洞庭湖浮游植物增長(zhǎng)的限制性營(yíng)養(yǎng)元素研究
2014-07-16 05:40:16李利強(qiáng)黃代中熊劍張屹田琪何英余建清王琦
生態(tài)環(huán)境學(xué)報(bào) 2014年2期
關(guān)鍵詞:營(yíng)養(yǎng)實(shí)驗(yàn)
李利強(qiáng),黃代中*,熊劍,張屹,田琪,何英,余建清,王琦
1. 湖南省洞庭湖生態(tài)環(huán)境監(jiān)測(cè)中心,湖南 岳陽(yáng) 414000;2. 岳陽(yáng)市環(huán)境監(jiān)測(cè)中心,湖南 岳陽(yáng) 414000
洞庭湖為湖南省第一大湖,全國(guó)第二大淡水湖,是承納湘、資、沅、澧四水和吞吐長(zhǎng)江的過(guò)水性洪道型湖泊,有溝通航運(yùn)、繁衍水產(chǎn)、調(diào)蓄長(zhǎng)江和改善生態(tài)環(huán)境等多種功能。洞庭湖區(qū)是湖南省主要造紙、石化輕工及紡織工業(yè)基地,部分在全國(guó)舉足輕重,每天都有大量廢水產(chǎn)生(黃金國(guó),2003)。相關(guān)研究表明,洞庭湖的 TN、TP、懸浮物和總大腸菌群是洞庭湖的主要污染因子,其中 TN和 TP尤為突出(米紅州等,2004;DU等,2001;徐開(kāi)欽等,2004)。洞庭湖水體N、P污染加劇,富營(yíng)養(yǎng)化日趨嚴(yán)重,致使洞庭湖生態(tài)系統(tǒng)中生物多樣性降低。據(jù)水產(chǎn)部門(mén)觀測(cè)調(diào)查,近 10年來(lái),短鱭魚(yú)、銅魚(yú)、黃尾魚(yú)、針魚(yú)、鱘魚(yú)、白鱘魚(yú)等 15個(gè)品種己經(jīng)少見(jiàn)或基本滅絕;銀魚(yú)、黃魚(yú)產(chǎn)量明顯減少;其次是常規(guī)魚(yú)類(lèi)品種退化(莊大昌等,2003)。
針對(duì)富營(yíng)養(yǎng)化發(fā)生過(guò)程與機(jī)制,國(guó)內(nèi)外已有一些研究報(bào)道,但是機(jī)理目前尚未完全明了(秦伯強(qiáng),2002;李文朝,1997;Philips等,1999)。因地理?xiàng)l件、點(diǎn)源面源營(yíng)養(yǎng)輸入、沉積物性質(zhì)、微生物群落結(jié)構(gòu)及藻類(lèi)群落組成等因素影響,各湖泊浮游植物生長(zhǎng)繁殖的限制性營(yíng)養(yǎng)元素及形態(tài)差異較大(Quiblier等,2008;Becker等,2010)。藻類(lèi)增長(zhǎng)的生物學(xué)實(shí)驗(yàn)(Nutrient Enrichment Bioassay,NEB)是一種有效檢測(cè)浮游植物營(yíng)養(yǎng)鹽限制的主要研究方法,能找出水體中浮游植物增長(zhǎng)限制性營(yíng)養(yǎng)元素,可以有效地進(jìn)行預(yù)測(cè)、控制藻類(lèi)增殖,該方法已經(jīng)廣泛地應(yīng)用于湖泊與海洋的限制營(yíng)養(yǎng)因子研究(吳雪峰等,2010;張亞克等,2011;Elser和Kimmel,1986)。