枯草芽孢桿菌BJ3-2精氨酸脫羧酶基因speA的克隆與序列分析
2014-04-24 11:31:40劉艷敏沈璽龍吳擁軍
中國釀造 2014年5期
劉艷敏,盧 彪,沈璽龍,王 濤,吳擁軍*
(貴州大學 生命科學學院,貴州 貴陽 550025)
精氨酸脫羧酶(arginine decarboxylase,ADC)廣泛分布于細菌、真菌、古生菌和植物體內,在多胺生物合成過程中扮演重要的角色[1],是多胺合成途徑中關鍵酶[2-3]。在枯草芽孢桿菌(Bacillus subtilis)中多胺生物合成只有一條途徑:始于精氨酸(arginine)[4],由精氨酸脫羧酶作用脫羧生成的胍基丁胺(agmatine),經過胍基丁胺酶(agmatinase)水解生成腐胺(putrescine,Put)和尿素(urea)[1,5];腐胺與脫羧S-腺苷甲硫氨酸(dAdoMet,dSAM)[6]提供的氨丙基在亞精胺合成酶(spermidine synthase)或精胺合成酶(spermine synthase)的催化作用下合成亞精胺(spermidine,Spd)或精胺(spermine,Spm)[2,7-9]。適量的生物胺在人和動物活細胞中發揮著重要的生理作用,但當體內生物胺含量過高或超過一定范圍則會引起身體不適等不良癥狀,還具有潛在的致癌性[10]。生物胺存在于食品中也會影響甚至改變其風味成分[11-12];同時精胺和亞精胺的存在會影響食品的風味和品質,產生難聞的氣味,還可能與亞硝酸鹽結合形成致癌的亞硝胺[10]。多聚胺如腐胺、尸胺、精胺和亞精胺等對人體沒有直接毒性,但會加強組胺和酪胺對人體解毒系統的影響。
目前貴州地區水豆豉生產主要以自然發酵為主,其中發酵的主要微生物枯草芽孢桿菌,擁有悠久的酶生產和食品發酵歷史,是細菌型水豆豉生產的主要食品安全發酵菌株。發酵食品中的生物胺主要由微生物通過分泌脫羧酶而生成,據報道芽孢桿菌具有氨基酸脫羧酶活性,張建華等[13]報道了納豆發酵過程中產生的生物胺主要以亞精胺為主。納豆和水豆豉都屬于發酵大豆食品,為控制水豆豉生產過程產生過量的亞精胺和精胺及其副產物,以實驗室前期已獲得豆豉生產菌株Bacillus subtilisBJ3-2[14-15]為目標菌株,根據GenBank中枯草芽孢桿菌168菌株的精氨酸脫羧酶基因序列設計特異性引物,以Bacillus subtilisBJ3-2基因組DNA為模板進行PCR擴增,克隆精氨酸脫羧酶基因speA,并與其他枯草芽孢桿菌的speA序列進行同源性比對分析,推測ADC酶活性結構域和酶活性位點,從而為產亞精胺和精胺的枯草芽孢桿菌的檢測提供理論依據,也為ADC活性影響因素的進一步研究奠定基礎,為水豆豉的安全生產提供一定的指導意義。
1 材料與方法
1.1 材料與試劑
Bacillus subtilisBJ3-2和E.coliDH5α:本實驗室保存。
pGEM-T載體:美國Promega公司;基因組DNA提取試劑盒、溶菌酶:美國Promega公司;Taq(聚合)酶、T4 DNA連接酶、DNAMarker、dNTPs、10×PCRBuffer等:大連TaKaRa公司;其他試劑均為國產分析純。E.Z.N.A.TMGel Extraction Kit(200)膠回收試劑盒、E.Z.N.A.TMPlasmid Mini KitⅠ(200)質粒提取試劑盒:美國Omega公司。
1.2 儀器與設備
MyCycler PCR儀、Universal Hood Ⅱ凝膠成像分析儀、PowerPac HC電泳儀:美國Bio-Rad公司。
1.3 引物設計與合成
參考GenBank中枯草芽孢桿菌168菌株(登錄號為AL009126.3)的精氨酸脫羧酶基因序列,應用DNAStar軟件設計一對精氨酸脫羧酶基因的特異性引物,引物序列:上游引物P1:5′-TTGTCGCAACATGAAACAC-3′;下游引物P2:5′-TTATTGAATTGCTTTTTGTTC-3′;預計擴增片段長度為1 473bp,由北京諾賽基因組研究中心有限公司合成。
