舟山群島2種瀕危植物生境特征與遷地保護研究
2014-05-29 13:15:20王鳳英彭紅玲胡永紅
浙江農林大學學報 2014年3期
王鳳英,田 旗,彭紅玲,胡永紅
(中國科學院 上海辰山植物科學研究中心/上海辰山植物園,上海 201602)
海洋島嶼生態系統因具有明顯的海域地理隔離而區別于陸地生態系統,被譽為生物地理與進化生態學研究的 “天然實驗室”[1]。島嶼物種由于分布范圍局限、生境脆弱且種群規模較小,較陸地物種具有更高的滅絕風險[2-3]。17世紀至20世紀地球上滅絕的維管植物共計384種,其中島嶼植物達139種,約占36%;漸危或瀕危維管植物中島嶼分布種類約占40%[4]。舟山群島位于長江口以南,杭州灣外緣的東海海域,地史較為年輕,至今為7000~9000 a[5]。舟山群島自與大陸脫離后,歷經數千年海洋性氣候的作用,在植物區系性質上已與大陸形成一定差異,歷來為中外學者所關注[6]。舟山群島自然分布的瀕危植物達21種,其中普陀樟Cinnamomum japonicum var.chenii和舟山新木姜子Neolitsea sericea為國務院1999年批準的《國家重點保護野生植物名錄:第1批》中列入的國家二級重點保護植物[7-8]。普陀樟系樟科Lauraceae樟屬Cinnamomum常綠喬木,間斷分布于中國東部沿海島嶼,自上海金山縣大金山島西坡延伸至浙江舟山群島,中國臺灣、朝鮮、日本也有分布,常見于海拔200~400 m的低山溝谷常綠闊葉林內[9-10]。舟山新木姜子為樟科新木姜子屬Neolitsea常綠木本孑遺植物,分布于浙江舟山群島(上海崇明佘山嶼曾有分布的記錄)海拔150~350 m的丘陵谷地,中國臺灣、日本以及朝鮮半島的沿海島嶼也有分布[11]。由于這2種植物具有很高的觀賞價值,被廣泛開發用于園林綠化。近些年來人為挖掘現象加重,人為干擾可能是導致這2種植物種群數量急劇減少的最主要原因。鑒于島嶼生境的脆弱性所導致的物種瀕危狀態日趨嚴峻,對島嶼植物尤其是島嶼瀕危植物的保護迫在眉睫。歷史的經驗和現狀告訴我們,稀有瀕危物種需要在人類的幫助下才能得到有效的保護,遷地保護就是其中最主要的措施之一[12]。植物園是遷地保護稀有瀕危植物最主要的場所。綜合氣候特點、生境特點、保護區工作的便利等各方面因素,將辰山植物園內的辰山西北坡作為保存物種的遷地保護區,將普陀樟和舟山新木姜子的野外種群遷地到辰山上長期保護及繁衍。
1 材料與方法
1.1 材料
本研究主要以舟山群島野生群落中的普陀樟和舟山新木姜子為研究對象,對其自然分布、群落組成、土壤等生境特點進行了調查與分析,同時于2008-2009年將這2種植物遷地保護于上海辰山植物園內。
1.2 研究方法
1.2.1 群落調查 在物種所在群落內設置10 m×10 m的樣方,調查了普陀島、桃花島、朱家尖、大貓島、懸鵓鴣島、東白蓮島及西峰島等分布區以及遷地保護區的群落組成,用每木調查法統計樣方內的喬木和灌木植物。在10 m×10 m的大樣方內隨機選擇5個1 m×1 m的小樣方,記錄樣方內草本層植物。