基于室內(nèi)營(yíng)養(yǎng)添加實(shí)驗(yàn)的結(jié)果發(fā)現(xiàn),N、P都可以成為淡水藻類(lèi)生長(zhǎng)的限制因子(Henry等,1984;Zhou等,2009)。同一湖泊不同季節(jié)時(shí),藻類(lèi)生長(zhǎng)的限制因子也可能不同。在冬春季,太湖浮游植物生物量和生長(zhǎng)速率隨磷增加而顯著增加,與氮無(wú)關(guān),表明浮游植物生長(zhǎng)的磷限制;但是,在夏秋季水華期,氮是主要限制營(yíng)養(yǎng)因子(Xu等,2010)。洞庭湖氮、磷污染嚴(yán)重,入湖 TN、TP 總量分別為 59049 t·a-1和 6913 t·a-1(秦迪嵐等,2011)。水體氮、磷污染勢(shì)必影響洞庭湖浮游植物生物量及群落結(jié)構(gòu)。營(yíng)養(yǎng)鹽賦存形態(tài)及其濃度與浮游植物生物量增長(zhǎng)的偶聯(lián)關(guān)系如何等科學(xué)問(wèn)題在洞庭湖少有研究。本項(xiàng)目在了解洞庭湖浮游植物生物量及主要營(yíng)養(yǎng)鹽形態(tài)組成與其含量的年變化基礎(chǔ)上,通過(guò) NEB實(shí)驗(yàn)確定洞庭湖浮游植物增長(zhǎng)的限制性營(yíng)養(yǎng)元素,可為洞庭湖水污染治理及富營(yíng)養(yǎng)化防治提供理論依據(jù)。
1 材料與方法
1.1 水樣采集
洞庭湖常規(guī)水質(zhì)監(jiān)測(cè)斷面分布見(jiàn)圖1。每個(gè)監(jiān)測(cè)斷面設(shè)左、中、右3條垂線,分別采表層(0.5 m)水樣。水質(zhì)采樣于月初進(jìn)行。
NEB實(shí)驗(yàn)于2011年9月和2012年5月在東洞庭湖采集表層水80 L作為實(shí)驗(yàn)水樣。每次采集水樣后測(cè)定生物化學(xué)指標(biāo),具體環(huán)境特征參數(shù)見(jiàn)表 1。采集實(shí)驗(yàn)水樣時(shí)現(xiàn)場(chǎng)測(cè)定水體透明度、水溫和 pH等指標(biāo),實(shí)驗(yàn)室分析TN、TP、氨態(tài)氮(NH4+-N)、硝態(tài)氮(NO3--N)、SRP、溶解氧(DO)和葉綠素a(Chla)等指標(biāo),以確定實(shí)驗(yàn)水樣初始營(yíng)養(yǎng)物濃度和浮游植物初級(jí)生產(chǎn)力水平。
1.2 研究方法