1.4 方法
1.4.1Bacillus subtilisBJ3-2基因組DNA的提取
挑取B.subtilisBJ3-2單菌落,接種到5mL LB培養基中,37℃條件下振蕩培養過夜,參考Promega公司的革蘭氏陽性細菌中提取基因組的試劑盒說明書提取全基因組。
1.4.2 PCR擴增
以提取的B.subtilisBJ3-2的基因組DNA為模板,用合成的引物進行PCR擴增,PCR反應體系:DNA模板1μL、dNTPs 1.6μL、10×PCR Buffer 2μL、上下游引物各0.5μL、Taq酶0.1μL、ddH2O 14.3μL。總體系:20μL。反應程序:94℃預熱5min,94℃變性40s,50℃退火40s,72℃延伸90s,進行30個循環后,72℃延伸10min,擴增產物經1%瓊脂糖凝膠電泳檢測。
1.4.3 精氨酸脫羧酶基因(speA)的克隆與鑒定
將回收純化的PCR產物與pGEM-T載體按常規方法[16]進行連接并轉化感受態細胞E.coliDH5α。將菌落PCR和質粒PCR驗證正確的克隆質粒送北京諾賽基因組研究中心有限公司測序。
1.4.4speA序列分析
用DNAStar和MEGA5生物學軟件對speA基因的核苷酸序列及推導的氨基酸序列進行編輯,與GenBank中已發表的其他speA基因序列進行同源性分析并繪制ADC遺傳進化樹。
2 結果與分析
2.1 Bacillus subtilis BJ3-2基因組DNA的提取
用基因組提取試劑盒,對B.subtilisBJ3-2基因組DNA進行提取,DNA樣品經0.7%瓊脂糖凝膠電泳檢測。圖1顯示所提取的B.subtilisBJ3-2基因組DNA較完整,無顯著降解,可用作模板進行下一步目的基因的PCR擴增。

圖1 枯草芽孢桿菌BJ3-2基因組Fig.1 Genome of Bacillus subtilis BJ3-2
2.2 speA基因的PCR擴增
以B.subtilisBJ3-2基因組DNA為模板,PCR擴增speA基因,1%的瓊脂糖凝膠電泳,擴增出一條約與預期片段大小1 473bp相符的條帶,結果見圖2。

圖2 PCR擴增speA基因Fig.2 PCR amplification of speA
2.3 重組克隆質粒的菌落PCR鑒定
PCR產物純化后與pGEM-T 載體連接過夜,轉化E.coliDH5α。通過菌落PCR檢測,從圖3可以看出,約在1 473bp處出現了特異性擴增條帶,與基因組PCR擴增speA的目的條帶大小相吻合,獲得了陽性的重組菌落。
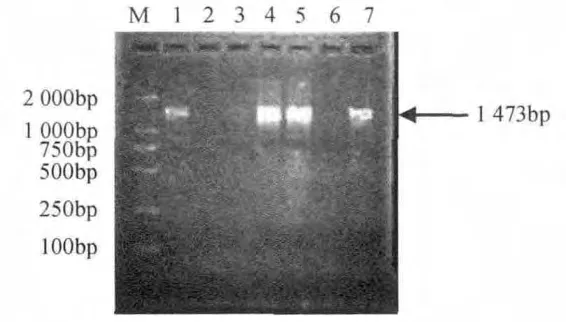
圖3 菌落PCR鑒定重組子Fig.3 PCR rapid identification of speA gene from recombinant clones
2.4 重組質粒PCR鑒定
將鑒定的陽性重組菌落進行增菌培養,提取質粒DNA,進行質粒PCR擴增,結果表明,約在1 473bp處出現一特異性條帶,所獲得的基因片段大小均與PCR擴增產物大小相吻合(見圖4),說明目的基因已正確克隆至pGEM-T載體中,并將提取的重組質粒送生物公司測序,并命名為pGEM-speA。
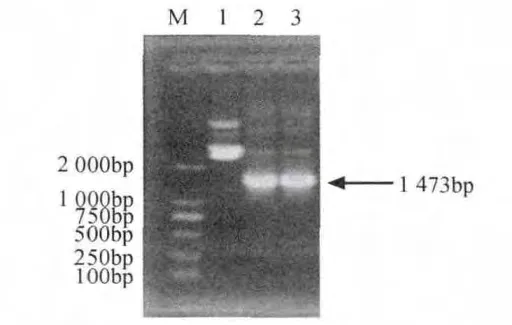
圖4 質粒PCR鑒定Fig.4 PCR identification of speA from recombinant plasmid pGEM-speA
2.5 Bacillus subtilis BJ3-2精氨酸脫羧酶基因speA的同源性分析及系統進化樹
