外源一氧化氮介導銅脅迫下番茄幼苗中鐵、 鋅、 錳的累積及亞細胞分布
2014-04-08 09:19:14梁國鵬姜春輝崔秀敏
植物營養與肥料學報 2014年3期
張 敏, 梁國鵬, 姜春輝, 崔秀敏
(土肥資源高效利用國家工程實驗室, 山東農業大學資源與環境學院,山東泰安 271018)
銅(Cu)是植物正常生命活動所必需的微量營養元素,廣泛參與各種生命活動。同時,銅也是一種重金屬元素,過量的 Cu 會對植物產生毒害作用,如破壞生理結構、 引發代謝紊亂、 抑制植物生長發育等問題[1-4]。目前,在農業生產中,污水灌溉和含銅的肥料及農藥的大量使用,導致土壤系統中 Cu 不斷累積,明顯高于其背景值[5]。
一氧化氮(nitric oxide,NO)是廣泛分布于生物體內的小分子信號物質。研究發現 NO參與植物的生長發育和對外界脅迫反應等多種生理過程。NO 能夠提高逆境脅迫下植物抗性和生物量[6-7],維持組織結構穩定[8],參與植物激素的調控,提高植物對過多銅的螯合能力[9]。鐵(Fe)、 鋅(Zn)、 錳(Mn),作為微量元素亦在植物生命代謝中發揮著重要作用。如 Fe 元素參與光合作用和固氮過程,并在葉綠素合成中起關鍵作用; Zn 是植物體 59 種復合酶的組分或活化劑,并參與生長素代謝; Mn對種子發芽、 幼苗生長、 花粉管發育和植物莖的機械強度均有重要作用[10]。Fe2+、 Zn2+、 Mn2+在植物根系吸收及轉運過程中與 Cu2+存在復雜的相互作用。研究發現,銅脅迫下,外源 NO 可調控番茄組織與細胞內過多的銅趨于更合理的分布[11],勢必也會影響帶有相同電荷的Fe2+、 Zn2+、 Mn2+的宏觀及微域分布。
本文以番茄為試驗材料,采用營養液培養,研究外源 NO 調控的 Cu 脅迫下番茄幼苗中 Fe、 Zn、 Mn 的累積及亞細胞分布,探索 Cu 脅迫下外源 NO 介導的番茄的營養機理。
1 材料和方法
1.1 供試材料
供試番茄(SolanumlycopersicumL.)為“改良毛粉802F1”。Hoagland營養液組成(mmol/L): Ca(NO3)2-4H2O 1.25、 KNO30.5、 NH4NO30.73、 KH2PO40.5、 H3BO34.6×10-2、 Na2MoO42.6×10-3、 MgSO41.0、 EDTA-Fe 2×10-2、 MnCl29×10-3、 ZnSO47.6×10-4,以上試劑均為分析純,用蒸餾水配制。