22RNAi/vg-Gal4果蠅的翅膀變小而且多褶皺(圖2c).這些異常表型暗示:L22干擾后引起眼和翅膀discs發育不完全,可能是由細胞增殖阻滯或細胞死亡引起的.
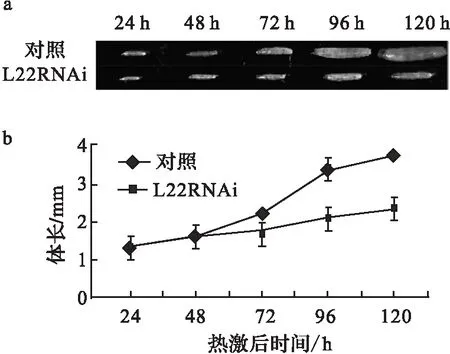
a.熱激后L22RNAi/Hsp70-Gal4 幼蟲和對照組幼蟲長度比較;b.前48 h,L22RNAi/Hsp70-Gal4幼蟲和對照組體長相差不顯著; 72 h 后,L22RNAi/Hsp70-Gal4 幼蟲生長顯著慢于對照組(P<0.05).圖1 減少L22表達阻滯果蠅幼蟲發育Fig.1 Decreased expression of L22 caused the Drosophila larvae growth arrest
2.3 減少L22表達降低S2細胞數目和果蠅翅膀disc中出現細胞凋亡
已知細胞數目反應了細胞增長和細胞凋亡之間的關系[14].所以在果蠅S2細胞中干擾L22表達,以GFP作為對照,統計細胞數.結果發現,在L22干擾后S2細胞數目減少(圖3a).且這種變化隨著時間的延長越來越明顯.這一結果與異常的眼和翅膀表型吻合.表明L22RNAi后影響了果蠅的發育,且可能是通過細胞凋亡的方式完成的.同時吖啶橙(細胞凋亡熒光染料)染色實驗證實,在L22RNAi/vg-Gal4三期幼蟲的翅膀discs中確實有明顯的細胞凋亡信號出現(圖2d).

a.與野生型相比,6%L22RNAi/ey-Gal4 眼睛變小,77%無頭;b.在統計的500只果蠅中L22RNAi/ey-Gal4各種表型所占的比例;c.L22RNAi/vg-Gal4后代翅膀缺陷;d.吖啶橙染色顯示,L22RNAi/vg-Gal4 三期幼蟲的翅膀 disc 細胞出現凋亡現象.圖2 減少L22表達造成果蠅眼睛和翅膀缺陷Fig.2 Eye and wing defects in L22-depleted flies
2.4 L22刪除后細胞凋亡/細胞周期相關基因表達異常
以上結果表明,L22干擾后引起了細胞凋亡,而凋亡與細胞周期緊密相關.因此通過RT-PCR檢測了與細胞凋亡相關基因(hid,reaper,dcp-1和p53)和細胞周期相關基因(MCM5,cdc45,incenp和cyclinB)在S2細胞中L22干擾后的變化情況.結果發現,刪除L22,引起hid,reaper,dcp-1和p53基因表達水平升高(圖3b),而這種變化與他們各自在細胞凋亡中的作用是一致的.已知p53是一腫瘤抑制蛋白,在各種應激條件下均可以開啟凋亡的發生[2],通常是通過刺激細胞色素C釋放來激活caspase活性引起的[15].Dcp-1是果蠅caspase家族的一個成員,也在細胞凋亡過程中有著重要作用[16].Reaper和hid是果蠅中另外2個與細胞凋亡相關的基因[17-18],它們可以直接或間接通過激活p53來參與凋亡[19],另外發現它們在發揮細胞凋亡過程中也激活了caspase通路[2-21].