1.2.2 生境土壤特征 在野外進行2種植物的群落調查時,舟山群島分布有普陀樟和舟山新木姜子的幾個島嶼(普陀島、桃花島、朱家尖、懸鵓鴣島、東白蓮島、西峰島、大貓島、梅西島)及遷地保護區分別進行土壤物理性狀調查,在樣方內沿樣方對角線兩端和中間用混合采樣法采集0~30 cm土樣,混合均勻后帶回風干后,送上海市園林科學研究所土壤分析室分析化學性質。土樣測定方法:pH值用LY/T 1239-1999《森林土壤pH值的測定》方法測定;電導率用LY/T 1251-1999《森林土壤水溶性鹽分分析》方法測定;有機質用LY/T 1237-1999《森林土壤有機質的測定及碳氮比的計算》方法測定;堿解氮用LY/T 1229-1999《森林土壤水解性氮的測定》方法測定;有效磷用LY/T 1233-1999《森林土壤有機磷的測定》測定;速效鉀用LY/T 1236-1999《森林土壤速效鉀的測定》測定。
1.2.3 物候觀測與記錄 對新定植的植物定期進行物候觀測,主要記錄葉變化期 [芽萌動期、新葉出、幼葉期(顏色)、成葉期(顏色)],花變化期[花蕾(顏色)、初花期(顏色)、盛花期、落花期],果變化期[幼果出現(顏色),果實成熟(顏色)、果落期]。
2 結果與分析
2.1 自然生境與遷地保護區生境特征比較
2.1.1 自然環境氣候特征 舟山群島氣候屬北亞熱帶南緣海洋性季風氣候,年平均氣溫為15.6~16.6℃,極端最低氣溫為-7.9℃,年平均降水量為936.3~1330.2 mm,年平均日照時數為1941~2257 h,無霜期為251~303 d,由于受季風不穩定性影響,夏季易受熱帶風暴(臺風)侵襲,冬季多大風,7-8月間常遇干旱[8]。遷地保護區屬于亞熱帶季風氣候,受冷暖空氣影響,四季分明,氣候溫和,雨水充沛,日照充足,無霜期長。年平均氣溫為15.4℃,極端最低日氣溫-10.5℃,年平均降水量1103.2 mm,年平均日照時數為1997.1 h,年均無霜期為230 d。遷地保護地選在上海辰山植物園的辰山上,辰山山體總高71 m。
2.1.2 群落特征 筆者對舟山群島普陀樟和舟山新木姜子的幾個樣地進行了調查與分析。普陀樟所處群落中喬木層以普陀樟,樸樹Celtis sinensis,天仙果Ficus beecheyana和紅楠Machilus thunbergii為主,它們的重要值分別為73.98,42.48,40.43和30.21,這些物種在群落中占有優勢地位;灌木層普陀樟幼苗占優勢,其他灌木種類較少;草本層中以燈臺蓮Arisaema bockii,大吳風草Farfugium japonicum等植物比較豐富[13]。舟山新木姜子所處群落中(表1)喬木層以舟山新木姜子,杉木Cunninghamia lanceolata,野鴉椿Euscaphis japonica,紅楠和樸樹為主,這些物種的重要值分別為75.10,35.00,31.11,31.01和24.55,在群落中占有優勢地位;灌木層舟山新木姜子苗和茶樹Camellia sinensis苗略微占優勢;草本層以鱗毛蕨屬Dryopteris植物及舟山新木姜子小苗占優勢。從對辰山遷地保護區的原生境群落調查結果分析(表2),喬木層以刺槐Robinia pseudoacacia,樟樹Cinnamomum camphora,構樹Broussonetia papyrifera,樸樹,冬青Ilex chinensis等為主,灌木層中小蠟Ligustrum sinense占優勢,草本層主要以野菊花Dendranthema indicum和蓬藟Rubus hirsutus為主。