NEB實(shí)驗(yàn)不需對(duì)培養(yǎng)水樣進(jìn)行滅菌、過(guò)濾和添加藻種等處理,而是直接利用水體中原始的生態(tài)群落進(jìn)行培養(yǎng),可實(shí)際反映不同時(shí)期生物群落的水體添加營(yíng)養(yǎng)鹽后對(duì)浮游植物增長(zhǎng)的影響。經(jīng)形態(tài)鑒定,2011年9月和2012年5月東洞庭湖原水樣中浮游植物均以硅藻為主,優(yōu)勢(shì)種為針桿藻(Synedrasp.)、菱形藻(Nitzschiasp.)和小環(huán)藻(Cyclotellasp.)。直接量取1 L水樣分裝到1 L錐形瓶中在實(shí)驗(yàn)條件下培養(yǎng),每個(gè)營(yíng)養(yǎng)鹽水平做6個(gè)平行樣,培養(yǎng)10 d后,一部分錐形瓶水樣取樣測(cè)定各指標(biāo),另一部分培養(yǎng)20 d后測(cè)定水樣中各指標(biāo)。實(shí)驗(yàn)條件為溫度25 ℃;光照4000 lx;光暗時(shí)間比12:12。以原湖水TN和TP作為初始營(yíng)養(yǎng)物水平,以初始營(yíng)養(yǎng)物水平的0.5倍、1.0倍和2.0倍添加營(yíng)養(yǎng)鹽,添加方案見(jiàn)表2(張亞克等,2011)。添加方式為一次性添加,以磷酸二氫鉀(KH2PO4)、硝酸鉀(KNO3)和氯化銨(NH4Cl)為添加營(yíng)養(yǎng)鹽。
1.3 測(cè)定方法
TN、TP、Chla、NH4+-N、NO3--N、亞硝態(tài)氮(NO2--N)等監(jiān)測(cè)項(xiàng)目均根據(jù)《水和廢水監(jiān)測(cè)分析方法》(國(guó)家環(huán)境保護(hù)總局,2002)推薦的方法進(jìn)行分析。DIN為NH4+-N、NO3--N和NO2--N之和。溶解態(tài)總磷(DTP)用過(guò)0.45 μm濾膜后的水樣測(cè)定,實(shí)驗(yàn)步驟與TP一致。SRP用鉬藍(lán)比色法測(cè)定(MURPHY和RILEY,1962)。顆粒態(tài)磷(PP)為T(mén)P與DTP的差值,溶解態(tài)有機(jī)磷(DOP)為DTP與SRP的差值。
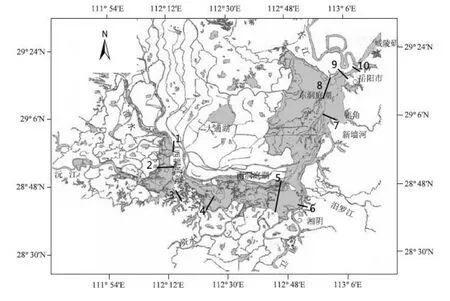
圖1 洞庭湖水質(zhì)監(jiān)測(cè)斷面分布Fig.1 Sampling sections of water quality monitoring in Dongting Lake
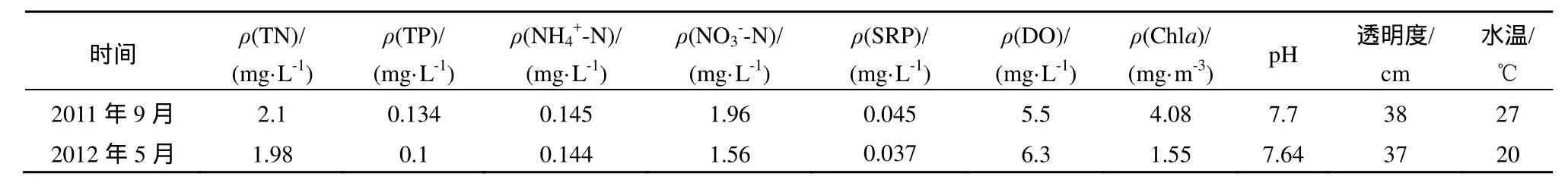
表1 東洞庭湖水體環(huán)境特征參數(shù)Table 1 Environmental parameters in water phase of Dongting Lake
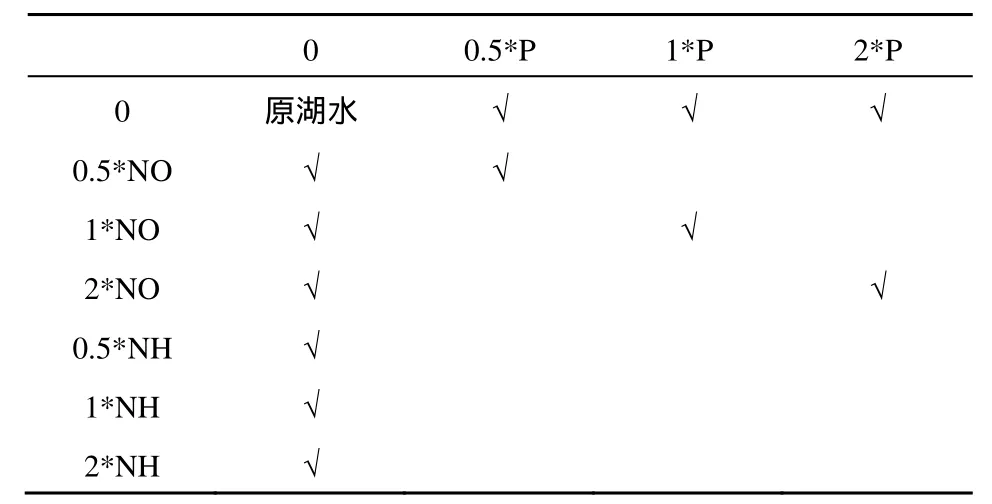
表2 浮游植物增長(zhǎng)實(shí)驗(yàn)營(yíng)養(yǎng)鹽添加方案Table 2 Nutrient addition methods of phytoplankton growth experiment
1.4 數(shù)據(jù)處理
為了能夠消除不同時(shí)期原湖水帶來(lái)的差異(吳雪峰等,2010;張亞克等,2011),采用相對(duì)比較系數(shù)法表示添加營(yíng)養(yǎng)物對(duì)浮游植物生長(zhǎng)的影響:

式中:DT為某營(yíng)養(yǎng)物添加組與對(duì)照組的比較系數(shù);AT為某營(yíng)養(yǎng)添加組ρ(Chla);AC為對(duì)照組ρ(Chla)。
采用SigmaPlot 10.0軟件作圖,采用SPSS 13.0軟件進(jìn)行數(shù)據(jù)統(tǒng)計(jì)。相關(guān)性用Pearson相關(guān)系數(shù)表示,營(yíng)養(yǎng)添加組與對(duì)照組之間的顯著性差異性比較用單因素方差分析(LSD)。
2 結(jié)果與分析
2.1 水體營(yíng)養(yǎng)鹽及Chl a逐月分布特征
洞庭湖水體中ρ(TN)較高,月平均值變化范圍為 1.53~2.21 mg·L-1,1-4 月ρ(TN)高于其它月份,均超過(guò) 2 mg·L-1(圖 2)。ρ(NO3--N)年平均值為 1.38 mg·L-1,月平均值介于 1.17~1.60 mg·L-1之間;ρ(NH4+-N)年平均值為 0.23 mg·L-1,月平均值介于0.06~0.56 mg·L-1之間;ρ(NO2--N)年平均值為 0.039 mg·L-1,月平均值介于 0.016~0.058 mg·L-1之間。洞庭湖水體中 TN 以溶解態(tài)無(wú)機(jī)氮為主,ρ(DIN)占ρ(TN)比例在 71.3%~98.7%之間,且溶解態(tài)無(wú)機(jī)氮中ρ(NO3--N)>ρ(NH4+-N)>ρ(NO2--N)。ρ(NO3--N)在當(dāng)年11月至次年3月較高,ρ(NH4+-N)在4月出現(xiàn)最高峰,兩者均在夏季含量相對(duì)較低。
ρ(TP)年平均值為 0.093 mg·L-1,月平均值變化范圍為 0.054~0.129 mg·L-1,ρ(TP)于 3-4 月份相對(duì)較高,8月份最低(圖3)。洞庭湖水體中磷以溶解態(tài)磷為主,ρ(DTP)占ρ(TP)比例范圍為60%~85%。磷形態(tài)組成中,ρ(SRP)>ρ(PP)>ρ(DOP)。ρ(SRP)月變化趨勢(shì)與ρ(TP)基本一致,均在春季相對(duì)較高。
洞庭湖水體中ρ(Chla)較低,年平均值僅 2.49 mg·m-3,月均值介于 1.43~4.11 mg·m-3之間。ρ(Chla)于7月和9月較高,春季相對(duì)較低(圖4)。
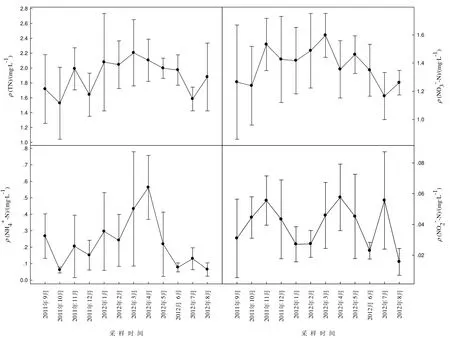
圖2 洞庭湖水體中氮含量月變化Fig.2 Monthly variations of nitrogen concentrations in water phase of Dongting Lake
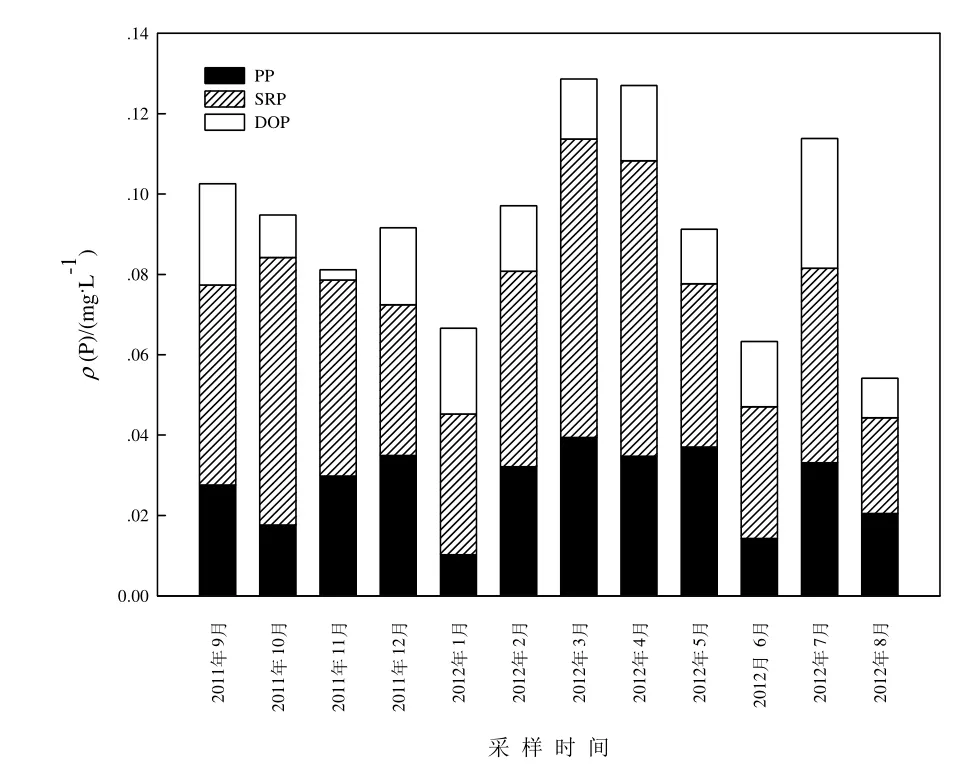
圖3 洞庭湖水體中磷含量月變化Fig.3 Monthly variations of phosphorus concentrations in water phase of Dongting Lake
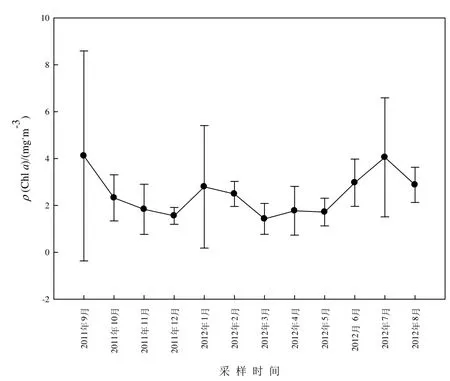