另外,L22RNAi后G1-S期的MCM5與cdc45 和G2-M期的incenp與cyclinB表達水平分別減少(圖3c).MCM5是p53的一個轉錄抑制target,在G1-S期中有著重要作用[22].細胞分裂周期蛋白45 (cdc45)對于細胞周期起始以及DNA復制延伸都有著重要作用[23].研究發現,Cdc45與MCM5/cdc46,MCM7/cdc47和ORC[24]相互作用.Cyclin B 有控制細胞進入有絲分裂期的作用,減少cyclin B表達導致G2期阻滯,而且該過程受到p53依賴的轉錄抑制調控[25].Incenp在細胞分裂過程中有重要作用.研究發現Incenp與Aurora B 和survivin 形成Aurora B/incenp/survivin復合體,在染色體正常分離過程中發揮作用[26-28].在黑色素細胞中,p53可以通過直接與survivin啟動子結合或者激活p21來抑制survivin表達[29].因此,Incenp與p53之間可能有一種負相關.
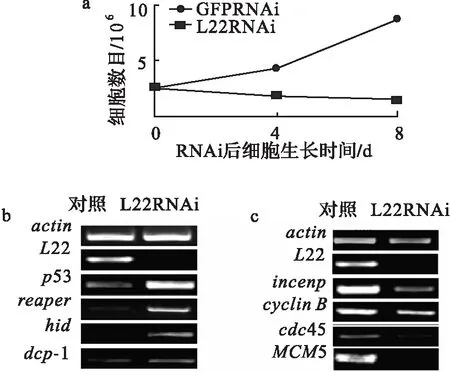
a.L22 RNAi后S2細胞生長受到抑制,而GFPRNAi對照組細胞生長正常;b.L22RNAi導致p53,hid,dcp-1和reaper的轉錄水平上升; c.actin轉錄正常,而MCM5,cdc45,cyclin B和incenp轉錄水平均下降.圖3 S2細胞中減少L22表達引起細胞凋亡和細胞周期相關基因表達異常Fig.3 Abnormal expression of the genes related to apoptosis and cell cycle in L22-depleted S2 cells
綜上所述,L22對于果蠅正常發育是不可缺少的,L22RNAi后主要是通過細胞凋亡的方式來影響果蠅發育的.
致謝:感謝清華大學孫方霖教授給予的實驗指導和經費支持;感謝清華大學陳蘇同學在實驗過程中所給的幫助.
參 考 文 獻:
[1] DRAPTCHINSKAIA N, GUSTAVSSON P, ANDERSSON B, et al.The gene encoding ribosomal protein S19 is mutated in Diamond-Blackfan anaemia[J].Nat Genet, 1999, 21: 169-175.
[2] FISHER E M, BEER-ROMERO P, BROWN L G, et al.Homologous ribosomal protein genes on the human X and Y chromosomes: escape from X inactivation and possible implications forTurnersyndrome[J].Cell, 1990, 63: 1205-1218.
[3] O’BRIEN T W, O’BRIEN B J, NORMAN R A.Nuclear MRP genes and mitochondrial disease[J].Gene, 2005, 354: 147-151.
[4] RUGGERO D, PANDOLFI P P.Does the ribosome translate cancer[J].Nat Rev Cancer,2003, 3: 179-192.
[5] SCHULTZ J.The minute reaction in the development ofDrosophilamelanogaster[J].Genetics, 1929, 14: 366-419.
[6] LAMBERTSSON A.The minute genes inDrosophilaand their molecular functions[J].Adv Genet, 1998, 38: 69-134.
[7] ENERLY E, LARSSON J, LAMBERTSSON A.Silencing theDrosophilaribosomal protein L14 gene using targeted RNA interference causes distinct somatic anomalies[J].Gene, 2003, 320: 41-48.
[8] WOOD J, FREDERICKSON R M, FIELDS S, et al.Hepatitis C virus 3’X region interacts with human ribosomal proteins[J].J Virol, 2001, 75: 1348-1358.
[9] NG T I, CHANG Y E, ROIZMAN B.Infected cell protein 22 of herpes simplex virus 1 regulates the expression of virion host shutoff gene U(L)41[J].Virology, 1997, 234: 226-234.
[10] DOBBELSTEIN M, SHENK T.In vitro selection of RNA ligands for the ribosomal L22 protein associated with Epstein-Barr virus-expressed RNA by using randomized and cDNA-derived RNA libraries[J].J Virol, 1995, 69: 8027-8034.
