鎂缺乏和過量脅迫對紐荷爾臍橙葉綠素?zé)晒馓匦缘挠绊?/h1>
2014-08-04 02:25:40凌麗俐彭良志吳玉婷江才倫淳長品
生態(tài)學(xué)報 2014年7期 關(guān)鍵詞:影響
凌麗俐,黃 翼,彭良志,*,吳玉婷,江才倫,曹 立,淳長品
(1.西南大學(xué)柑桔研究所/中國農(nóng)業(yè)科學(xué)院柑桔研究所,重慶400712;2.國家柑桔工程技術(shù)研究中心,重慶400712;3.西南大學(xué)園林園藝學(xué)院,重慶400712)
臍橙是當(dāng)今我國栽培規(guī)模最大的甜橙品種,中國已成為世界最大的臍橙產(chǎn)地,其中絕大部分品種為紐荷爾臍橙。但是,隨著生產(chǎn)上氮、磷、鉀化肥用量的增加和有機(jī)肥用量的減少,紐荷爾臍橙葉片缺鎂黃化現(xiàn)象在贛南等我國主要臍橙產(chǎn)區(qū)普遍存在[1-2],而且呈逐年加重趨勢,已成為阻礙臍橙產(chǎn)量和品質(zhì)提高的重要因素之一。鎂是植物生長的必需營養(yǎng)元素之一,是葉綠素的中心分子,可促進(jìn)光合碳同化[3],對于類囊體膜穩(wěn)定和跨膜電子梯度的建立具有十分重要的作用[4],而且在作物產(chǎn)量提高和品質(zhì)改善等方面也發(fā)揮著重要作用[5]。目前,缺鎂脅迫對植物光合生理影響的研究主要集中在龍眼[6]、黃瓜[7-8]、菜薹[9]、甜菜[10]、水稻[11]、擬南芥[12]和煙草[13]等方面。研究表明缺鎂對植物光合膜的垛疊、激發(fā)能在光系統(tǒng)I(PSⅠ)和光系統(tǒng)Ⅱ(PSⅡ)兩個光系統(tǒng)之間的分配、PSⅡ原初光能轉(zhuǎn)化效率、光合電子傳遞速率和光合碳代謝等一系列重要的生理生化過程都有顯著的影響。生產(chǎn)上,缺鎂臍橙主要采用增施無機(jī)鎂肥(鈣鎂磷肥、硫酸鎂、白云石、含鎂石灰)和有機(jī)肥料進(jìn)行矯治。但是,由于土壤類型和植株的差異性,常常難以做到平衡施肥,鎂肥過多施用時出現(xiàn)過量脅迫,影響植物的光合生理特性,導(dǎo)致植株生長發(fā)育受阻[8]。研究表明高鎂能導(dǎo)致煙草量子產(chǎn)量降低,光合電子傳遞受阻,減少對過剩的激發(fā)能的耗散,使植株對強(qiáng)光的保護(hù)性調(diào)節(jié)能力降低[13]。缺鎂紐荷爾臍橙的平衡矯治技術(shù)的實現(xiàn)必須先解決兩個問題,一為缺鎂植株不同葉齡葉片生理特征的受影響程度是否有差異?差異度多大?二為鎂過量施加是否會對紐荷爾臍橙植株生理特征產(chǎn)生影響?特別是對不同葉齡葉片的影響有何差異?目前,有關(guān)這方面的研究尚未見報道。本文研究了鎂缺乏和過量脅迫條件下紐荷爾臍橙不同葉齡葉片的葉綠素合成與熒光特性的動態(tài)變化,以期為紐荷爾臍橙的平衡施肥矯治技術(shù)和高產(chǎn)優(yōu)質(zhì)生產(chǎn)提供理論依據(jù)。
1 材料與方法
1.1 實驗材料
試驗于重慶市北碚區(qū)歇馬鎮(zhèn)中國農(nóng)業(yè)科學(xué)院柑桔研究所臍橙課題盆栽場內(nèi)進(jìn)行。供試品種為2齡枳[Poncirus trifoliate(L.)] 砧紐荷爾臍橙(Citrus sinensis Osbeck cv.‘Newhall’)嫁接苗,選取砧木直徑約1.5 cm、高度約35 cm生長良好且較一致的嫁接苗,將植株地上部剪至高25 cm,留5片2010年秋梢葉片,主根剪至5 cm。2011年3月23日定植于盆中,每盆一株,盆中石英砂與珍珠巖(體積比1∶1)8 kg,盆底放托盤,自然光照條件下培養(yǎng)。
1.2 實驗設(shè)計
春梢萌發(fā)初期(4月15日)用改良的Hoagland-Aron營養(yǎng)液配方進(jìn)行鎂梯度營養(yǎng)液脅迫處理。完全營養(yǎng)液配方為:Ca(NO3)2·4H2O 4 mmol/L,KNO36 mmol/L,NH4H2PO41 mmol/L,MgSO4·7H2O 2 mmol/L,H3BO346 μmol/L,MnCl2·4H2O 6 μmol/L,ZnSO4·7H2O 0.7 μmol/L,CuSO4·5H2O 0.3 μmol/L,H2MoO41 μmol/L,F(xiàn)e-EDTA 50 μmol/L,pH值6.0。實驗用營養(yǎng)液的大中量元素為完全營養(yǎng)液配方的1/2濃度。試驗設(shè)3個鎂水平,即低鎂(0.1 mmol/L)、正常供鎂(1 mmol/L)和高鎂(10 mmol/L),每個水平處理15盆,共45盆。其中LM為了維持離子濃度的平衡和避免硫元素的缺乏以添加Na2SO4替代MgSO4·7H2O。實驗處理2個月內(nèi),每2 d各實驗盆補(bǔ)澆500 mL營養(yǎng)液,每7 d用去離子水洗鹽1次,防止鹽分累積。由于實驗處理到3—4個月時正值夏季,植株生長旺盛,將每2 d澆灌方式改為每日1次,每次500 mL營養(yǎng)液,每3 d用去離子水洗鹽1次。實驗分別于6月中旬(實驗處理第2月)、7月中旬(實驗處理第3月)和8月中旬(實驗處理第4月)測定植株2010年秋梢葉(2齡秋梢葉)、2011年春梢葉(1齡春梢葉)、2011年夏梢葉(1齡夏梢葉)和2011年晚夏梢葉(1齡晚夏梢葉)全部葉片的相對葉綠素含量和原初光能轉(zhuǎn)化效率(Fv/Fm)值,并分別測定2齡秋梢、1齡春梢、1齡夏梢和1齡晚夏梢基部第1葉的快速光響應(yīng)曲線。
1.3 相對葉綠素含量測定
相對葉綠素含量([Soil-plant analysis development] chlorophyll meter reading,SPAD reading)測定采用SPAD-502葉綠素測定儀(日本Konica公司)進(jìn)行,于8:30—10:30點(diǎn)在每個葉片主脈兩側(cè)中部從葉尖到葉基部均等測6個點(diǎn),取其平均值,每次測定前用標(biāo)準(zhǔn)色板校正儀器基準(zhǔn)數(shù)值。
1.4 葉綠素?zé)晒馓卣鳒y定
(1)Fv/Fm測定
葉綠素?zé)晒獠捎肞AM-2500調(diào)制葉綠素?zé)晒鈨x(德國WALZ公司)測定。測定于6:00—8:00點(diǎn)進(jìn)行,葉片暗適應(yīng)20 min后,開啟檢測光(0.15 μmol m-2s-)得到葉綠素?zé)晒鈪?shù)初始熒光(Fo),再由飽和脈沖光(4000 μmol m-2s-1,0.8 s)測得最大熒光(Fm)。根據(jù)Fo和Fm,計算Fv/Fm:
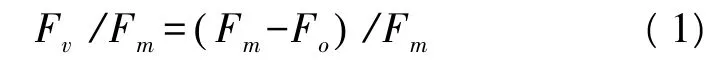
(2)快速光響應(yīng)曲線測定