圖4 洞庭湖水體中Chla月變化Fig.4 Monthly variations of chlorophyll a (Chla) concentration in water phase of Dongting Lake
2.2 添加不同營(yíng)養(yǎng)鹽對(duì)洞庭湖浮游植物生物量的影響
為能消除不同時(shí)期原湖水帶來(lái)的差異,采用相對(duì)比較系數(shù)方法表示添加不同營(yíng)養(yǎng)元素浮游植物生物量與對(duì)照的差值,其中對(duì)照組結(jié)果均表達(dá)為0%(張亞克等,2011),NEB實(shí)驗(yàn)結(jié)果如圖5-6所示。2011年9月,添加營(yíng)養(yǎng)鹽后浮游植物生物量較對(duì)照有不同程度的增加(除0.5*P外),實(shí)驗(yàn)后期浮游植物生物量高于實(shí)驗(yàn)前期,其中,添加2*NH實(shí)驗(yàn)后期相對(duì)比較系數(shù)最高,達(dá)677%。2012年5月,營(yíng)養(yǎng)鹽添加后浮游植物生物量相對(duì)比較系數(shù)低于2011年9月,且大多數(shù)處理中實(shí)驗(yàn)前期浮游植物生物量高于實(shí)驗(yàn)后期,添加2*NH實(shí)驗(yàn)前期相對(duì)比較系數(shù)最高,達(dá)385%。
3 討論
3.1 營(yíng)養(yǎng)鹽來(lái)源
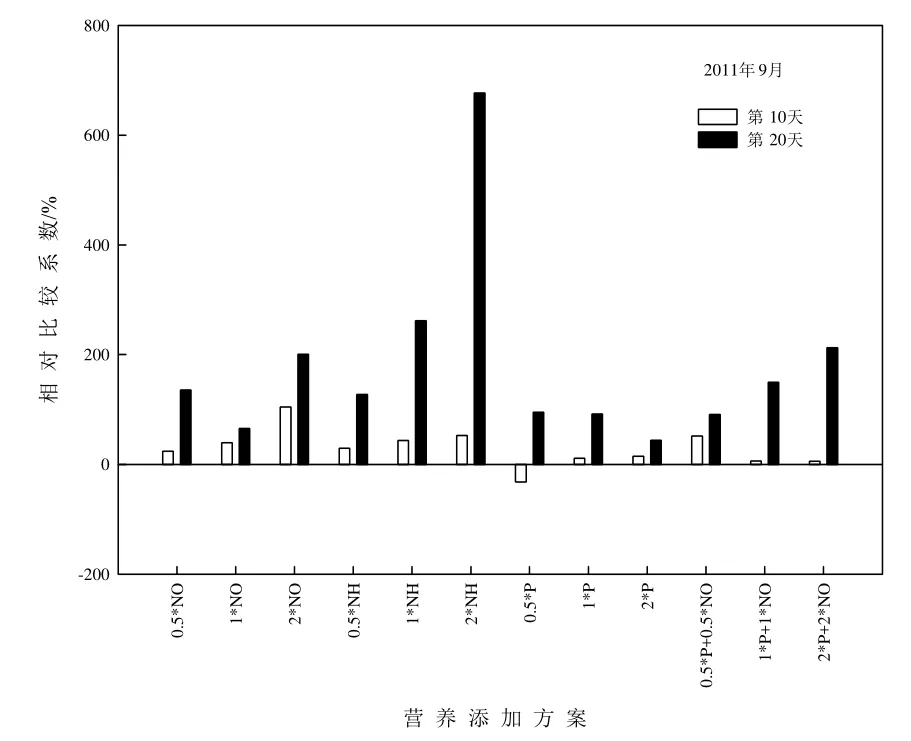
圖5 2011年9月NEB實(shí)驗(yàn)分析Fig.5 Analysis of nutrient enrichment bioassay (NEB)experiment in September, 2011
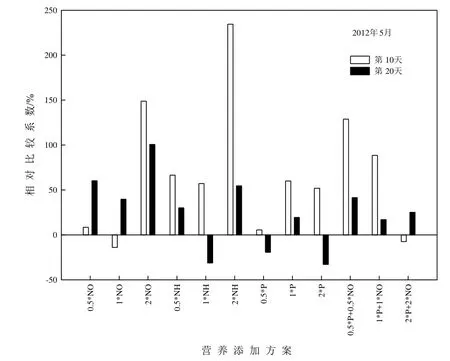
圖6 2012年5月NEB實(shí)驗(yàn)分析Fig. 6 Analysis of nutrient enrichment bioassay (NEB)experiment in May, 2012
TN、TP是影響洞庭湖水體富營(yíng)養(yǎng)化的主要營(yíng)養(yǎng)指標(biāo),二者一直維持較高的水平,20世紀(jì)90年代中期后全湖TN、TP超標(biāo),成為洞庭湖水質(zhì)惡化和水體營(yíng)養(yǎng)化程度加劇的重要因子(黃代中等,2013)。ρ(TN)波動(dòng)較大,存在明顯的區(qū)域性,而且自1998年以來(lái),一直在1.50 mg·L-1上下波動(dòng)。ρ(TP)在洞庭湖中分布較均勻,無(wú)明顯的區(qū)域性(楊漢等,1999)。春季洞庭湖水體中ρ(TN)、ρ(TP)較高,可能源于春季面源污染。洞庭湖水體中的 TN和 TP主要來(lái)源于畜禽養(yǎng)殖、農(nóng)田徑流和城鎮(zhèn)生活污染(秦迪嵐等,2011)。據(jù)監(jiān)測(cè),水體中溶解態(tài)無(wú)機(jī)氮平均占ρ(TN)的比例為 87%,溶解態(tài)總磷平均占ρ(TP)的比例為 70%,且大多數(shù)氮以硝態(tài)氮形式存在,大多數(shù)磷以磷酸鹽為主,這說(shuō)明氮、磷的來(lái)源與化肥使用有密切聯(lián)系(朱廣偉,2009)。受流域社會(huì)經(jīng)濟(jì)發(fā)展等因素的綜合影響,洞庭湖整體水質(zhì)呈下降趨勢(shì),富營(yíng)養(yǎng)化日趨嚴(yán)重。