NO和Cu2+的供體及適宜濃度由預備試驗完成, CuCl2提供Cu2+,硝普鈉[Na2Fe(CN)5、 NO、 SNP購自Sigma公司]提供NO,先用蒸餾水配成200 μmol/L的母液,4℃避光保存,用時按所需濃度稀釋。牛血紅蛋白(Hb,購自Sigma公司)為NO的清除劑。
1.2 試驗設計

1.3 測定項目與方法
1.3.1 生物量的測定 處理后第 8 d,移出番茄幼苗,測定各處理植株的株高,并將植株分為根系、 莖、 葉柄、 葉片四部分,用去離子水沖洗干凈,吸水紙吸干表面水分,稱鮮重。
1.4 數據處理
采用 Fe、 Zn、 Mn 轉運因子(transport factor,TF) 計算 Fe、 Zn、 Mn 從根到地上部的運輸特征[14]:
TF=(Cstem+Cleaf+Cpetiole) /Croot
式中: Cstem、 Cleaf、 Cpetiole和 Croot分別表示莖、 葉片、 葉柄和根系中的 Fe、 Zn、 Mn 的累積量。
采用Microsoft Excel軟件對數據進行處理及繪圖,采用 DPS 軟件進行數據統計分析,采用最小顯著差異法(LSD)進行多重比較和差異顯著性檢驗(α=0. 05)。
2 結果與分析
2.1 外源 NO 對 Cu 脅迫下番茄幼苗生長的影響
從圖 1 可以看出,不同處理下番茄幼苗的生物量和株高均有顯著差異。與對照(CK)相比,Cu 脅迫處理(Cu)下番茄幼苗生物量和株高顯著下降了33.7% 和 23.1%。添加外源 SNP (Cu+SNP)顯著緩解了 Cu 脅迫引起的生長抑制,使番茄幼苗的生物量幾乎恢復到對照水平,株高雖然沒有恢復到對照水平,但仍顯著高于Cu 脅迫處理。添加血紅蛋白后(Cu+SNP+Hb),顯著消除了 NO 的緩解效果,說明外源 NO 可以促進 Cu 脅迫下番茄幼苗的生長發育。

圖1 外源一氧化氮對銅脅迫下番茄幼苗生物量和株高的影響Fig.1 Effects of exogenous NO on the biomass and height of tomato seedlings under Cu stress
2.2 外源 NO 對 Cu 脅迫下番茄幼苗 Fe 含量和累積量的影響