每個處理選擇5株測定快速光響應(yīng)曲線。測定于8:30—10:30點(diǎn)進(jìn)行,連體葉片經(jīng)暗適應(yīng)10 s后,迅速打開葉夾,測得Fo和Fm;隨后,逐漸開啟光強(qiáng)度分別為53、93、134、178、237、408、660、820、999、1177、1387、1648 mmol m-2s-1的光化光,每個強(qiáng)度的光化光照射10 s后,經(jīng)檢測光和飽和脈沖光測得Ft、F'm,計算得到葉綠素?zé)晒馓卣鲄?shù)值,分別取平均值繪出快速光響應(yīng)曲線[14];隨后用Pam Win-3軟件依據(jù)Platt等的公式(2)對快速光響應(yīng)曲線進(jìn)行擬合[15],得到快速光響應(yīng)曲線。通過測定的葉綠素?zé)晒鈪?shù)計算出相對光合電子傳遞速率(rETRmax)和非光化學(xué)淬滅系數(shù)(NPQ)[14],公式如下:

1.5 數(shù)據(jù)處理
采用SPSS13.0軟件對實驗數(shù)據(jù)進(jìn)行處理和分析。鎂脅迫對植株相對葉綠素含量及熒光參數(shù)的影響采用單因素方差分析(one-way ANOVA)。用獨(dú)立樣本t測驗(Independent-Samples T test)檢驗各處理組植株與對照組植株間的差異。文中的圖表由Microsoft Excel軟件制作完成。
2 結(jié)果與分析
2.1 相對葉綠素含量的變化
由圖1可知,隨著實驗時間延長各實驗組2齡秋梢葉片的相對葉綠素含量均呈下降趨勢;與對照相較,低鎂導(dǎo)致SPAD值降低,而高鎂減緩了SPAD值的降低速度。第2、3和4月時低鎂組SPAD值分別比對照低1.8%(P>0.05)、12.9%(P<0.05)和12.3%(P<0.05);而高鎂組SPAD值分別為對照的101.6%(P>0.05)、97.8%(P>0.05)和107.3%(P<0.05)。
由圖1可知,前3個月各實驗組1齡春梢葉片的SPAD值均呈顯著的增加趨勢,但是到第4個月時低鎂組和高鎂組葉片SPAD值降低,分別為對照的89.5%(P<0.05)和95.5(P>0.05)。
由圖1可知,隨著實驗時間的延長,對照和低鎂組1齡夏梢葉片SPAD值呈顯著的增加趨勢,而高鎂組SPAD值無顯著變化。第3月和4月時低鎂組SPAD值分別比對照低3.1%(P>0.05)和4.0%(P>0.05),高鎂組SPAD值分別比對照低2.8%(P>0.05)和11.1%(P<0.05)。
由圖1可知,低鎂組和高鎂組1齡晚夏梢葉片的SPAD值分別比對照低4.0%(P>0.05)和7.5%(P<0.05)。
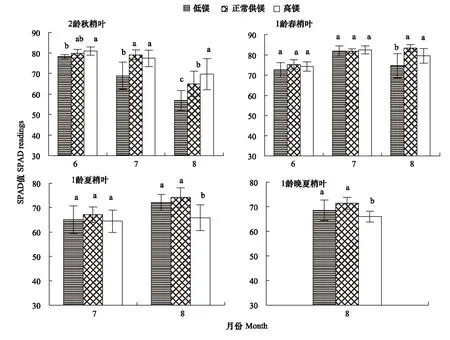
圖1 鎂脅迫對紐荷爾臍橙不同葉齡葉片相對葉綠素含量的影響Fig.1 The effect of Mg stress on relative chlorophyll content in leaves at different foliar age of Newhall navel orange
2.2 Fv/Fm的變化
Fv/Fm反映了當(dāng)所有的光系統(tǒng)Ⅱ(PSⅡ)反應(yīng)中心均處于開放態(tài)時的量子產(chǎn)量,可以直接作為原初光化學(xué)效率的指標(biāo)[16]。由圖2可知,低鎂和高鎂均導(dǎo)致不同葉齡葉片的Fv/Fm降低。對于2齡秋梢來說,實驗前2個月低鎂組Fv/Fm與對照無顯著差異,但是第3月和第4月時低鎂組Fv/Fm分別比對照低7.5%(P<0.05)和13.9%(P<0.05);而高鎂組Fv/Fm與對照無顯著差異(圖2)。對于1齡春梢來說,實驗前3個月低鎂組Fv/Fm與對照無顯著差異,但是第4月時前者比后者低12.6%(P<0.05);高鎂組Fv/Fm分別比對照低1.0%,1.4%和2.2%,無顯著差異(P>0.05)(圖2)。對于1齡夏梢葉和晚夏梢葉來說,低鎂和高鎂均導(dǎo)致Fv/Fm呈降低趨勢,但是與對照的差異不顯著(圖2)。
2.3 快速光響應(yīng)曲線的變化
2.3.1 光強(qiáng)-rETR曲線的變化
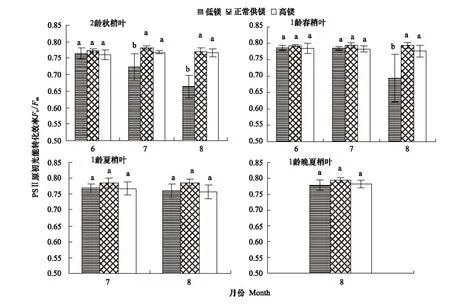
圖2 鎂脅迫對紐荷爾臍橙不同葉齡葉片F(xiàn)v/Fm的影響Fig.2 The effect of Mg stress on Fv/Fmin leaves at different foliar age of Newhall navel orange
rETR是反映實際光強(qiáng)下PSⅡ的表觀電子傳遞效率,用于度量光化學(xué)反應(yīng)導(dǎo)致碳固定的電子傳遞情況[17]。由圖3可知,實驗期間各實驗組2齡秋梢葉片的rETR均呈降低趨勢。第2月、3月和4月時,對照組rETRmax分別為81.6 μmol m-2s-1、69.0 μmol m-2s-1和59.5 μmol m-2s-1;與對照相較,低鎂組rETRmax分別降低3.4%(P>0.05)、22.9%(P<0.05)和35.7%(P<0.05),而高鎂組rETRmax分別為對照的101.1%、96.4%和110.1%,差異不顯著(P>0.05)。
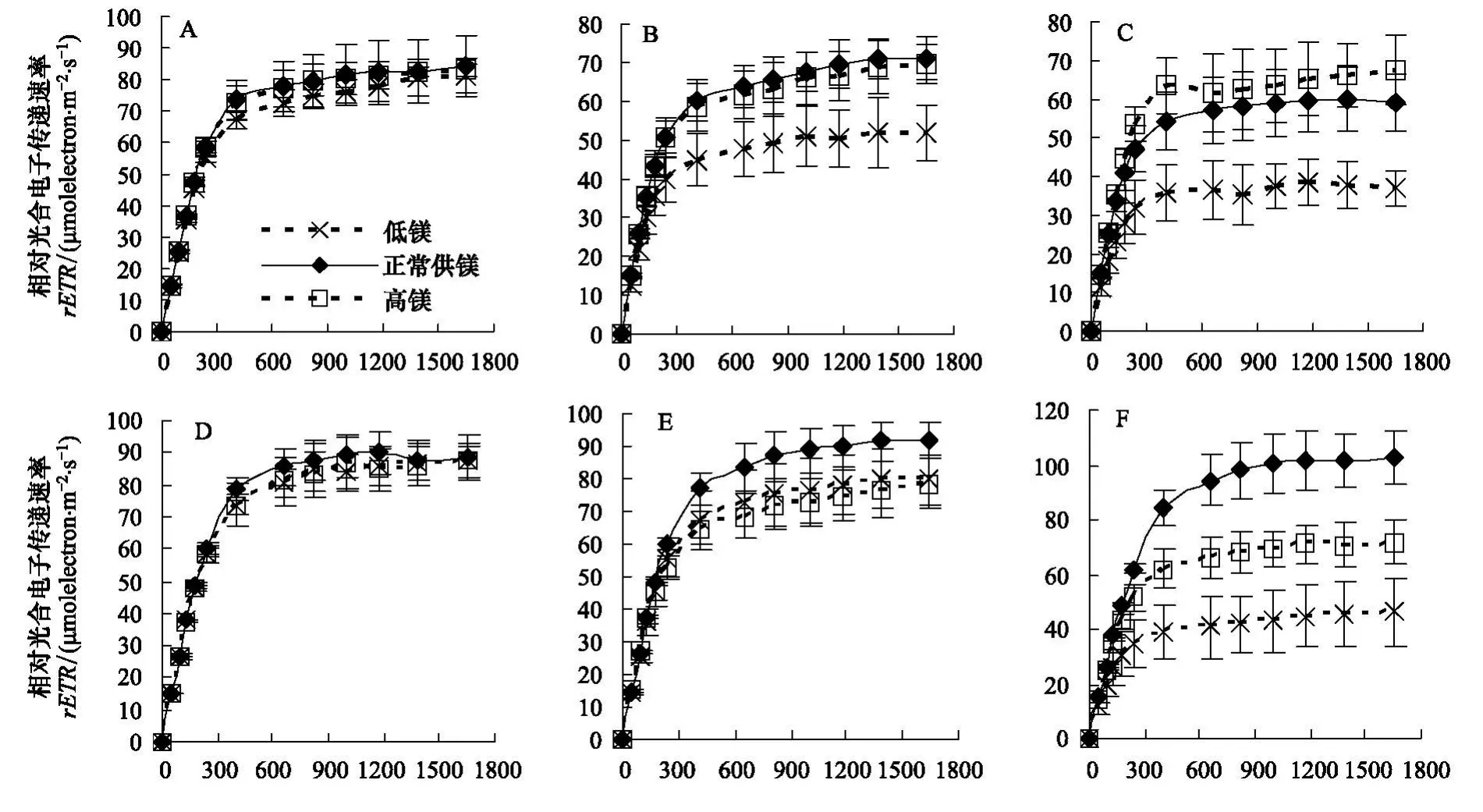
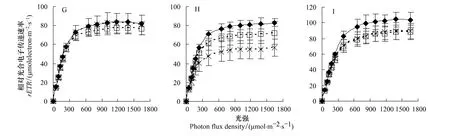
圖3 鎂脅迫對紐荷爾臍橙不同葉齡葉片的相對電子傳遞速率(rETR)的影響Fig.3 The effect of Mg stress on relative electron transport ratio(rETR)in leaves at different foliar age of Newhall navel orange
由圖3可知,隨著實驗時間增加,對照組1齡春梢和1齡夏梢葉片的rETR呈上升趨勢,而低鎂組和高鎂組的rETR呈顯著降低趨勢。對于1齡春梢來說,第2月、3月和4月時,對照組rETRmax分別為90.0 μmol m-2s-1、91.0 μmol m-2s-1和97.6 μmol m-2s-1,而低鎂組rETRmax分別比對照低4.6%(P>0.05)、24.6%(P<0.05)和56.2%(P<0.05),高鎂組rETRmax分別比對照低3.6%(P>0.05)、17.5%(P<0.05)和31.2%(P<0.05)。對于1齡夏梢來說,第4月時低鎂組和高鎂組rETRmax分別比對照低32.6%和12.8%,差異顯著(P<0.05)。對于1齡晚夏梢來說,低鎂組和高鎂組的rETRmax分別比對照低15.2%和15.5%,差異顯著(P<0.05)。
2.3.1 光強(qiáng)-NPQ曲線的變化