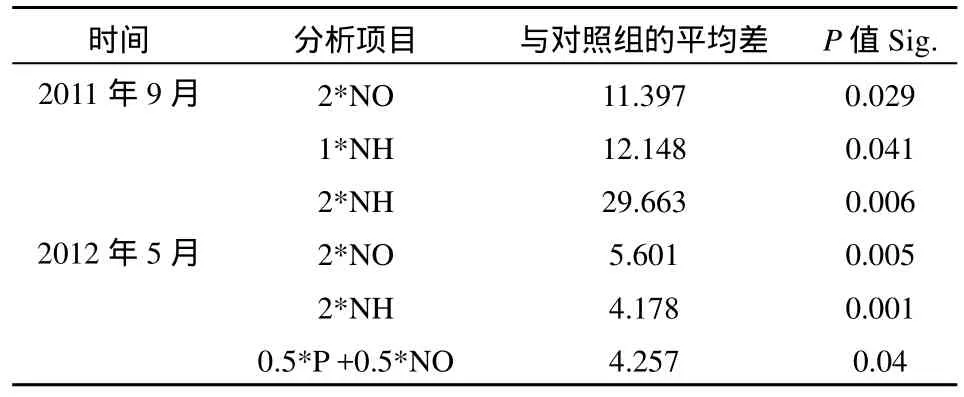
表3 NEB實(shí)驗(yàn)結(jié)果方差分析(平均ρ(Chla))Table 3 Analysis of variance for the nutrient enrichment bioassay (NEB) experiment: Chla
3.2 浮游植物增長(zhǎng)的營(yíng)養(yǎng)限制因子
2011年9月份單獨(dú)添加 NH4-N對(duì)浮游植物生長(zhǎng)具有明顯的促進(jìn)作用,方差分析結(jié)果顯示添加1*NH和2*NH均與對(duì)照有顯著性差異(P<0.05,表3);單獨(dú)添加NO3--N和單獨(dú)添加P對(duì)浮游植物生長(zhǎng)有一定的促進(jìn)作用,單獨(dú)添加P有時(shí)出現(xiàn)抑制作用;添加P和NO3-N與單獨(dú)添加NO3-N結(jié)果相當(dāng),P和NO3-N不存在交互作用。2012年5月份高氮組平均ρ(Chla)與對(duì)照有顯著性差異(P<0.05,表3)。兩次 NEB實(shí)驗(yàn)表明氮對(duì)洞庭湖浮游植物生長(zhǎng)有明顯的促進(jìn)作用,且促進(jìn)作用隨氮濃度的增加而加強(qiáng),氮是洞庭湖浮游植物增長(zhǎng)的主要營(yíng)養(yǎng)限制性因子,而磷對(duì)浮游植物生長(zhǎng)影響不大,有時(shí)出現(xiàn)抑制作用,硝態(tài)氮與磷之間不存在交互作用。洞庭湖水體中浮游植物ρ(Chla)與營(yíng)養(yǎng)鹽的相關(guān)性結(jié)果見(jiàn)表 4,2011 年 9 月ρ(Chla)與ρ(NH4+-N)顯著正相關(guān),2012年5月ρ(Chla)與ρ(TN)、ρ(NO3--N)和ρ(NO2--N)顯著正相關(guān),兩次采樣水體中ρ(Chla)均與ρ(TP)顯著負(fù)相關(guān),這與 NEB實(shí)驗(yàn)結(jié)果一致。洞庭湖水體中TN以溶解態(tài)無(wú)機(jī)氮為主,溶解態(tài)營(yíng)養(yǎng)鹽更易被浮游植物吸收利用,浮游植物對(duì)氨態(tài)氮的利用優(yōu)于硝態(tài)氮和亞硝態(tài)氮。洞庭湖浮游植物量以硅藻為主,時(shí)空分布上均占優(yōu);其次為綠藻和藍(lán)藻,浮游植物生長(zhǎng)亦可能受一些微量元素影響,如Si、Fe、Mn等。
4 結(jié)論