由表 1 可以看出,番茄幼苗各部位的 Fe 含量和累積量變化幅度較大,根系和葉柄的 Fe 含量和累積量最高。不同處理間各部位 Fe 含量和累積量差異顯著。與對照(CK)相比,Cu 脅迫處理顯著降低番茄幼苗根系、 莖、 葉柄和葉片的 Fe 含量,分別減少了 43.6%、 21.5%、 5.5% 和 13.7%。添加外源SNP(Cu+SNP)可顯著提高Cu脅迫下番茄幼苗的Fe含量,根系、 莖、 葉柄、 葉片中Fe含量分別比Cu脅迫處理增加了 262.2%、 35.3%、 29.3%和 16.3%,添加血紅蛋白后(Cu+SNP+Hb),番茄幼苗莖和葉柄中的Fe含量比Cu+SNP處理降低了15.6%和7.1%,根系和葉片中Fe含量增加了13.3%和 45.5%,均達到顯著差異水平。
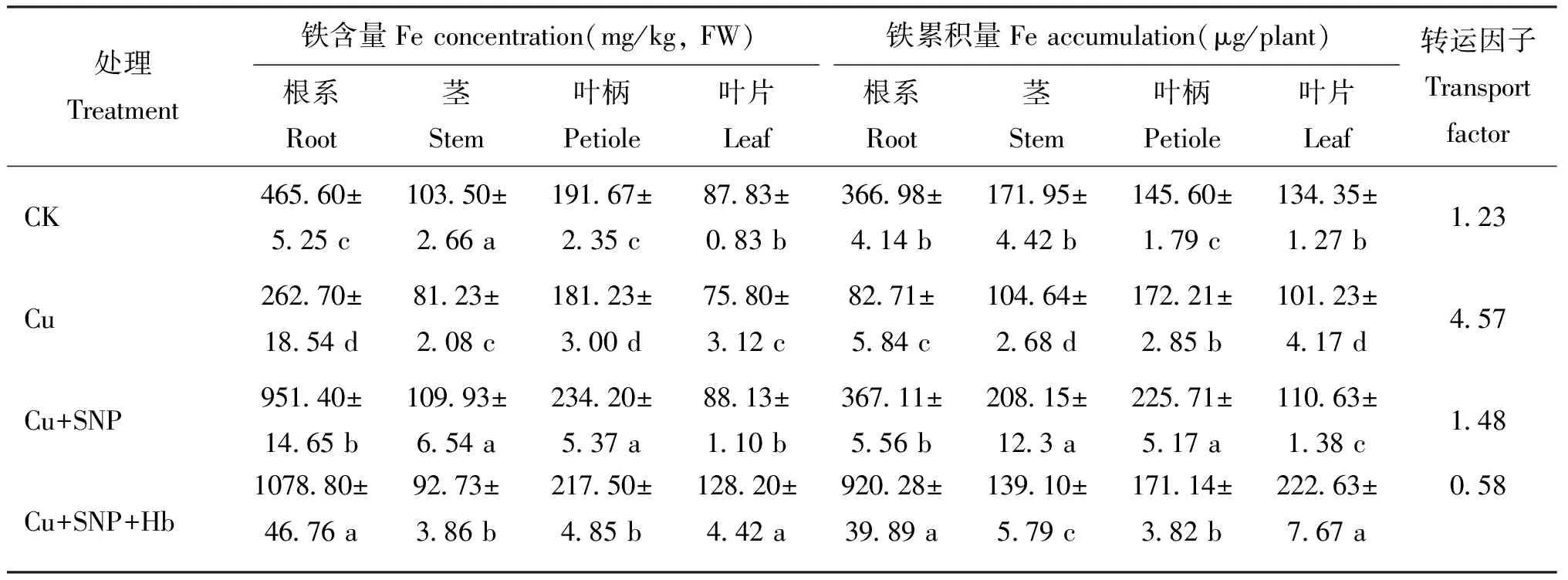
表1 番茄幼苗不同部位的鐵含量和累積量
與 CK 相比,Cu 脅迫處理下,番茄幼苗各部位 Fe 累積量均顯著降低。與 Cu 脅迫處理相比,Cu+SNP 處理顯著提高番茄各部位 Fe 累積量。Cu+SNP+Hb 處理的番茄幼苗莖和葉柄中 Fe 含量顯著降低,根系和葉片中 Fe 含量顯著增加,但仍顯著高于 CK 和 Cu 脅迫處理。
CK 處理的 Fe 轉運因子 (TF) 為 1.23,Cu 脅迫處理 TF 為 4.57,添加外源 SNP 使 TF 降低為 1.48,表明外源 SNP 能顯著降低根系中的 Fe 向地上部轉運。
2.3 外源 NO 對 Cu 脅迫下番茄幼苗 Fe 的亞細胞分布的影響
表 2 顯示,Cu 脅迫處理與對照(CK)相比,番茄幼苗根系細胞中細胞壁的 Fe 含量顯著提高。 添加外源 SNP(Cu+SNP)可顯著提高細胞器和液泡的 Fe 含量,分別比 Cu 脅迫處理提高了 54.5% 和 133.2%,而細胞壁的 Fe 含量顯著降低 27.3%,差異均達顯著水平。添加血紅蛋白后(Cu+SNP+Hb),顯著消除了 NO 的效果,與 Cu+SNP 處理相比,細胞壁和液泡 Fe 含量顯著降低 35.1% 和 39.4%,而細胞器的 Fe 含量顯著上升 21. 4%。表明番茄幼苗根系受到 Cu 脅迫后,外源 NO 可以提高細胞器中的活性鐵,有利于細胞進行正常代謝。