NPQ在強(qiáng)光下可以耗散過剩的激發(fā)能,減少過剩光能對PSⅡ反應(yīng)中心的破壞,可有效的保護(hù)光合系統(tǒng)不受強(qiáng)光的破壞[18]。由圖4可知,對于2齡秋梢來說,實驗2個月內(nèi)低鎂和高鎂對NPQ的影響不顯著;第3個月時低鎂組NPQ顯著增大,而高鎂組NPQ顯著降低,特別是第4個月時低鎂組NPQ顯著降低,其最大值僅為對照的52.8%。
由圖4可知,低鎂和高鎂導(dǎo)致1齡春梢、1齡夏梢和1齡晚夏梢葉片NPQ顯著增加,但是第4個月時低鎂1齡春梢和1齡夏梢葉片的NPQ顯著降低,其最大值分別僅為對照的73.4%、80.2%。
3 討論
3.1 鎂缺乏和過量脅迫對紐荷爾臍橙不同葉齡葉片光合色素合成的影響
鎂是構(gòu)成葉綠素的中心元素,其含量與相對葉綠素呈極顯著相關(guān)[19]。Schimansky用放射性同位素28Mg對菜豆、大麥進(jìn)行示蹤,證明鎂可以通過韌皮部由老葉轉(zhuǎn)移至新葉[20]。對大豆[21]和黃瓜[8]的研究表明,葉片葉綠素含量隨根系供鎂含量的增加呈先上升后下降的趨勢;雖然缺鎂黃瓜葉片葉綠素a、葉綠素b、胡蘿卜素含量均極顯著降低,但是其葉綠素a/b值最高。Hermans等的研究表明缺鎂脅迫引起葉綠體結(jié)構(gòu)與功能異常、葉綠素含量下降、葉片異常黃化,進(jìn)而直接影響PSⅠ和PSⅡ的功能[10]。申燕等對“春見”橘橙的研究表明,缺鎂脅迫導(dǎo)致葉綠體內(nèi)類囊體片層模糊,片層數(shù)明顯減少,線粒體膜模糊且出現(xiàn)異常的黑色顆粒,葉綠素含量降低;鎂過量脅迫會導(dǎo)致淀粉粒和質(zhì)體小球異常增多增大,線粒體膜模糊,葉綠體內(nèi)片層消失,葉綠素含量顯著降低[22]。本文研究表明,缺鎂不僅會導(dǎo)致老葉(2齡秋梢葉)葉綠素含量降低,也會導(dǎo)致新葉(1齡春梢葉、1齡夏梢葉和1齡晚夏梢葉)葉綠素含量降低;雖然短期(2個月)缺鎂對2齡秋梢葉和1齡春梢葉葉綠素的合成影響不顯著,但是隨著缺鎂脅迫時間的增加植株葉綠素合成受抑制程度顯著增大。相對于鎂缺乏脅迫來說,鎂過量脅迫對減緩老葉和1齡春梢葉的葉綠素含量下降具有顯著效果,而其對新葉(1齡夏梢葉和1齡晚夏梢葉)葉綠素合成的影響則相反,其機(jī)制是元素拮抗還是其他原因有待進(jìn)一步的探討。
3.2 鎂缺乏和過量脅迫對紐荷爾臍橙不同葉齡葉片光合作用和早衰的影響
鎂脅迫不僅影響植株的葉綠素合成,而且也會影響植株的光合作用。缺鎂脅迫越嚴(yán)重,植物的光合作用效率就越低。現(xiàn)有的研究表明,缺鎂會引起光合膜垛疊受阻,降低PSⅠ與PSⅡ相對熒光產(chǎn)量的比值[23-24];缺鎂黃瓜葉片凈光合速率(Pn)、氣孔導(dǎo)度(Gs)和蒸騰速率(Tr)的日變化呈單峰曲線,易發(fā)生顯著的光抑制,葉綠素?zé)晒鈪?shù)Fv/Fm、Fv/Fo、qP和qN降低[7];缺鎂龍眼光補(bǔ)償點(diǎn)和CO2補(bǔ)償點(diǎn)提高,光飽和點(diǎn)和CO2飽和點(diǎn)下降,PSⅡ活性下降[6]。

圖4 鎂脅迫對紐荷爾臍橙不同葉齡葉片的非光化學(xué)淬滅(NPQ)的影響Fig.4 The effect of Mg stress on non-photoche-mical quenching(NPQ)in leaves at different foliar age of Newhall navel orange
本研究表明,隨著缺鎂時間的延長,紐荷爾臍橙新葉和老葉Fv/Fm受脅迫程度呈顯著增大趨勢;老葉Fv/Fm被抑制的表現(xiàn)時間顯著早于新葉,而且其Fv/Fm降低程度顯著大于新葉。此外,PSⅡ光能活化、活性氧代謝與植物葉片的早衰有密切的聯(lián)系[11,25]。在水稻葉片的早衰研究中發(fā)現(xiàn),在衰老過程中劍葉的Fv/Fm、qP、ФPSⅡ、rETR逐漸下降,qN上升,過剩的光能導(dǎo)致活性氧、H2O2和膜脂過氧化產(chǎn)物MDA的積累,導(dǎo)致葉綠素衰減,發(fā)生光氧化早衰[26]。在夏季強(qiáng)光照條件下葉片吸收的光能如不能完全通過光化學(xué)、非光化學(xué)和光子躍遷淬滅,過量的光能將會對光系統(tǒng)Ⅱ產(chǎn)生傷害,導(dǎo)致葉綠素含量降低,顯現(xiàn)黃化早衰癥狀[11]。缺鎂至第3月時紐荷爾臍橙新葉和老葉rETR呈顯著下降趨勢,第4月時2齡秋梢、1齡春梢葉和1齡夏梢葉的NPQ不增反降,熱耗散能力顯著降低。表明,在夏季高光條件下缺鎂植株吸收的過量光能很難以熱的形式耗散掉,易受到光抑制,產(chǎn)生光傷害;結(jié)合2齡秋梢和1齡春梢葉Fv/Fm的顯著降低結(jié)果可知,夏季高光已導(dǎo)致缺鎂紐荷爾臍橙葉片受到光傷害。
鎂缺乏和過量脅迫均能使煙草葉片量子產(chǎn)量、光合電子傳遞速率、非光化學(xué)猝滅降低,使植株對強(qiáng)光的保護(hù)性調(diào)節(jié)能力下降。鎂過量脅迫下黃瓜Pn和Tr呈雙峰曲線,胞間CO2濃度(Ci)、Gs呈近似倒拋物線型,光飽和點(diǎn)、CO2飽和點(diǎn)以及表觀量子效率和羧化效率都大于缺鎂脅迫[13]。本研究表明,與鎂缺乏相比,鎂過量對紐荷爾臍橙Fv/Fm的脅迫影響顯著較低,這與植株rETR和NPQ的光響應(yīng)能力變化有關(guān)。短期(2個月)鎂過量脅迫對紐荷爾臍橙2齡秋梢和1齡春梢葉rETR和NPQ的光響應(yīng)能力無顯著影響。雖然隨著脅迫時間的增加,新葉(1齡春梢葉、1齡夏梢葉和1齡晚夏梢葉)的rETR均呈顯著降低趨勢,但是其NPQ均高于對照,熱耗散能力較高,不易受到過量光能的抑制,產(chǎn)生光傷害。對于紐荷爾臍橙植株老葉來說,雖然自第3個月開始NPQ低于對照,熱耗散能力有所下降,但是其rETR與對照無顯著差異,也不易受到過量光能的抑制,不易產(chǎn)生光傷害。
綜上所述,缺鎂脅迫加速2齡秋稍葉和1齡春梢葉的衰老,而鎂過量脅迫延緩其衰老;夏季高光易導(dǎo)致缺鎂脅迫臍橙葉片受到光傷害,而對鎂過量脅迫臍橙葉片的影響較小,不易產(chǎn)生光傷害。因此,生產(chǎn)上應(yīng)重視鎂肥的施用。由本研究結(jié)果可知,缺鎂脅迫對紐荷爾臍橙植株的不利影響顯著大于鎂過量脅迫;而且生產(chǎn)上一般的鎂肥過量應(yīng)不會對樹體產(chǎn)生顯著的不利影響。
[1] Peng L Z,Lai J J,Chun C P,Cao L,Zhong L H,Jiang C L,Ling L L.Role of compound magnesium fertilizer in correcting leaf yellowing caused by nutrient element deficiency on navel orange in Gannan.South China Fruits,2008,37(3):10-12.