表1 舟山新木姜子群落喬木層樹種的群落特征Table1 Characteristics of species for tree layer of Neolitsea sericea community
2.1.3 土壤特征 由表3可見:普陀樟原生境砂石少,土層厚度大于30 cm,容重為1.0~1.2 g·cm-3,孔隙度較大。pH 4.70~5.23,典型酸性土壤,電導率為0.04~0.06 mS·cm-1,有機質為 20.67(懸鵓鴣島)~48.84 g·kg-1(桃花島),水解性氮質量分數為47.45(朱家尖)~163.79 mg·kg-1(普陀島),有效磷質量分數為 4.64(西峰島)~13.37 mg·kg-1(桃花島),速效鉀質量分數為 66.99(東白蓮島)~295.7 mg·kg-1(普陀島)。舟山新木姜子野生生境砂石較多,土層厚度約30 cm。土壤養分狀況良好,舟山新木姜子原生境土壤主要為壤土,其中以黏壤土居多。酸性土壤,pH 4.82~5.38,電導率為0.04~0.10 mS·cm-1,有機質質量分數為 24.18 (朱家尖)~69.31 g·kg-1(桃花島)。水解性氮質量分數為 45.55(朱家尖)~230.20 mg·kg-1(桃花島),有效磷質量分數為5.48(大貓島)~56.09 mg·kg-1(梅西島),速效鉀質量分數為60.80(朱家尖)~371.05 mg·kg-1(桃花島)。遷地保護地土壤以粉砂質黏壤土和粉砂質壤土為主,個別地方也有砂土和黏土。山體上較厚的枯枝落葉層,成為植株生長所需營養的儲存庫。大部分土層厚度約60 cm,個別山石多的地方僅20 cm。辰山土壤pH 6.01,電導率為0.05 mS·cm-1,有機質為58.99 g·kg-1,水解性氮質量分數為221.21 mg·kg-1,有效磷質量分數為20.52 mg·kg-1,速效鉀質量分數為69.30 mg·kg-1。
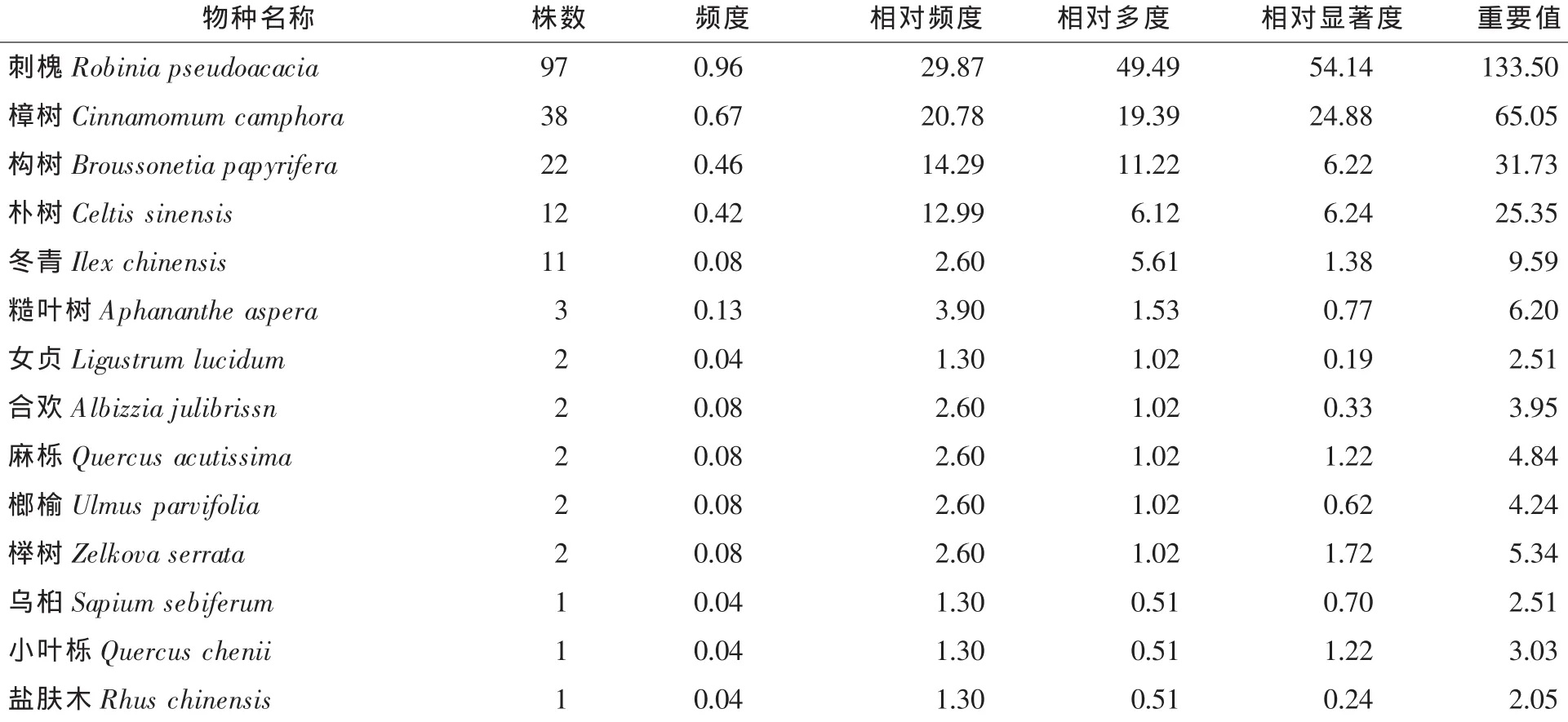
表2 遷地保護區群落喬木層樹種的群落特征Table2 Characteristics of species for tree layer of Cinnamomum japonicum var.chenii community

表3 普陀樟和舟山新木姜子原生境和遷地保護區生境土壤特征比較Table3 Soil characteristic indices of Cinnamomum japonicum var.chenii and Neolitsea sericea between the natural and ex-situ habitats