與 CK 相比,Cu 脅迫處理提高了番茄幼苗莖細胞中細胞器和液泡的 Fe 含量。Cu 脅迫處理下,莖細胞中細胞器和液泡的 Fe 含量分別比CK 提高 49.2% 和 16.6%,而細胞壁的 Fe 含量顯著降低 52.9%。添加外源 SNP(Cu+SNP) 后,細胞壁 Fe 含量占莖細胞 Fe 總含量的比例與 Cu 脅迫處理相比升高 10.6%,細胞器 Fe 含量所占比例升高 9.1%, 液泡 Fe 含量所占比例降低 19.7%。添加血紅蛋白后(Cu+SNP+Hb),不同亞細胞的 Fe 含量均顯著降低。
Cu 脅迫處理與 CK 相比,顯著提高番茄幼苗葉柄細胞中細胞壁和液泡的 Fe 含量,分別比CK 提高 13.1% 和 15.3%, 而細胞器的 Fe 含量顯著降低了 19.2%。與 Cu 脅迫處理相比,添加外源 SNP(Cu+SNP)后顯著增加了番茄幼苗根細胞中不同亞細胞的 Fe 含量,細胞壁、 細胞器和液泡的 Fe 含量分別提高了 30.2%、 43.5% 和 18.8%。 添加血紅蛋白后(Cu+SNP+Hb),細胞壁、 細胞器和液泡中的 Fe 含量比 Cu+SNP 處理分別降低了 11.3%、 14.9% 和 19.4%。
Cu 脅迫處理下,番茄幼苗葉片細胞中液泡的 Fe 含量比 CK 提高 8.9%,而細胞壁和細胞器則比 CK 分別降低了 11.6%、 20.5%。與 Cu 脅迫處理相比,Cu+SNP 處理的細胞器和液泡的 Fe 含量分別提高 14.9% 和 15.0% 細胞壁 Fe 含量降低了 24.0%。Cu+SNP+Hb 處理的細胞壁和細胞器的 Fe 含量占葉片細胞 Fe 總含量的比例,與 Cu 脅迫處理相比分別提高了 5.0% 和 4.3%,液泡的 Fe 含量所占比例下降了 9.3%。

表2 番茄幼苗中鐵的亞細胞分布
2.4 外源 NO 對 Cu 脅迫下番茄幼苗 Zn 含量和累積量的影響
由表 3 可以看出,番茄幼苗各部位的 Zn 含量和累積量變化幅度較大,根系和葉片的Zn 含量和累積量最高。不同處理間番茄幼苗各部位的 Zn 含量和累積量差異顯著。Cu 脅迫處理下,根系、 莖、 葉柄的 Zn 含量分別比 CK 減少了 85.1%、 19.8%、 15.0%,而葉片中 Zn 含量增加了 28.4%。