[11] ANDERSON S J, LAURITSEN J P, HARTMAN M G, et al.Ablation of ribosomal protein L22 selectively impairs alphabeta T cell development by activation of a p53-dependent checkpoint[J].Immunity, 2007, 26: 759-772.
[12] NI J Q, LIU L P, HESS D, et al.Drosophilaribosomal proteins are associated with linker histone H1 and suppress gene transcription[J].Genes Dev, 2006, 20: 1959-1973.
[13] SHI W, STAMPAS A, ZAPATA C, et al.The pineapple eye gene is required for survival ofDrosophilaimaginal disc cells[J].Genetics, 2003, 165: 1869-1879.
[14] KASTAN M B, CANMAN C E, LEONARD C J.Leonard.P53, cell cycle control and apoptosis: implications for cancer[J].Cancer Metastasis Rev, 1995, 14: 3-15.
[15] SCHULER M, BOSSY-WETZEL J C, GOLDSTEIN J C, et al.P53 induces apoptosis by caspase activation through mitochondrial cytochrome c release[J].J Biol Chem, 2000, 275: 7337-7342.
[16] SONG Z, MCCALL K, STELLER H.DCP-1, aDrosophilacell death protease essential for development[J].Science, 1997, 275: 536-540.
[17] GRETHER M E, ABRAMS J M, AGAPITE J, et al.The head involution defective gene ofDrosophilamelanogasterfunctions in programmed cell death[J].Genes Dev, 1995, 9: 1694-1708.
[18] WHITE K, GRETHER M E, ABRAMS J M, et al.Genetic control of programmed cell death inDrosophila[J].Science, 1994, 264: 677-683.
[19] BRODSKY M H, WEINERT B T, TSANG G, et al.DrosophilamelanogasterMNK/Chk2 and p53 regulate multiple DNA repair and apoptotic pathways following DNA damage[J].Mol Cell Biol, 2004, 24: 1219-1231.
[20] WHITE K, TAHAOGLU E, STELLER H.Cell killing by theDrosophilagene reaper[J].Science, 1996, 271: 805-807.
[21] ZHOU L, SCHNITZLER A, AGAPITE J, et al.Cooperative functions of the reaper and head involution defective genes in the programmed cell death ofDrosophilacentral nervous system midline cells[J].Proc Natl Acad Sci, 1997, 94: 5131-5136.
[22] AGARWAL M K, RUHUL AMIN A R, AGARWA L M L.DNA replication licensing factor minichromosome maintenance deficient 5 rescues p53-mediated growth arrest[J].Cancer Res, 2007, 67: 116-121.
[23] POLLOK S, GROSSE F.Cdc45 degradation during differentiation and apoptosis[J].Biochem Biophys Res Commun, 2007, 362: 910-915.
[24] HARDY C F.Identification of Cdc45p, an essential factor required for DNA replication[J].Gene, 1997, 187: 239-246.
[25] KRAUSE K, WASNER M, REINHARD W, et al.The tumour suppressor protein p53 can repress transcription of cyclin B[J].Nucleic Acids Res, 2000, 28: 4410-4418.
[26] SUGIHARA Y, HONDA H, IIDA T, et al.Proteomic analysis of rodent ribosomes revealed heterogeneity including ribosomal proteins L10-like, L22-like 1, and L39-like[J].J Proteome Res, 2010, 9: 1351-1366.
[27] CHEESEMAN I M, ANDERSON S, JWA M, et al.Phospho-regulation of kinetochore-microtubule attachments by the Aurora kinase Ipl1p[J].Cell, 2002, 111: 163-172.
[28] BOLTON M A, LAN W, POWERS S E, et al.Aurora B kinase exists in a complex with survivin and INCENP and its kinase activity is stimulated by survivin binding and phosphorylation[J].Mol Biol Cell, 2002, 13: 3064-3077.
[29] RAJ D, LIU T, SAMADASHWILY G, et al.Survivin repression by p53, Rb and E2F2 in normal human melanocytes[J].Carcinogenesis, 2008, 29: 194-201.