[2] Li J,Xie Z C,Xie W L,Wu X M,Shi Q.Relationship between leafveinsplittingandmineralnutritionofcitrus.Acta Horticulturae Sinica,2011,38(3):425-433.
[3] Wu W H,Peters J,Berkowitz G A.Surface charge mediated effects of Mg2+on K+flux across the chloroplast envelope are associated with regulation of stromal pH and photosynthesis.Plant Physiology,1991,97(2):580-587.
[4] Hao N P,Li T Z,Zhang Q D,Lou S Q,Tang C Q,Kuang T Y.Structure and function of chloroplast membranesⅧ.Effect of magnesium ions on the polymerization of chlorophyll-protein complexesofchloroplastmembranes.ActaBiochimicaet Biophysica Sinica,1981,13(4):365-372.
[5] Sun N,Zeng X B,Gao J S,Wang B R.Effects of magnesium compound fertilizer on daylily(Hemerocallis citrina Baroni)growth and soil Nutrients.Scientia Agricultura Sinica,2006,39(1):95-101.
[6] Li Y,Liu X H,Zhuang W M.The effect of magnesium deficiency on photosynthesis of Longan(Dimocarpus long ana Lour.)seedlings.Acta Horticulturae Sinica,2001,28(2):101-106.
[7] Yang G D,Zhu Z J,Ji Y M.Effect of light intensity and magnesium deficiency on chlorophyll fluorescence and active oxygen in cucumber leaves.Plant Nutrition and Fertilizer Science,2002,8(1):115-118.
[8] Xie X Y,Liu H T,Cheng Z W.Effects of magnesium stress on photosynthetic characterofcucumberingreenhouse.China Vegetables,2009,(6):36-40.
[9] Liu H C,Chen X M,Chen R Y,Song S W,Sun G W.Effects of magnesium deficiency on photosynthesis characteristic of flowering Chinese cabbage.Horticulturae Sinica,2006,33(2):311-316.
[10] Hermans C,Johnson G N,Strasser R J,Verbruggen N.Physiological characterisation of magnesium deficiency in sugar beet:Acclimationtolowmagnesiumdifferentiallyaffects photosystemsⅠandⅡ.Planta,2004,220(2):344-355.
[11] Yang Y,Jiang D A,Sun J W,Huang Z A,Jin S H.Effects of different magnesium nutrition levels on chlorophyll fluorescence characteristics and excitation energy dissipation in rice leaves.Plant Nutrition and Fertilizer Science,2005,11(1):79-86.
[12] Hermans C,Verbruggen N.Physiological characterization of Mg deficiency inArabidopsisthaliana.JournalofExperimental Botany,2005,56(418):2153-2161.
[13] Guan G S,Tu N M,Xiao H Q,Zhu L S,Zhu Y H,Wang Z M.Effectsofmagnesiumontobaccogrowthandchlorophyll fluorescence parameters of tobacco leaves.Plant Nutrition and Fertilizer Science,2008,14(1):151-155.
[14] Scheriber U,Gademann R,Ralph P J,Larkum A W D.AssessmentofphotosyntheticperformanceofProchloronin Lissoclinumpatellainhospitebychlorophyllfluorescence measurements.Plant and Cell Physiology,1997,38(8):945-951.
[15] Platt T,Gallegos C L,Harrison W G.Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton.Journal of Marine Research,1980,38(4):687-701.
[16] Demmig-Adams B,Adams W WⅢ,Barker D H,Logan B A,Bowling D R,Verhoeven A S.Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation.Physiologia Plantarum,1996,98(2):253-264.
[17] Genty B,Briantais J M,Baker N R.The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence.Biochimica et Biophysica Acta,1989,990(1):87-92.
[18] Horton P,Ruban A V,Walters R G.Regulation of light harvesting in green plants(indicationby nonphotochemical quenching of chlorophyll fluorescence).Plant Physiology,1994,106(2):415-420.
[19] Zhang G Y,Peng L Z,Chun C P,Zeng M,Ling L L,Lai J J,Wang Z X.Seasonal changes in leaf magnesium and boron contents and their relationships to leaf yellowing of navel orange(Citrus sinensis Osbeck).Acta Horticulturae Sinica,2010,37(8):1317-1324.
[20] Schimansky C.Possibilities and limitations of utilizing28Mg for studying Mg-specific problems in higher plants.Journal of Plant Nutrition,1985,8(6):467-489.
[21] Li Y Z,Li F,Gao M,Chen X,Lv C L,F(xiàn)u H L,Sun D Q.Effects of soil magnesium excess on several physiological indexes of soybean.Agro-Environmental Protection,1990,9(2):41-42.
[22] Shen Y,Xiao J X,Yang H,Zhang S L.Effects of magnesium stress on growth,distribution of several mineral elements and leaf ultrastructure of‘Harumi’tangor.Acta Horticulturae Sinica,2011,38(5):849-858.
[23] Zhang Q D,Tang C Q,Li S Y.Structure and function of chloroplast membraneⅪ.The effects of linolenic acid on the structure and the absorption and fluorescence spectra of wheat chloroplast membranes as well as regulations by MgCl2.Acta Botanica Sinica,1982,24(4):326-333.
[24] Lin S Q,Xu C H,Zhang Q D,Xu L,Mao D Z,Kuang T Y.Some application of chlorophyll fluorescence kinetics to plant stress physiology to ecology and agricultural modernization.Chinese Bulletin of Botany,1992,9(1):1-16.
[25] Li Y,Liu X H.Effects of magnesium deficiency on senescence of Dimocarpus longana leaves.Chinese Journal of Applied Ecology,2002,13(3):311-314.
[26] Jiao D M,Li X,Huang X Q,Ji B H.The relationship among photoinhibition,Photooxidationandearlyagingatlater developmental stages in different high yield varieties.Scientia Agricultura Sinica,2002,35(5):487-492.
參考文獻(xiàn):
[1] 彭良志,賴九江,淳長品,曹立,鐘莉華,江才倫,凌麗俐.復(fù)合鎂肥對贛南臍橙缺素黃化的矯治作用.中國南方果樹,2008,37(3):10-12.
[2] 李健,謝鐘琛,謝文龍,吳應(yīng)明,施清.柑橘葉脈開裂癥與礦質(zhì)營養(yǎng)的關(guān)系.園藝學(xué)報,2011,38(3):425-433.
[4] 郝廼斌,李桐柱,張其德,婁世慶,唐崇欽,匡廷云.葉綠體膜的結(jié)構(gòu)與功能Ⅷ.鎂離子對葉綠體類囊體膜的葉綠素-蛋白復(fù)合體聚合的影響.生物化學(xué)與生物物理學(xué)報,1981,13(4):365-372.
[5] 孫楠,曾希柏,高菊生,王伯仁.含鎂復(fù)合肥對黃花菜生長及土壤養(yǎng)分含量的影響.中國農(nóng)業(yè)科學(xué),2006,39(1):95-101.
[6] 李延,劉星輝,莊衛(wèi)民.缺鎂對龍眼光合作用的影響.園藝學(xué)報,2001,28(2):101-106.
[7] 楊廣東,朱祝軍,計玉妹.不同光強(qiáng)和缺鎂脅迫對黃瓜葉片葉綠素?zé)晒馓匦院突钚匝醍a(chǎn)生的影響.植物營養(yǎng)與肥料學(xué)報,2002,8(1):115-118.
[8] 謝小玉,劉海濤,程志偉.鎂對溫室黃瓜光合特性的影響.中國蔬菜,2009,(6):36-40.
[9] 劉厚誠,陳細(xì)明,陳日遠(yuǎn),宋世威,孫光聞.缺鎂對菜薹光合作用特性的影響.園藝學(xué)報,2006,33(2):311-316.
[11] 楊勇,蔣德安,孫駿威,黃宗安,金松恒.不同供鎂水平對水稻葉片葉綠素?zé)晒馓匦院湍芰亢纳⒌挠绊?植物營養(yǎng)與肥料學(xué)報,2005,11(1):79-86.