測序結果經DNAStar 軟件編輯處理,結果表明,B.subtilisBJ3-2精氨酸脫羧酶基因speA開放閱讀框大小為1 473bp,編碼490個氨基酸,推測蛋白質分子質量為53.53ku,G+C含量為44%,等電點為5.29,酸性氨基酸19.6%,堿性氨基酸12.2%。將序列提交至GenBank中,獲得基因登錄號為KJ561348。
將所克隆的基因序列和推導的蛋白序列與GenBank數據庫進行序列同源性比對,分析結果表明,B.subtilisBJ3-2的speA基因序列與已報道枯草芽孢桿菌的speA基因序列和蛋白序列高度同源,分別達到93%和95%以上,核苷酸序列與Bacillussp.JS(speA)的同源性最高,達99%;與其他枯草芽孢桿菌(speA)的核苷酸序列也在93%及其以上;與解淀粉芽孢桿菌(Bacillus amyloliquefaciens)和萎縮芽孢桿菌(Bacillus atrophaeus)的同源性為79%,表明克隆的基因為編碼精氨酸脫羧酶的基因。
B.subtilisBJ3-2精氨酸脫羧酶氨基酸序列與芽孢桿菌JS(Bacillussp.JS)、枯草芽孢桿菌168菌株(Bacillus subtilis168)、枯草芽孢桿菌納豆亞種BEST195菌株(Bacillus subtilis subsp.natto BEST195)、枯草芽孢桿菌BEST7613(Bacillus subtilisBEST7613)最高,為99%;與其他枯草芽孢桿菌精氨酸脫羧酶氨基酸序列在≥97%;與芽孢桿菌屬其他菌種的精氨酸脫羧酶氨基酸序列同源性在97%~72%之間。
乳酸菌(Lactobacillussp.30a)的鳥氨酸脫羧酶家族(ornithine decarboxylase family)作為大多數細菌磷酸吡哆醛(pyridoxal 5'-phosphate,PLP)依賴型脫羧酶代表,具有Ⅲ型PLP型脫羧酶的活性位點和PLP結合位點以及二級結構特征[17],選擇Lactobacillussp.30a的ODC氨基酸序列作比對,推測B.subtilisBJ3-2的ADC底物結合位點和酶活性位點。研究發現,Ⅲ型PLP型脫羧酶和厚壁菌門細菌的PLP依賴型脫羧酶與大腸桿菌的保守結構區域不同[17-18],嗜熱厭氧解木聚糖桿菌(Thermoanaerobacterium xylanolyticumLX-11)和梭狀芽孢桿菌(Clostridium stercorariumsubsp.stercorariumDSM 8532)都屬于厚壁菌門細菌,選擇二者的ADC序列作比對,增加推測B.subtilisBJ3-2的底物結合位點和酶活性位點的可靠性;并根據大腸桿菌E.coliATCC 8739 ADC 和Lactobacillussp.30a ODC、B.subtilis168 ADC 和T.xylanolyticumLX-11 ADC、C.stercorariumDSM 8532 ADC的氨基酸序列以及同源性較高的枯草芽孢桿菌ADC序列構建系統進化樹,進一步分析其間的進化關系。
ClustalW多序列比對,GeneDoc輸出,結果顯示B.subtilisBJ3-2的ADC基因包括有PLP依賴型脫羧酶家族的氨基酸保守結構域:即PLP結合位點:[FDSAW]、[RNNHKSVYNSA]、[S-X-H-K];[S-X-H-K]序列中Lys(K)是PLP依賴型脫羧酶的活性位點,PLP通過與Lys氨基酸殘基形成醛亞胺鍵(席夫堿),使酶具有催化活性[17],如圖5所示。