2.2 遷地保護區的建立
2.2.1 遷地保護區改造 為給遷地保護植物足夠的生長空間,對原有群落進行疏伐除雜,主要采取間疏式伐木,重點清理刺槐、構樹及帶刺類灌木,適當修剪大樹樹冠,為定植植株提供充足的陽光和合適的生態位。隨著這些植物的生長再逐漸去枝、疏伐,以滿足植株在不同發育階段對光照等環境條件的要求。
2.2.2 植物遷地保護群落的組建 群落的組成與結構的設計要滿足保護物種及其遺傳多樣性的要求。不僅要保證不同物種不同發育階段的植物生長所要求的條件,使其能正常繁育,最終達到從種子到種子的自行繁衍;還需保證整個群落的穩定性及不同植物間的相互協調性,對群落中的主要優勢種群能保證其持續穩定的發展,且能基本上保持其遺傳組成與遺傳結構的穩定[14]。由于大的喬木引種困難,而且成活率低,在群落建造初期,主要以來源于野外實生苗和野外采種播種繁殖的小苗為主。因此,初建成的群落仍以原生境物種為優勢種。為保證普陀樟和舟山新木姜子完整的遺傳多樣性,盡可能收集了多個分部區的植株。普陀樟引自舟山群島的桃花島、懸鵓鴣島、西峰島、東白蓮島和朱家尖等5個分布區,舟山新木姜子引自舟山群島的大貓島、東白蓮島、朱家尖以及桃花島的大佛巖和對峙山。于2008-2009年共種植了普陀樟76株,舟山新木姜子67株,滿足了最小可存活種群50株的要求。根據對這2種植物野外群落調查的結果,栽植以下優勢伴生種:紅楠、樸樹、野鴉椿、天仙果和鱗毛蕨科Dryopteridaceae植物等。苗木定植后,初期對每株植株搭三腳架起支撐防風作用,冬季將樹干用麻袋片綁好保溫,并且用草簾搭防風墻,在定植后1 a之內,對植株進行了精細養護,及時對植株葉片和樹干進行噴水,視具體情況進行澆水,在生長速度最快的季節進行施肥,樹穴內及周圍經常除草松土,伐去妨礙植株生長的喬、灌枝條,遇到病蟲害及時采取措施。2010年的統計結果表明普陀樟和舟山新木姜子在遷地保護區的成活率分別為91%和88%。
2.2.3 物候觀測記錄 對遷地引種的普陀樟和舟山新木姜子連續2 a的物候觀測與記錄(表4),基本掌握了它們在遷地保護區的物候期。普陀樟4-6月抽新葉,5月開花,10-11月結果。舟山新木姜子4-6月抽新葉,11-12月開花,第2年10-12月結果。
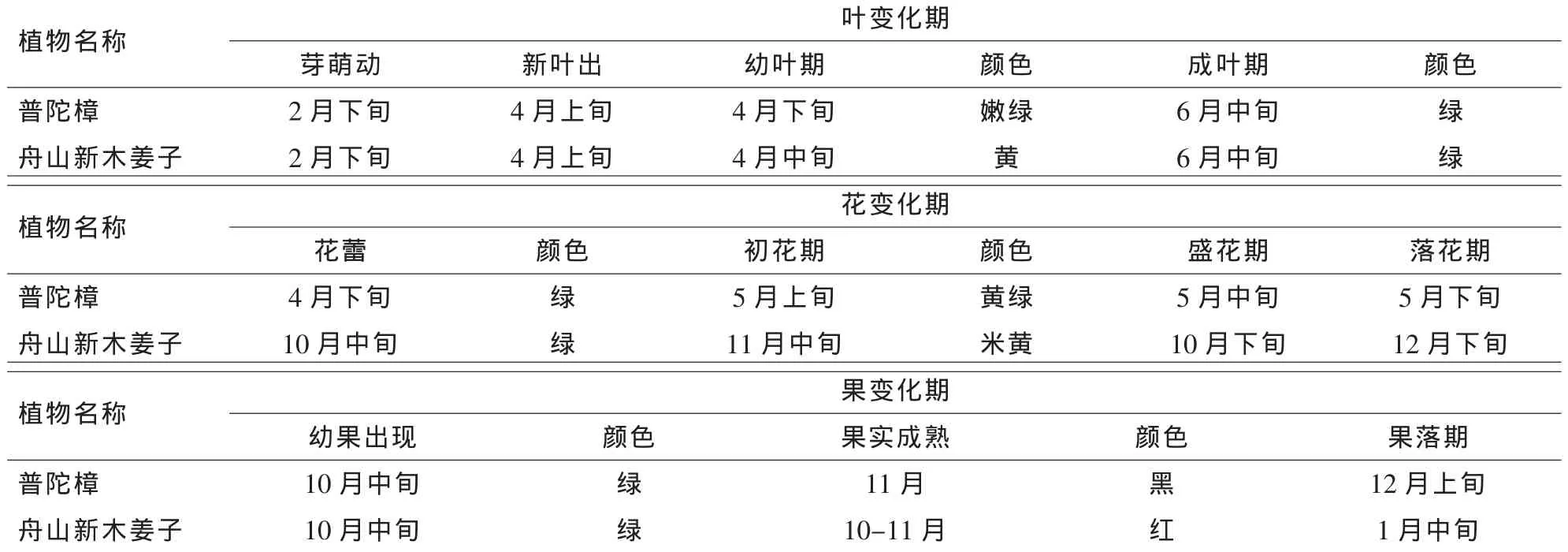
表4 普陀樟和舟山新木姜子在遷地保護區的物候記錄Table4 Phenological period of Cinnamomum japonicum var.chenii and Neolitsea sericea in ex-situ conservation
3 結論與討論
3.1 自然生境與遷地保護區生境特征比較