[13] 關(guān)廣晟,屠乃美,肖漢乾,朱列書,朱英華,王中美.鎂對煙草生長及葉片葉綠素?zé)晒鈪?shù)的影響.植物營養(yǎng)與肥料學(xué)報,2008,14(1):151-155.
[19] 張廣越,彭良志,淳長品,曾明,凌麗俐,賴九江,王振興.臍橙葉片鎂、硼含量變化與缺素黃化的關(guān)系.園藝學(xué)報,2010,37(8):1317-1324.
[21] 李亞洲,李沸,高銘,陳旭,呂春莉,付海龍,孫大群.土壤中過量鎂對大豆幾項生理指標(biāo)的影響.農(nóng)業(yè)環(huán)境保護(hù),1990,9(2):41-42.
[22] 申燕,肖家欣,楊慧,張紹鈴.鎂脅迫對‘春見’橘橙生長和礦質(zhì)元素分布及葉片超微結(jié)構(gòu)的影響.園藝學(xué)報,2011,38(5):849-858.
[23] 張其德,唐崇欽,李世儀.葉綠體膜的結(jié)構(gòu)與功能Ⅺ.亞麻酸對小麥葉綠體膜結(jié)構(gòu)、吸收光譜和熒光光譜的影響以及鎂離子的調(diào)節(jié)作用.植物學(xué)報,1982,24(4):326-333.
[24] 林世青,許春輝,張其德,徐黎,毛大璋,匡廷云.葉綠素?zé)晒鈩恿W(xué)在植物抗性生理學(xué)、生態(tài)學(xué)和農(nóng)業(yè)現(xiàn)代化中的應(yīng)用.植物學(xué)通報,1992,9(1):1-16.
[25] 李延,劉星輝.缺鎂脅迫對龍眼葉片衰老的影響.應(yīng)用生態(tài)學(xué)報,2002,13(3):311-314.
[26] 焦德茂,李霞,黃雪清,季本華.不同高產(chǎn)水稻品種生育后期葉片光抑制、光氧化和早衰的關(guān)系.中國農(nóng)業(yè)科學(xué),2002,35(5):487-492.
猜你喜歡
是什么影響了滑動摩擦力的大小中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08 哪些顧慮影響擔(dān)當(dāng)?當(dāng)代陜西(2021年2期)2021-03-29 07:41:24 影響大師家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18 沒錯,痛經(jīng)有時也會影響懷孕媽媽寶寶(2017年3期)2017-02-21 01:22:28 擴(kuò)鏈劑聯(lián)用對PETG擴(kuò)鏈反應(yīng)與流變性能的影響中國塑料(2016年3期)2016-06-15 20:30:00 基于Simulink的跟蹤干擾對跳頻通信的影響通信電源技術(shù)(2016年3期)2016-03-26 07:13:38 如何影響他人知識經(jīng)濟(jì)·中國直銷(2016年3期)2016-02-27 16:15:49 APRIL siRNA對SW480裸鼠移植瘤的影響現(xiàn)代檢驗醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04 對你有重要影響的人閱讀與作文(小學(xué)低年級版)(2011年3期)2011-01-01 00:00:00
凌麗俐,黃 翼,彭良志,*,吳玉婷,江才倫,曹 立,淳長品
(1.西南大學(xué)柑桔研究所/中國農(nóng)業(yè)科學(xué)院柑桔研究所,重慶400712;2.國家柑桔工程技術(shù)研究中心,重慶400712;3.西南大學(xué)園林園藝學(xué)院,重慶400712)
臍橙是當(dāng)今我國栽培規(guī)模最大的甜橙品種,中國已成為世界最大的臍橙產(chǎn)地,其中絕大部分品種為紐荷爾臍橙。但是,隨著生產(chǎn)上氮、磷、鉀化肥用量的增加和有機(jī)肥用量的減少,紐荷爾臍橙葉片缺鎂黃化現(xiàn)象在贛南等我國主要臍橙產(chǎn)區(qū)普遍存在[1-2],而且呈逐年加重趨勢,已成為阻礙臍橙產(chǎn)量和品質(zhì)提高的重要因素之一。鎂是植物生長的必需營養(yǎng)元素之一,是葉綠素的中心分子,可促進(jìn)光合碳同化[3],對于類囊體膜穩(wěn)定和跨膜電子梯度的建立具有十分重要的作用[4],而且在作物產(chǎn)量提高和品質(zhì)改善等方面也發(fā)揮著重要作用[5]。目前,缺鎂脅迫對植物光合生理影響的研究主要集中在龍眼[6]、黃瓜[7-8]、菜薹[9]、甜菜[10]、水稻[11]、擬南芥[12]和煙草[13]等方面。研究表明缺鎂對植物光合膜的垛疊、激發(fā)能在光系統(tǒng)I(PSⅠ)和光系統(tǒng)Ⅱ(PSⅡ)兩個光系統(tǒng)之間的分配、PSⅡ原初光能轉(zhuǎn)化效率、光合電子傳遞速率和光合碳代謝等一系列重要的生理生化過程都有顯著的影響。生產(chǎn)上,缺鎂臍橙主要采用增施無機(jī)鎂肥(鈣鎂磷肥、硫酸鎂、白云石、含鎂石灰)和有機(jī)肥料進(jìn)行矯治。但是,由于土壤類型和植株的差異性,常常難以做到平衡施肥,鎂肥過多施用時出現(xiàn)過量脅迫,影響植物的光合生理特性,導(dǎo)致植株生長發(fā)育受阻[8]。研究表明高鎂能導(dǎo)致煙草量子產(chǎn)量降低,光合電子傳遞受阻,減少對過剩的激發(fā)能的耗散,使植株對強(qiáng)光的保護(hù)性調(diào)節(jié)能力降低[13]。缺鎂紐荷爾臍橙的平衡矯治技術(shù)的實現(xiàn)必須先解決兩個問題,一為缺鎂植株不同葉齡葉片生理特征的受影響程度是否有差異?差異度多大?二為鎂過量施加是否會對紐荷爾臍橙植株生理特征產(chǎn)生影響?特別是對不同葉齡葉片的影響有何差異?目前,有關(guān)這方面的研究尚未見報道。本文研究了鎂缺乏和過量脅迫條件下紐荷爾臍橙不同葉齡葉片的葉綠素合成與熒光特性的動態(tài)變化,以期為紐荷爾臍橙的平衡施肥矯治技術(shù)和高產(chǎn)優(yōu)質(zhì)生產(chǎn)提供理論依據(jù)。
1 材料與方法
1.1 實驗材料
試驗于重慶市北碚區(qū)歇馬鎮(zhèn)中國農(nóng)業(yè)科學(xué)院柑桔研究所臍橙課題盆栽場內(nèi)進(jìn)行。供試品種為2齡枳[Poncirus trifoliate(L.)] 砧紐荷爾臍橙(Citrus sinensis Osbeck cv.‘Newhall’)嫁接苗,選取砧木直徑約1.5 cm、高度約35 cm生長良好且較一致的嫁接苗,將植株地上部剪至高25 cm,留5片2010年秋梢葉片,主根剪至5 cm。2011年3月23日定植于盆中,每盆一株,盆中石英砂與珍珠巖(體積比1∶1)8 kg,盆底放托盤,自然光照條件下培養(yǎng)。
1.2 實驗設(shè)計
春梢萌發(fā)初期(4月15日)用改良的Hoagland-Aron營養(yǎng)液配方進(jìn)行鎂梯度營養(yǎng)液脅迫處理。完全營養(yǎng)液配方為:Ca(NO3)2·4H2O 4 mmol/L,KNO36 mmol/L,NH4H2PO41 mmol/L,MgSO4·7H2O 2 mmol/L,H3BO346 μmol/L,MnCl2·4H2O 6 μmol/L,ZnSO4·7H2O 0.7 μmol/L,CuSO4·5H2O 0.3 μmol/L,H2MoO41 μmol/L,F(xiàn)e-EDTA 50 μmol/L,pH值6.0。實驗用營養(yǎng)液的大中量元素為完全營養(yǎng)液配方的1/2濃度。試驗設(shè)3個鎂水平,即低鎂(0.1 mmol/L)、正常供鎂(1 mmol/L)和高鎂(10 mmol/L),每個水平處理15盆,共45盆。其中LM為了維持離子濃度的平衡和避免硫元素的缺乏以添加Na2SO4替代MgSO4·7H2O。實驗處理2個月內(nèi),每2 d各實驗盆補(bǔ)澆500 mL營養(yǎng)液,每7 d用去離子水洗鹽1次,防止鹽分累積。由于實驗處理到3—4個月時正值夏季,植株生長旺盛,將每2 d澆灌方式改為每日1次,每次500 mL營養(yǎng)液,每3 d用去離子水洗鹽1次。實驗分別于6月中旬(實驗處理第2月)、7月中旬(實驗處理第3月)和8月中旬(實驗處理第4月)測定植株2010年秋梢葉(2齡秋梢葉)、2011年春梢葉(1齡春梢葉)、2011年夏梢葉(1齡夏梢葉)和2011年晚夏梢葉(1齡晚夏梢葉)全部葉片的相對葉綠素含量和原初光能轉(zhuǎn)化效率(Fv/Fm)值,并分別測定2齡秋梢、1齡春梢、1齡夏梢和1齡晚夏梢基部第1葉的快速光響應(yīng)曲線。