從圖6可以看出,枯草芽孢桿菌的ADC位于遺傳距離較近的同一進化分支,彼此親緣關系較近,B.subtilisBJ3-2的與B.subtilis168的ADC遺傳距離最近,與T.xylanolyticumLX-11和C.stercorariumDSM8532的遺傳距離次之。B.subtilisBJ3-2的ADC與Lactobacillussp.30a的ODC位于遺傳距離稍遠的同一進化分支,與E.coliATCC 8739的ADC遺傳距離最遠。以上分析顯示B.subtilisBJ3-2與乳酸菌和其他厚壁菌門細菌的遺傳距離較近,具有Lactobacillus.30a鳥氨酸脫羧酶家族(ornithine decarboxylase family)相似的保守結構域,屬于典型的III型磷酸吡哆醛(pyridoxal phosphate,PLP)依賴型鳥氨酸/賴氨酸/精氨酸脫羧酶家族(Orn/Lys/Arg decarboxylase family)成員[17-18]。

圖5 B.subtilis BJ3-2和其他細菌ADC/ODC的氨基酸多序列比對Fig.5 ADC/ODC amino acid multiple sequence alignments between B.subtilis BJ3-2 and other bacteria

圖6 不同細菌的ADC/ODC氨基酸序列遺傳進化樹Fig.6 Genetic evolutionary tree of ADC/ODC amino acid sequences of different bacteria
3 結論與討論
Bacillus subtilisBJ3-2的ADC基因編碼的脫羧酶屬于丙氨酸消旋酶(alanine racemase)家族,其的功能依賴于PLP位點,屬于典型的III型PLP依賴型鳥氨酸/賴氨酸/精氨酸脫羧酶家族成員。在[FDSAW]結構域中,天冬氨酸Asp(D)與丙氨酸Ala(A)高度保守,Asp在酶催化過程中能穩定電荷,Ala的甲基側鏈處在酶輔助因子的后面,對于酶的空間構象具有重要作用。[S-X-H-K]序列高度保守,組氨酸His(H)在PLP底物結合去質子化過程中提供質子,穩定電荷,該位點的定點突變對酶活性會產生影響[19]。[RNNHKSVYNSA]中His存在所有的PLP依賴型脫羧酶中,代表了轉氨酶向脫羧酶進化的一個最重大的特征,His殘基在特定的組胺酰基試劑修飾后會使酶失活[17,20]。在不同細菌ADC的序列比對中還存在[GTS]、[STSPFY]、[YPPG]、[PGE]高度保守序列,這可能其他輔助因子相關。PLP型輔助因子通過氫鍵、π-堆積以及范德華力相互作用,以確保ADC酶活性,保守結構域和活性位點的分析對于酶活性調控具有重要作用。
在枯草芽孢桿菌中,以ATG為起始密碼子的基因的占78%,以TTG為起始密碼子的占13%,以GTG的占9%。大腸桿菌中的起始tRNA分子可以同時識別AUG、GUG和UUG這3種起始密碼子,但識別頻率并不相同,通常GUG為AUG的50%,而UUG只及AUG的25%[21]。以實驗目的菌株B.subtilisBJ3-2克隆得到的speA基因的起始密碼子為TTG,是枯草芽孢桿菌中特有的遺傳起始密碼子,在精氨酸脫羧酶基因的表達后續試驗中需要選擇合適的表達載體和宿主菌,或在大腸桿菌中表達宿主細胞中進行密碼子優化,增強精氨酸脫羧酶基因的表達效率。
在革蘭氏陽性菌枯草芽孢桿菌中生物多胺合成只有一條途徑,起始于精氨酸,由精氨酸脫羧酶、胍基丁胺酶、S-腺苷甲硫氨酸脫羧酶以及亞精胺合酶、精胺合酶共同參與作用。實驗應該同時研究這幾個基因,以進一步了解枯草芽孢桿菌生物多胺的合成途徑,speA基因的克隆與ADC酶保守結構域和酶活性位點的推測,為有效控制水豆豉產品中生物胺的含量和農業生產以及植物抗性關于精氨酸脫羧酶[6,22]的相關研究提供一定的理論依據。
[1]FOROUHAR F,LEW S,SEETHARAMAN J,et al.Structures of bacterial biosynthetic arginine decarboxylases[J].Acta Crystallogr F,2010,66(12):1562-1566.