1) 洞庭湖水體主要污染物為T(mén)N和TP,其月平均值變化范圍分別為 1.53~2.21 mg·L-1和0.054~0.129 mg·L-1。洞庭湖水體中TN以DIN為主,ρ(DIN)占ρ(TN)比例在 71.3%~98.7%之間,且 DIN中ρ(NO3--N)>ρ(NH4+-N)>ρ(NO2--N)。磷形態(tài)組成中,ρ(DTP)占ρ(TP)比例范圍為 60%~85%,ρ(SRP)>ρ(PP)>ρ(DOP)。大多數(shù)氮以硝態(tài)氮形式存在,大多數(shù)磷以磷酸鹽為主,而且春季洞庭湖水體中ρ(TN)、ρ(TP)更高,這說(shuō)明氮、磷的來(lái)源與化肥使用有密切聯(lián)系。
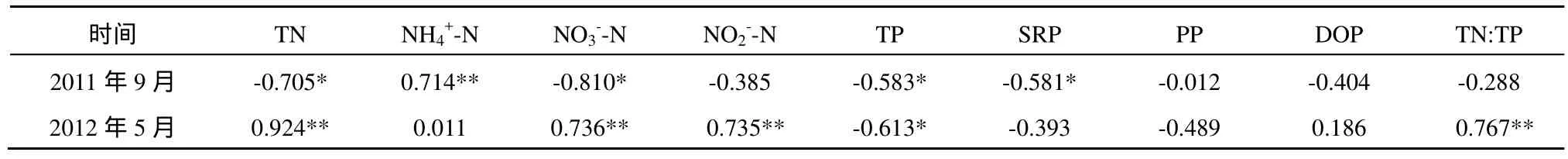
表4 洞庭湖水體中Chla與營(yíng)養(yǎng)鹽的相關(guān)性分析Table 4 Pearson correlation coefficients between Chlorophyll a concentration and nutrient content in water phase of Dongting Lake
2) 藻類(lèi)增長(zhǎng)生物學(xué)評(píng)價(jià)實(shí)驗(yàn)表明,氮是洞庭湖浮游植物增長(zhǎng)的主要營(yíng)養(yǎng)限制因子,添加氮(尤其是氨態(tài)氮)能明顯促進(jìn)浮游植物生物量增加,且促進(jìn)作用隨氮濃度的增加而加強(qiáng),添加磷對(duì)浮游植物生長(zhǎng)影響不大,有時(shí)出現(xiàn)抑制作用。洞庭湖水體中ρ(Chla)與氮顯著正相關(guān),與磷顯著負(fù)相關(guān)。這些研究結(jié)果表明洞庭湖富營(yíng)養(yǎng)化控制過(guò)程中應(yīng)特別注重氮的控制。
BECKER V, CAPUTO L, ORDONEZ J, et al. 2010.Driving factors of the phytoplankton functional groups in a deep Mediterranean reservoir[J].Water Research, 44(11): 3345-3354.
DU Y, CAI S M, ZHANG X Y, et al. 2001.Inteprretation of the environmental change of Dongting Lake, middle reach of Yangtze River, China, by210Pb measurement and satellite image analysis[J].Geomorphology, 41: 171-181.
ELSER J J, KIMMEL B L. 1986. Alteration of phytoplankton phosphorus status during enrichment experiments: implications for interpreting nutrient enrichment bioassay results[J]. Hydrobiologia, 133(3):217-222.