1.3 相對葉綠素含量測定
相對葉綠素含量([Soil-plant analysis development] chlorophyll meter reading,SPAD reading)測定采用SPAD-502葉綠素測定儀(日本Konica公司)進(jìn)行,于8:30—10:30點(diǎn)在每個葉片主脈兩側(cè)中部從葉尖到葉基部均等測6個點(diǎn),取其平均值,每次測定前用標(biāo)準(zhǔn)色板校正儀器基準(zhǔn)數(shù)值。
1.4 葉綠素?zé)晒馓卣鳒y定
(1)Fv/Fm測定
葉綠素?zé)晒獠捎肞AM-2500調(diào)制葉綠素?zé)晒鈨x(德國WALZ公司)測定。測定于6:00—8:00點(diǎn)進(jìn)行,葉片暗適應(yīng)20 min后,開啟檢測光(0.15 μmol m-2s-)得到葉綠素?zé)晒鈪?shù)初始熒光(Fo),再由飽和脈沖光(4000 μmol m-2s-1,0.8 s)測得最大熒光(Fm)。根據(jù)Fo和Fm,計算Fv/Fm:
(2)快速光響應(yīng)曲線測定
每個處理選擇5株測定快速光響應(yīng)曲線。測定于8:30—10:30點(diǎn)進(jìn)行,連體葉片經(jīng)暗適應(yīng)10 s后,迅速打開葉夾,測得Fo和Fm;隨后,逐漸開啟光強(qiáng)度分別為53、93、134、178、237、408、660、820、999、1177、1387、1648 mmol m-2s-1的光化光,每個強(qiáng)度的光化光照射10 s后,經(jīng)檢測光和飽和脈沖光測得Ft、F'm,計算得到葉綠素?zé)晒馓卣鲄?shù)值,分別取平均值繪出快速光響應(yīng)曲線[14];隨后用Pam Win-3軟件依據(jù)Platt等的公式(2)對快速光響應(yīng)曲線進(jìn)行擬合[15],得到快速光響應(yīng)曲線。通過測定的葉綠素?zé)晒鈪?shù)計算出相對光合電子傳遞速率(rETRmax)和非光化學(xué)淬滅系數(shù)(NPQ)[14],公式如下:
1.5 數(shù)據(jù)處理
采用SPSS13.0軟件對實驗數(shù)據(jù)進(jìn)行處理和分析。鎂脅迫對植株相對葉綠素含量及熒光參數(shù)的影響采用單因素方差分析(one-way ANOVA)。用獨(dú)立樣本t測驗(Independent-Samples T test)檢驗各處理組植株與對照組植株間的差異。文中的圖表由Microsoft Excel軟件制作完成。
2 結(jié)果與分析
2.1 相對葉綠素含量的變化
由圖1可知,隨著實驗時間延長各實驗組2齡秋梢葉片的相對葉綠素含量均呈下降趨勢;與對照相較,低鎂導(dǎo)致SPAD值降低,而高鎂減緩了SPAD值的降低速度。第2、3和4月時低鎂組SPAD值分別比對照低1.8%(P>0.05)、12.9%(P<0.05)和12.3%(P<0.05);而高鎂組SPAD值分別為對照的101.6%(P>0.05)、97.8%(P>0.05)和107.3%(P<0.05)。
由圖1可知,前3個月各實驗組1齡春梢葉片的SPAD值均呈顯著的增加趨勢,但是到第4個月時低鎂組和高鎂組葉片SPAD值降低,分別為對照的89.5%(P<0.05)和95.5(P>0.05)。
由圖1可知,隨著實驗時間的延長,對照和低鎂組1齡夏梢葉片SPAD值呈顯著的增加趨勢,而高鎂組SPAD值無顯著變化。第3月和4月時低鎂組SPAD值分別比對照低3.1%(P>0.05)和4.0%(P>0.05),高鎂組SPAD值分別比對照低2.8%(P>0.05)和11.1%(P<0.05)。
由圖1可知,低鎂組和高鎂組1齡晚夏梢葉片的SPAD值分別比對照低4.0%(P>0.05)和7.5%(P<0.05)。
圖1 鎂脅迫對紐荷爾臍橙不同葉齡葉片相對葉綠素含量的影響Fig.1 The effect of Mg stress on relative chlorophyll content in leaves at different foliar age of Newhall navel orange
2.2 Fv/Fm的變化
Fv/Fm反映了當(dāng)所有的光系統(tǒng)Ⅱ(PSⅡ)反應(yīng)中心均處于開放態(tài)時的量子產(chǎn)量,可以直接作為原初光化學(xué)效率的指標(biāo)[16]。由圖2可知,低鎂和高鎂均導(dǎo)致不同葉齡葉片的Fv/Fm降低。對于2齡秋梢來說,實驗前2個月低鎂組Fv/Fm與對照無顯著差異,但是第3月和第4月時低鎂組Fv/Fm分別比對照低7.5%(P<0.05)和13.9%(P<0.05);而高鎂組Fv/Fm與對照無顯著差異(圖2)。對于1齡春梢來說,實驗前3個月低鎂組Fv/Fm與對照無顯著差異,但是第4月時前者比后者低12.6%(P<0.05);高鎂組Fv/Fm分別比對照低1.0%,1.4%和2.2%,無顯著差異(P>0.05)(圖2)。對于1齡夏梢葉和晚夏梢葉來說,低鎂和高鎂均導(dǎo)致Fv/Fm呈降低趨勢,但是與對照的差異不顯著(圖2)。
2.3 快速光響應(yīng)曲線的變化
2.3.1 光強(qiáng)-rETR曲線的變化
圖2 鎂脅迫對紐荷爾臍橙不同葉齡葉片F(xiàn)v/Fm的影響Fig.2 The effect of Mg stress on Fv/Fmin leaves at different foliar age of Newhall navel orange
rETR是反映實際光強(qiáng)下PSⅡ的表觀電子傳遞效率,用于度量光化學(xué)反應(yīng)導(dǎo)致碳固定的電子傳遞情況[17]。由圖3可知,實驗期間各實驗組2齡秋梢葉片的rETR均呈降低趨勢。第2月、3月和4月時,對照組rETRmax分別為81.6 μmol m-2s-1、69.0 μmol m-2s-1和59.5 μmol m-2s-1;與對照相較,低鎂組rETRmax分別降低3.4%(P>0.05)、22.9%(P<0.05)和35.7%(P<0.05),而高鎂組rETRmax分別為對照的101.1%、96.4%和110.1%,差異不顯著(P>0.05)。
圖3 鎂脅迫對紐荷爾臍橙不同葉齡葉片的相對電子傳遞速率(rETR)的影響Fig.3 The effect of Mg stress on relative electron transport ratio(rETR)in leaves at different foliar age of Newhall navel orange
由圖3可知,隨著實驗時間增加,對照組1齡春梢和1齡夏梢葉片的rETR呈上升趨勢,而低鎂組和高鎂組的rETR呈顯著降低趨勢。對于1齡春梢來說,第2月、3月和4月時,對照組rETRmax分別為90.0 μmol m-2s-1、91.0 μmol m-2s-1和97.6 μmol m-2s-1,而低鎂組rETRmax分別比對照低4.6%(P>0.05)、24.6%(P<0.05)和56.2%(P<0.05),高鎂組rETRmax分別比對照低3.6%(P>0.05)、17.5%(P<0.05)和31.2%(P<0.05)。對于1齡夏梢來說,第4月時低鎂組和高鎂組rETRmax分別比對照低32.6%和12.8%,差異顯著(P<0.05)。對于1齡晚夏梢來說,低鎂組和高鎂組的rETRmax分別比對照低15.2%和15.5%,差異顯著(P<0.05)。
2.3.1 光強(qiáng)-NPQ曲線的變化
NPQ在強(qiáng)光下可以耗散過剩的激發(fā)能,減少過剩光能對PSⅡ反應(yīng)中心的破壞,可有效的保護(hù)光合系統(tǒng)不受強(qiáng)光的破壞[18]。由圖4可知,對于2齡秋梢來說,實驗2個月內(nèi)低鎂和高鎂對NPQ的影響不顯著;第3個月時低鎂組NPQ顯著增大,而高鎂組NPQ顯著降低,特別是第4個月時低鎂組NPQ顯著降低,其最大值僅為對照的52.8%。
由圖4可知,低鎂和高鎂導(dǎo)致1齡春梢、1齡夏梢和1齡晚夏梢葉片NPQ顯著增加,但是第4個月時低鎂1齡春梢和1齡夏梢葉片的NPQ顯著降低,其最大值分別僅為對照的73.4%、80.2%。
3 討論
3.1 鎂缺乏和過量脅迫對紐荷爾臍橙不同葉齡葉片光合色素合成的影響