[2]TIBURCIO A F,MASDEU M A,GALSTON A W.Polyamine metabolism and osmotic stress[J].J Plant physiol,1986,82(2):369-374.
[3]汪 天,孟艷瓊,谷 鳳,等.多胺代謝及其與農產品貯藏保鮮關系的研究[C].合肥:2005 安徽食品安全博士論壇,2005.
[4]SEKOWSKA A,BERTIN P,DANCHIN A.Characterization of polyamine synthesis pathway inBacillus subtilis168[J].Mol Microbiol,1998,29(3):851-858.
[5]曹學武,高鈺琪.精氨酸-胍基丁胺代謝的研究進展[J].生命科學,2004,16(3):170-172.
[6]張慶福.枳精氨酸脫羧酶基因(PtADC)轉化煙草提高抗脫水研究[D].武漢:華中農業大學碩士論文,2010.
[7]SEKOWSKA A,COPPEE J Y,LE CAER J P,et al.S-adenosylmethionine decarboxylase ofBacillus subtilisis closely related toarchaebacterial counterparts[J].Mol Microbiol,2000,36(5):1135-1147.
[8]EVANS P T,MALMBERG R L.Do polyamines have roles in plant development[J].Annu Rev Plant Physiol Plant Mol Biol,1989,40:235-269.
[9]馬 瑛,劉 靜.植物體中多胺代謝及其功能研究進展[J].陜西理工學院學報:自然科學版,2010,26(2):47-54.
[10]HERNANDEZ-JOVER T,IZQUIERDO-PULIDO M,VECIANA-NOGUES M T,et al.Biogenic amines and polyamine contents in meat and meat products[J].J Agr Food Chem,1997,45(6):2098-2102.
[11]SHALABY A R.Significance of biogenic amines to food safety and human health[J].Food Res Int,1996,29(7):675-690.
[12]ONAL A.A review:current analytical methods for the determination of biogenic amines in foods[J].Food Chem,2007,103(7):1475-1486.
[13]張建華,沈 翔,于湘莉.納豆發酵過程中的生物胺[J].上海交通大學學報:農業科學版,2007,25(1):1-5.
[14]吳擁軍,孟望霓,李耀中.純種強化發酵細菌型豆豉研究[J].中國釀造,2011,30(9):56-60.
[15]賈東旭,吳擁軍,李耀中,等.細菌型豆豉發酵芽孢桿菌的篩選與鑒定[J].食品科學,2009,30(5):217-221.
[16]J.薩姆布魯克D.W 拉塞爾.黃培堂,等譯.分子克隆實驗指南(第三版)[M].北京:科學出版社,2002.
[17]MOMANY C,GHOSH R,HACKERT M L.Structural motifs for pyridoxal-5'-phosphate binding in decarboxylases:an analysis based on the crystal structure of theLactobacillus30a ornithine decarboxylase[J].Protein Sci,1995,4(5):849-854.
[18]BURRELL M,HANFREY C C,MURRAY E J,et al.Evolution and multiplicity of arginine decarboxylases in polyamine biosynthesis and essential role inBacillus subtilisbiofilm formation[J].J Biol Chem,2010,285(50):39224-39238.
[19]GANI D.A structural and mechanistic comparison of pyridoxal 5'-phosphate dependent decarboxylase and transaminase enzymes[J].Philos T Roy Soc B,1991,332(1263):131-139.
[20]MISHIN A A,SUKHAREVA B S.Glutamate decarboxylase fromEscherichia coli:catalytic role of the histidine residue[J].Dokl Akad Nauk SSSR,1986,290(5):1268-1271.
[21]白俊琴.枯草芽孢桿菌lipA 基因表達的研究[D].天津:天津大學碩士論文,2009.
[22]郭玉蘭.精氨酸脫羧酶活性與冬小麥抗旱性關系的研究初報[J].河北師范大學學報,1990(1):77-78.