首先,根據20世紀初期德國慕尼黑大學邁依爾提出的 “氣候相似論”,木本植物引種成功的最大可能性在于樹種原生地和新栽培地區的氣候條件有相似的地方。舟山群島和上海辰山植物園均隸屬于華東東部地區,經緯度十分接近,均屬于亞熱帶氣候,年平均溫度約為15℃,年平均降水量約1000 mm,年平均日照時數約2000 h,溫度、降水、日照等氣候參數都很接近,滿足了氣候相似的原則。其次,普陀樟和舟山新木姜子野外所處的群落各層次物種種類有一定程度的相似性,與遷地保護區群落物種相似性不高,但我們在引種普陀樟和舟山新木姜子的同時,還引種了原生境的伴生種,營造了與野生群落相似度較高的群落環境。此外,在生態條件相似的遷地保護過程中,土壤條件也是必不可少的因素,普陀樟、舟山新木姜子原生境與遷地保護區土壤均呈酸性。普陀樟、舟山新木姜子原生境所有樣點有機質質量分數均高于20 g·kg-1(園藝發達國家最低標準),說明普陀樟和舟山新木姜子原生境土壤有機質豐富,而遷地保護區辰山上的土壤有機質質量分數達到58.99 g·kg-1,足以能保證遷地植物的正常生長。土壤速效養分是衡量土壤養分供應能力的現實指標。從樣點分布來看,桃花島樣地各速效養分質量分數整體處于較高水平,朱家尖較其他樣地來說偏低,遷地保護區除速效鉀質量分數稍偏低外,其他2項指標均處于較高水平。以上土壤理化特征顯示普陀樟和舟山新木姜子原生境與遷地保護區土壤酸堿性一致,原生境土壤相關肥料指標與遷地保護區也基本相似,足以滿足普陀樟、舟山新木姜子的生長。綜上所述,普陀樟和舟山新木姜子原生境與遷地保護區生境中氣候條件、群落特征、土壤特征均有很大程度的相似性,有利于遷地保護的成功。
3.2 遷地保護區的建立
遷地保護群落垂直結構的形成是一個動態的過程。在群落建造初期,原生境中的物種起著至關重要的作用,隨著群落的動態變化,遷地保護植物在群落中的作用和地位會逐漸凸顯。采用逐步疏伐的方法是根據不同植物的生態生物學特性為各物種選擇合適的生態位,并在植株生長發育過程中及時去枝或疏伐,以滿足植株在不同發育階段對光照等環境條件的要求。經適當疏伐后,基本上保留著原生境群落的垂直結構不變,新栽植的小苗在群落中只是充當了灌木層和草本層的角色。引進喬木植物的種群結構在群落建造初期因無法滿足不同年齡級的個體要求,故需要采用分期分批定植的方案,在植物不斷生長過程中逐步完善各種群的年齡結構。
遷地保護的最終目的是要保護多樣性的物種遺傳基因。普陀樟和舟山新木姜子各引種了5個種群,多個種群的引入不僅能夠更完整地保存物種的遺傳多樣性,還有利于基因交流,突破了傳統的引種來源單一問題。
3.3 遷地保護有效性評價
遷地保護的有效性評價需綜合考慮數量、材料來源、適應性等多方面因素。本研究遷地保護了普陀樟和舟山新木姜子分別為76和67株,滿足了遷地保護最小種群大于50株的理論要求[16]。分別引入5個不同來源地的種群,滿足了保存物種遺傳多樣性的要求。此外,經后期觀察與物候記錄,普陀樟和舟山新木姜子已經在遷地保護正常開花與結果。從目前的理論研究及植物實際的長勢來觀察,遷地保護獲得了一定的成效。
[1]魏娜,王中生,冷欣,等.海洋島嶼生物多樣性保育研究進展[J].生態學雜志,2008,27(3):460-468.WEI Na,WANG Zhongsheng,LENG Xin,et al.Conservation of oceanic island biodiversity∶a review [J].Chin J Ecol,2008,27(3)∶460 - 468.