鎂是構(gòu)成葉綠素的中心元素,其含量與相對葉綠素呈極顯著相關(guān)[19]。Schimansky用放射性同位素28Mg對菜豆、大麥進(jìn)行示蹤,證明鎂可以通過韌皮部由老葉轉(zhuǎn)移至新葉[20]。對大豆[21]和黃瓜[8]的研究表明,葉片葉綠素含量隨根系供鎂含量的增加呈先上升后下降的趨勢;雖然缺鎂黃瓜葉片葉綠素a、葉綠素b、胡蘿卜素含量均極顯著降低,但是其葉綠素a/b值最高。Hermans等的研究表明缺鎂脅迫引起葉綠體結(jié)構(gòu)與功能異常、葉綠素含量下降、葉片異常黃化,進(jìn)而直接影響PSⅠ和PSⅡ的功能[10]。申燕等對“春見”橘橙的研究表明,缺鎂脅迫導(dǎo)致葉綠體內(nèi)類囊體片層模糊,片層數(shù)明顯減少,線粒體膜模糊且出現(xiàn)異常的黑色顆粒,葉綠素含量降低;鎂過量脅迫會導(dǎo)致淀粉粒和質(zhì)體小球異常增多增大,線粒體膜模糊,葉綠體內(nèi)片層消失,葉綠素含量顯著降低[22]。本文研究表明,缺鎂不僅會導(dǎo)致老葉(2齡秋梢葉)葉綠素含量降低,也會導(dǎo)致新葉(1齡春梢葉、1齡夏梢葉和1齡晚夏梢葉)葉綠素含量降低;雖然短期(2個月)缺鎂對2齡秋梢葉和1齡春梢葉葉綠素的合成影響不顯著,但是隨著缺鎂脅迫時間的增加植株葉綠素合成受抑制程度顯著增大。相對于鎂缺乏脅迫來說,鎂過量脅迫對減緩老葉和1齡春梢葉的葉綠素含量下降具有顯著效果,而其對新葉(1齡夏梢葉和1齡晚夏梢葉)葉綠素合成的影響則相反,其機(jī)制是元素拮抗還是其他原因有待進(jìn)一步的探討。
3.2 鎂缺乏和過量脅迫對紐荷爾臍橙不同葉齡葉片光合作用和早衰的影響
鎂脅迫不僅影響植株的葉綠素合成,而且也會影響植株的光合作用。缺鎂脅迫越嚴(yán)重,植物的光合作用效率就越低。現(xiàn)有的研究表明,缺鎂會引起光合膜垛疊受阻,降低PSⅠ與PSⅡ相對熒光產(chǎn)量的比值[23-24];缺鎂黃瓜葉片凈光合速率(Pn)、氣孔導(dǎo)度(Gs)和蒸騰速率(Tr)的日變化呈單峰曲線,易發(fā)生顯著的光抑制,葉綠素?zé)晒鈪?shù)Fv/Fm、Fv/Fo、qP和qN降低[7];缺鎂龍眼光補(bǔ)償點(diǎn)和CO2補(bǔ)償點(diǎn)提高,光飽和點(diǎn)和CO2飽和點(diǎn)下降,PSⅡ活性下降[6]。
圖4 鎂脅迫對紐荷爾臍橙不同葉齡葉片的非光化學(xué)淬滅(NPQ)的影響Fig.4 The effect of Mg stress on non-photoche-mical quenching(NPQ)in leaves at different foliar age of Newhall navel orange
本研究表明,隨著缺鎂時間的延長,紐荷爾臍橙新葉和老葉Fv/Fm受脅迫程度呈顯著增大趨勢;老葉Fv/Fm被抑制的表現(xiàn)時間顯著早于新葉,而且其Fv/Fm降低程度顯著大于新葉。此外,PSⅡ光能活化、活性氧代謝與植物葉片的早衰有密切的聯(lián)系[11,25]。在水稻葉片的早衰研究中發(fā)現(xiàn),在衰老過程中劍葉的Fv/Fm、qP、ФPSⅡ、rETR逐漸下降,qN上升,過剩的光能導(dǎo)致活性氧、H2O2和膜脂過氧化產(chǎn)物MDA的積累,導(dǎo)致葉綠素衰減,發(fā)生光氧化早衰[26]。在夏季強(qiáng)光照條件下葉片吸收的光能如不能完全通過光化學(xué)、非光化學(xué)和光子躍遷淬滅,過量的光能將會對光系統(tǒng)Ⅱ產(chǎn)生傷害,導(dǎo)致葉綠素含量降低,顯現(xiàn)黃化早衰癥狀[11]。缺鎂至第3月時紐荷爾臍橙新葉和老葉rETR呈顯著下降趨勢,第4月時2齡秋梢、1齡春梢葉和1齡夏梢葉的NPQ不增反降,熱耗散能力顯著降低。表明,在夏季高光條件下缺鎂植株吸收的過量光能很難以熱的形式耗散掉,易受到光抑制,產(chǎn)生光傷害;結(jié)合2齡秋梢和1齡春梢葉Fv/Fm的顯著降低結(jié)果可知,夏季高光已導(dǎo)致缺鎂紐荷爾臍橙葉片受到光傷害。
鎂缺乏和過量脅迫均能使煙草葉片量子產(chǎn)量、光合電子傳遞速率、非光化學(xué)猝滅降低,使植株對強(qiáng)光的保護(hù)性調(diào)節(jié)能力下降。鎂過量脅迫下黃瓜Pn和Tr呈雙峰曲線,胞間CO2濃度(Ci)、Gs呈近似倒拋物線型,光飽和點(diǎn)、CO2飽和點(diǎn)以及表觀量子效率和羧化效率都大于缺鎂脅迫[13]。本研究表明,與鎂缺乏相比,鎂過量對紐荷爾臍橙Fv/Fm的脅迫影響顯著較低,這與植株rETR和NPQ的光響應(yīng)能力變化有關(guān)。短期(2個月)鎂過量脅迫對紐荷爾臍橙2齡秋梢和1齡春梢葉rETR和NPQ的光響應(yīng)能力無顯著影響。雖然隨著脅迫時間的增加,新葉(1齡春梢葉、1齡夏梢葉和1齡晚夏梢葉)的rETR均呈顯著降低趨勢,但是其NPQ均高于對照,熱耗散能力較高,不易受到過量光能的抑制,產(chǎn)生光傷害。對于紐荷爾臍橙植株老葉來說,雖然自第3個月開始NPQ低于對照,熱耗散能力有所下降,但是其rETR與對照無顯著差異,也不易受到過量光能的抑制,不易產(chǎn)生光傷害。
綜上所述,缺鎂脅迫加速2齡秋稍葉和1齡春梢葉的衰老,而鎂過量脅迫延緩其衰老;夏季高光易導(dǎo)致缺鎂脅迫臍橙葉片受到光傷害,而對鎂過量脅迫臍橙葉片的影響較小,不易產(chǎn)生光傷害。因此,生產(chǎn)上應(yīng)重視鎂肥的施用。由本研究結(jié)果可知,缺鎂脅迫對紐荷爾臍橙植株的不利影響顯著大于鎂過量脅迫;而且生產(chǎn)上一般的鎂肥過量應(yīng)不會對樹體產(chǎn)生顯著的不利影響。
[1] Peng L Z,Lai J J,Chun C P,Cao L,Zhong L H,Jiang C L,Ling L L.Role of compound magnesium fertilizer in correcting leaf yellowing caused by nutrient element deficiency on navel orange in Gannan.South China Fruits,2008,37(3):10-12.
[2] Li J,Xie Z C,Xie W L,Wu X M,Shi Q.Relationship between leafveinsplittingandmineralnutritionofcitrus.Acta Horticulturae Sinica,2011,38(3):425-433.
[3] Wu W H,Peters J,Berkowitz G A.Surface charge mediated effects of Mg2+on K+flux across the chloroplast envelope are associated with regulation of stromal pH and photosynthesis.Plant Physiology,1991,97(2):580-587.
[4] Hao N P,Li T Z,Zhang Q D,Lou S Q,Tang C Q,Kuang T Y.Structure and function of chloroplast membranesⅧ.Effect of magnesium ions on the polymerization of chlorophyll-protein complexesofchloroplastmembranes.ActaBiochimicaet Biophysica Sinica,1981,13(4):365-372.
[5] Sun N,Zeng X B,Gao J S,Wang B R.Effects of magnesium compound fertilizer on daylily(Hemerocallis citrina Baroni)growth and soil Nutrients.Scientia Agricultura Sinica,2006,39(1):95-101.
[6] Li Y,Liu X H,Zhuang W M.The effect of magnesium deficiency on photosynthesis of Longan(Dimocarpus long ana Lour.)seedlings.Acta Horticulturae Sinica,2001,28(2):101-106.