HENRY R, TUNDISI J G, CURI P R. 1984.Effects of phosphorus and nitrogen enrichment on the phytoplankton in a tropical reservoir (Lobo Reservoir, Brazil) [J]. Hydrobiologia, 118(2): 177-185.
MURPHY J, RILEY J P. 1962.A modified single solution method for determination of phosphorus in natural waters[J]. Analytic Chimica Acta, 26(1): 1-36.
PHILIPS G, BRAMWELL A, PITT J, et al. 1999.Practical application of 25 years’ research into the management of shallow lakes[J].Hydrobiologia, 136: 61-76.
QUIBLIER C, LEBOULANGER C, SANE S, et al. 2008.Phytoplankton growth control and risk of cyanobacterial blooms in the lower Senegal River delta region[J]. Water Research, 42(4/5): 1023-1034.
XU H, PAERL H W, QIN B Q, et al. 2010.Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu, China[J].Limnology and Oceanography, 55(1): 420-432.
ZHOU G J, BI Y H, ZHAO X M, et al. 2009.Algal growth potential and nutrient limitation in spring in Three-Gorges reservoir, China[J].Fresenius Environmental Bulletin, 18(9): 1642-1647.
國(guó)家環(huán)境保護(hù)總局. 2002. 水和廢水監(jiān)測(cè)分析方法[M]. 4版. 北京: 中國(guó)環(huán)境科學(xué)出版社.
黃代中, 萬(wàn)群, 李利強(qiáng), 等. 2013. 洞庭湖 20年水質(zhì)與富營(yíng)養(yǎng)化狀態(tài)變化[J]. 環(huán)境科學(xué)研究, 26(1): 27-33.
黃金國(guó). 2003.洞庭湖區(qū)濕地資源開(kāi)發(fā)中的生態(tài)環(huán)境問(wèn)題及對(duì)策[J]. 水土保持通報(bào), 23(l): 73-75.
李文朝. 1997.淺水湖泊生態(tài)系統(tǒng)的多穩(wěn)態(tài)理論及其應(yīng)用[J]. 湖泊科學(xué),9(2): 97-104.
米紅州, 莫多聞, 蘇成. 2004.洞庭湖演變趨勢(shì)探討[J]. 地理研究, 23(1):78-86.
秦伯強(qiáng). 2002.長(zhǎng)江中下游淺水湖泊富營(yíng)養(yǎng)化發(fā)生機(jī)制與控制途徑初探[J]. 湖泊科學(xué), 14(3): 193-202.
秦迪嵐, 黃哲, 羅岳平, 等. 2011.洞庭湖區(qū)污染控制區(qū)劃與控制對(duì)策[J].環(huán)境科學(xué)研究, 24(7): 748-755.
吳雪峰, 程曦, 李小平. 2010.淀山湖浮游植物營(yíng)養(yǎng)限制因子的研究[J].長(zhǎng)江流域資源與環(huán)境, 19(3): 292-298.
徐開(kāi)欽, 林誠(chéng)二, 牧秀明, 等. 2004.長(zhǎng)江干流主要營(yíng)養(yǎng)鹽含量的變化特征—1998-1999年日中合作調(diào)查結(jié)果分析[J]. 地理學(xué)報(bào), 59(l):118-124.
楊漢, 黃艷芳, 李利強(qiáng), 等. 1999.洞庭湖的富營(yíng)養(yǎng)化研究[J]. 甘肅環(huán)境研究與監(jiān)測(cè), 12(3): 120-122.
張亞克, 梁霞, 方焰星, 等. 2011.淀山湖浮游藻類(lèi)增長(zhǎng)的氮磷限制性營(yíng)養(yǎng)研究[J]. 環(huán)境化學(xué), 30(10): 1743-1750.
朱廣偉. 2009.太湖水質(zhì)的時(shí)空分異特征及其與水華的關(guān)系[J]. 長(zhǎng)江流域資源與環(huán)境, 18(5): 439-445.
莊大昌, 丁登山, 董明輝. 2003.洞庭湖濕地資源退化的生態(tài)經(jīng)濟(jì)損益評(píng)估[J]. 地理科學(xué), 23(6): 680-685.
猜你喜歡
小獼猴智力畫(huà)刊(2022年9期)2022-11-04 02:31:42
中學(xué)生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
今日農(nóng)業(yè)(2021年11期)2021-08-13 08:53:34
小哥白尼(趣味科學(xué))(2019年6期)2019-10-10 01:01:50
中國(guó)生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
作文周刊·小學(xué)一年級(jí)版(2017年17期)2017-06-27 19:32:55
廣東第二課堂·小學(xué)(2017年2期)2017-02-20 15:10:08
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