[2]FRANKHAM R.Do island populations have less genetic variation than mainland populations [J]Heredity,1997,78∶311-327.
[3]RAVEN P H.Plant conservation in a changing genetics of endangered island plants [M]//AVISE J C,HAMRICK J.Conservation Genetics:Case Histories from Nature.New York∶Chapman and Hall,1998:305 - 334.
[4]REID W V,MILLER K R.Keeping Options Alive:The Scientific Basis for Conserving Biodiversity [R].Washington D C∶World Resources Institute,1989.
[5]LI Yiming,NIEMELA J,LI Dianmo.Nested distribution of amphibians in the Zhoushan archipelago,China∶can selective extinction cause nested subsets of species[J].Oecologia,1998,113∶557 - 564.
[6]萬利琴,丁炳揚,郭水良.舟山群島主要島嶼間種子植物區系的差異及其影響因素[J].浙江大學學報:農業與生命科學版,2008,34(6)∶677-683.WAN Liqin,DING Bingyang,GUO Shuiliang.Difference of spermatophyte flora among main islands of Zhoushan Archipelago and its influencing factors [J].J Zhejiang Univ Agric&Life Sci,2008,34(6)∶677 - 683.
[7]傅立國.中國植物紅皮書——稀有瀕危植物:第1冊[M].北京:科學出版社,1991.
[8]俞慈英,陳葉平,袁燕飛,等.舟山群島普陀樟等3種特有樹種種質資源清查[J].浙江林學院學報,2007,24(4):413 - 418.YU Ciying,CHEN Yeping,YUAN Yanfei,et al.Resource s of Cinnamomum japonicum var.chenii,Neolitsea sericea and Tilia miqueliana in Zhoushan archipelago [J].J Zhejiang For Coll,2007,24(4)∶413 - 418.
[9]王景祥.浙江植物志:第2卷[M].杭州:浙江科學技術出版社,1992:352.
[10]張若蕙.浙江珍稀瀕危植物[M].杭州:浙江科學技術出版社,1994:156.
[11]王中生,安樹青,冷欣,等.島嶼植物舟山新木姜子居群遺傳多樣性的RAPD分析[J].生態學報,2004,24(3):414 - 422.WANG Zhongsheng,AN Shuqing,LENG Xin,et al.Population genetic diversity of the insular plant Neolitsea sericea based on random amplified polymorphic DNA (RAPD)[J].Acta Ecol Sin,2004,24(3)∶414 - 422.
[12]康明,葉其剛,黃宏文.植物遷地保護中的遺傳風險[J].遺傳,2005,27(1):160-166.KANG Ming,YE Qigang,HUANG Hongwen.Genetic risks in plant ex-situ conservation [J].Hereditas,2005,27(1)∶160-166.
[13]田旗,黃姝博,胡永紅.中國特有植物普陀樟群落特征及物種多樣性[J].植物分類與資源學報,2010,32(增 17):81- 88.TIAN Qi,HUANG Shubo,HU Yonghong.Characteristics and species diversity of Cinnamomum japonicum var.chenii(Lauraceae) community in China [J].Plant Diversity Resour,2010,32(supp 17)∶81 - 88.
[14]葉其剛,王暢,王詩云.三峽庫區稀有瀕危植物異地保護群落設計的初步研究[J].武漢植物學研究,2000,18(1):33 - 41.YE Qigang,WANG Chang,WANG Shiyun.A preliminary study on the design of a community for ex-situ conservation of the rare and endangered plants in the Three Gorge Reservoir Area [J].J Wuhan Bot Res,2000,18(1)∶33 - 41.
[15]許再富.稀有瀕危植物遷地保護的原理與方法[M].昆明:云南科技出版社,1998.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