[7] Yang G D,Zhu Z J,Ji Y M.Effect of light intensity and magnesium deficiency on chlorophyll fluorescence and active oxygen in cucumber leaves.Plant Nutrition and Fertilizer Science,2002,8(1):115-118.
[8] Xie X Y,Liu H T,Cheng Z W.Effects of magnesium stress on photosynthetic characterofcucumberingreenhouse.China Vegetables,2009,(6):36-40.
[9] Liu H C,Chen X M,Chen R Y,Song S W,Sun G W.Effects of magnesium deficiency on photosynthesis characteristic of flowering Chinese cabbage.Horticulturae Sinica,2006,33(2):311-316.
[10] Hermans C,Johnson G N,Strasser R J,Verbruggen N.Physiological characterisation of magnesium deficiency in sugar beet:Acclimationtolowmagnesiumdifferentiallyaffects photosystemsⅠandⅡ.Planta,2004,220(2):344-355.
[11] Yang Y,Jiang D A,Sun J W,Huang Z A,Jin S H.Effects of different magnesium nutrition levels on chlorophyll fluorescence characteristics and excitation energy dissipation in rice leaves.Plant Nutrition and Fertilizer Science,2005,11(1):79-86.
[12] Hermans C,Verbruggen N.Physiological characterization of Mg deficiency inArabidopsisthaliana.JournalofExperimental Botany,2005,56(418):2153-2161.
[13] Guan G S,Tu N M,Xiao H Q,Zhu L S,Zhu Y H,Wang Z M.Effectsofmagnesiumontobaccogrowthandchlorophyll fluorescence parameters of tobacco leaves.Plant Nutrition and Fertilizer Science,2008,14(1):151-155.
[14] Scheriber U,Gademann R,Ralph P J,Larkum A W D.AssessmentofphotosyntheticperformanceofProchloronin Lissoclinumpatellainhospitebychlorophyllfluorescence measurements.Plant and Cell Physiology,1997,38(8):945-951.
[15] Platt T,Gallegos C L,Harrison W G.Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton.Journal of Marine Research,1980,38(4):687-701.
[16] Demmig-Adams B,Adams W WⅢ,Barker D H,Logan B A,Bowling D R,Verhoeven A S.Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation.Physiologia Plantarum,1996,98(2):253-264.
[17] Genty B,Briantais J M,Baker N R.The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence.Biochimica et Biophysica Acta,1989,990(1):87-92.
[18] Horton P,Ruban A V,Walters R G.Regulation of light harvesting in green plants(indicationby nonphotochemical quenching of chlorophyll fluorescence).Plant Physiology,1994,106(2):415-420.
[19] Zhang G Y,Peng L Z,Chun C P,Zeng M,Ling L L,Lai J J,Wang Z X.Seasonal changes in leaf magnesium and boron contents and their relationships to leaf yellowing of navel orange(Citrus sinensis Osbeck).Acta Horticulturae Sinica,2010,37(8):1317-1324.
[20] Schimansky C.Possibilities and limitations of utilizing28Mg for studying Mg-specific problems in higher plants.Journal of Plant Nutrition,1985,8(6):467-489.
[21] Li Y Z,Li F,Gao M,Chen X,Lv C L,F(xiàn)u H L,Sun D Q.Effects of soil magnesium excess on several physiological indexes of soybean.Agro-Environmental Protection,1990,9(2):41-42.
[22] Shen Y,Xiao J X,Yang H,Zhang S L.Effects of magnesium stress on growth,distribution of several mineral elements and leaf ultrastructure of‘Harumi’tangor.Acta Horticulturae Sinica,2011,38(5):849-858.
[23] Zhang Q D,Tang C Q,Li S Y.Structure and function of chloroplast membraneⅪ.The effects of linolenic acid on the structure and the absorption and fluorescence spectra of wheat chloroplast membranes as well as regulations by MgCl2.Acta Botanica Sinica,1982,24(4):326-333.
[24] Lin S Q,Xu C H,Zhang Q D,Xu L,Mao D Z,Kuang T Y.Some application of chlorophyll fluorescence kinetics to plant stress physiology to ecology and agricultural modernization.Chinese Bulletin of Botany,1992,9(1):1-16.
[25] Li Y,Liu X H.Effects of magnesium deficiency on senescence of Dimocarpus longana leaves.Chinese Journal of Applied Ecology,2002,13(3):311-314.
[26] Jiao D M,Li X,Huang X Q,Ji B H.The relationship among photoinhibition,Photooxidationandearlyagingatlater developmental stages in different high yield varieties.Scientia Agricultura Sinica,2002,35(5):487-492.
參考文獻(xiàn):
[1] 彭良志,賴九江,淳長品,曹立,鐘莉華,江才倫,凌麗俐.復(fù)合鎂肥對贛南臍橙缺素黃化的矯治作用.中國南方果樹,2008,37(3):10-12.
[2] 李健,謝鐘琛,謝文龍,吳應(yīng)明,施清.柑橘葉脈開裂癥與礦質(zhì)營養(yǎng)的關(guān)系.園藝學(xué)報,2011,38(3):425-433.
[4] 郝廼斌,李桐柱,張其德,婁世慶,唐崇欽,匡廷云.葉綠體膜的結(jié)構(gòu)與功能Ⅷ.鎂離子對葉綠體類囊體膜的葉綠素-蛋白復(fù)合體聚合的影響.生物化學(xué)與生物物理學(xué)報,1981,13(4):365-372.
[5] 孫楠,曾希柏,高菊生,王伯仁.含鎂復(fù)合肥對黃花菜生長及土壤養(yǎng)分含量的影響.中國農(nóng)業(yè)科學(xué),2006,39(1):95-101.
[6] 李延,劉星輝,莊衛(wèi)民.缺鎂對龍眼光合作用的影響.園藝學(xué)報,2001,28(2):101-106.
[7] 楊廣東,朱祝軍,計玉妹.不同光強(qiáng)和缺鎂脅迫對黃瓜葉片葉綠素?zé)晒馓匦院突钚匝醍a(chǎn)生的影響.植物營養(yǎng)與肥料學(xué)報,2002,8(1):115-118.
[8] 謝小玉,劉海濤,程志偉.鎂對溫室黃瓜光合特性的影響.中國蔬菜,2009,(6):36-40.
[9] 劉厚誠,陳細(xì)明,陳日遠(yuǎn),宋世威,孫光聞.缺鎂對菜薹光合作用特性的影響.園藝學(xué)報,2006,33(2):311-316.
[11] 楊勇,蔣德安,孫駿威,黃宗安,金松恒.不同供鎂水平對水稻葉片葉綠素?zé)晒馓匦院湍芰亢纳⒌挠绊?植物營養(yǎng)與肥料學(xué)報,2005,11(1):79-86.
[13] 關(guān)廣晟,屠乃美,肖漢乾,朱列書,朱英華,王中美.鎂對煙草生長及葉片葉綠素?zé)晒鈪?shù)的影響.植物營養(yǎng)與肥料學(xué)報,2008,14(1):151-155.
[19] 張廣越,彭良志,淳長品,曾明,凌麗俐,賴九江,王振興.臍橙葉片鎂、硼含量變化與缺素黃化的關(guān)系.園藝學(xué)報,2010,37(8):1317-1324.
[21] 李亞洲,李沸,高銘,陳旭,呂春莉,付海龍,孫大群.土壤中過量鎂對大豆幾項生理指標(biāo)的影響.農(nóng)業(yè)環(huán)境保護(hù),1990,9(2):41-42.
[22] 申燕,肖家欣,楊慧,張紹鈴.鎂脅迫對‘春見’橘橙生長和礦質(zhì)元素分布及葉片超微結(jié)構(gòu)的影響.園藝學(xué)報,2011,38(5):849-858.
[23] 張其德,唐崇欽,李世儀.葉綠體膜的結(jié)構(gòu)與功能Ⅺ.亞麻酸對小麥葉綠體膜結(jié)構(gòu)、吸收光譜和熒光光譜的影響以及鎂離子的調(diào)節(jié)作用.植物學(xué)報,1982,24(4):326-333.
[24] 林世青,許春輝,張其德,徐黎,毛大璋,匡廷云.葉綠素?zé)晒鈩恿W(xué)在植物抗性生理學(xué)、生態(tài)學(xué)和農(nóng)業(yè)現(xiàn)代化中的應(yīng)用.植物學(xué)通報,1992,9(1):1-16.
[25] 李延,劉星輝.缺鎂脅迫對龍眼葉片衰老的影響.應(yīng)用生態(tài)學(xué)報,2002,13(3):311-314.
[26] 焦德茂,李霞,黃雪清,季本華.不同高產(chǎn)水稻品種生育后期葉片光抑制、光氧化和早衰的關(guān)系.中國農(nóng)業(yè)科學(xué),2002,35(5):487-492.
猜你喜歡
中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識經(jīng)濟(jì)·中國直銷(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級版)(2011年3期)2011-01-01 00:00:00