Cu+SNP 處理可顯著提高 Cu 脅迫下番茄幼苗根系、 葉柄和葉片的 Zn 含量,分別比 Cu 脅迫處理下提高了 8.3%、 27.9% 和 17.2%,而莖中 Zn 含量降低了 17.1%。Cu+SNP+Hb 處理的番茄幼苗莖、 葉柄和葉片中的 Zn 含量與 Cu+SNP 處理相比降低 18.0%、 7.4%和 21.3%,而根中的 Zn 含量為 Cu+SNP 處理的 3.83 倍,差異顯著。
Cu 脅迫處理下,番茄幼苗根系和莖的 Zn 累積量比 CK 顯著降低,葉柄和葉片中的 Zn 含量則顯著增加。與 Cu 脅迫處理相比,添加外源 SNP 顯著提高了番茄各部位 Zn 的累積量。添加血紅蛋白后,番茄幼苗根系和葉片的 Zn 累積量顯著提高,而莖和葉柄的 Zn 累積量顯著降低。
CK 處理的 Zn 轉運因子(TF)為 0.85,Cu脅迫處理后 TF 為 13.48,添加外源 SNP 使 TF 降低為 11.84,表明外源 SNP 能顯著降低根系中的 Zn 向地上部轉運。
2.5 外源 NO 對 Cu 脅迫下番茄幼苗 Zn 的亞細胞分布的影響
與 CK 相比,Cu 脅迫顯著降低番茄幼苗根系不同亞細胞結構中的 Zn 含量,細胞壁、 細胞器和液泡的 Zn 含量分別降低 23.9%、 75.7%和35.9% (表4)。添加外源 SNP(Cu+SNP)可顯著提高液泡的 Zn 含量,比 Cu 脅迫處理提高了 91.1%,細胞器的 Zn 含量僅提高了 0.7%,而細胞壁的 Zn 含量降低了 31.9%。添加血紅蛋白后(Cu+SNP+Hb),顯著消除了 NO 的效果,與 Cu+SNP 處理相比,細胞壁 Zn 含量顯著上升了 95.7%,液泡的 Zn 含量降低 13.4%.而細胞器的 Zn 含量無明顯變化。

表3 番茄幼苗不同部位的鋅含量和累積量
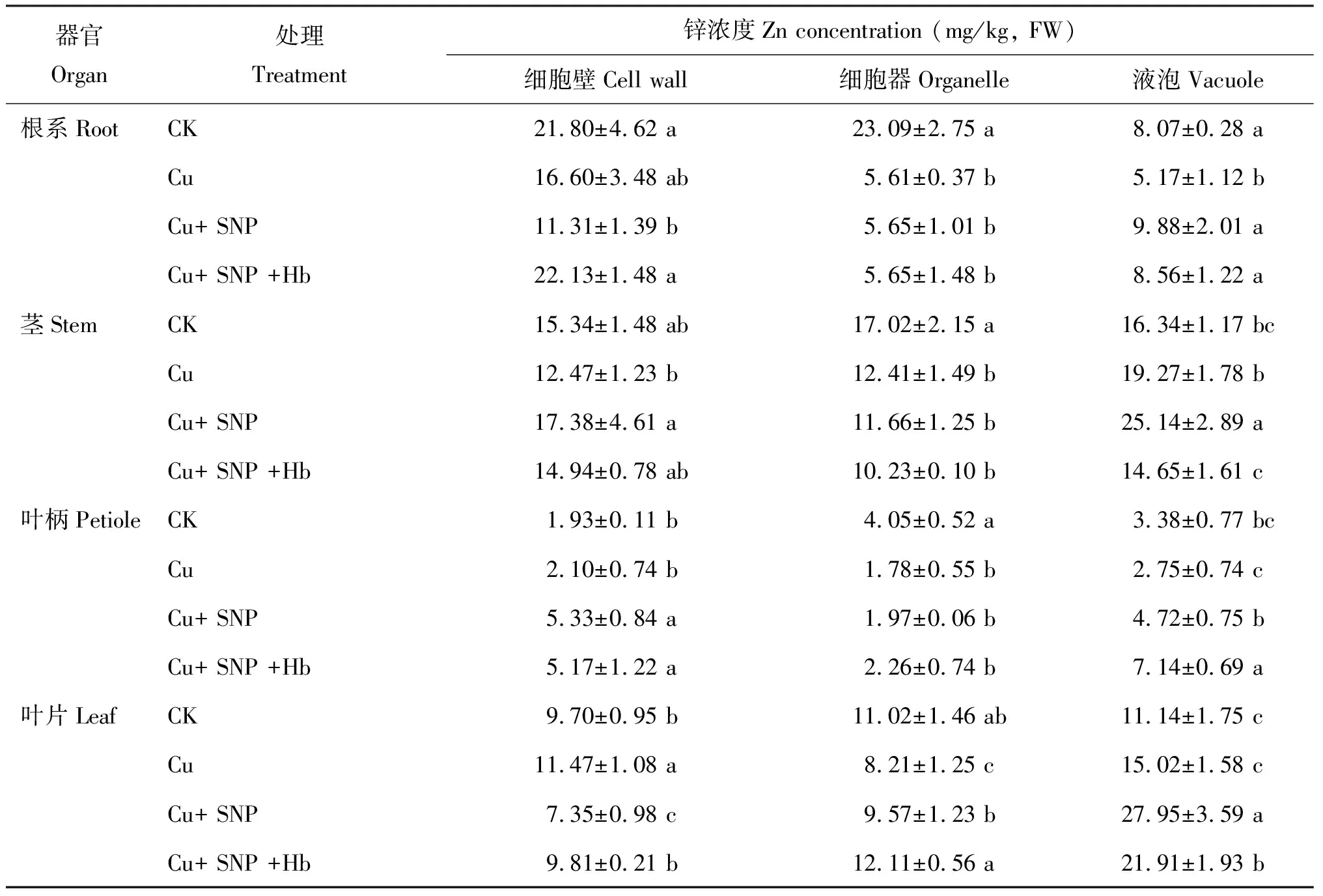
表4 番茄幼苗中鋅的亞細胞分布
Cu 脅迫處理比 CK 番茄幼苗莖細胞中液泡的 Zn 含量有所提高。 Cu 脅迫處理下,番茄幼苗莖細胞中液泡的 Zn 含量比 CK 提高 17.9%,而細胞壁和細胞器 Zn 含量則比 CK 分別降低了 18.7% 和 27.1%。添加外源 SNP 后(Cu+SNP),細胞壁和液泡 Zn 含量占莖細胞 Zn 總含量的比例與 Cu 脅迫處理相比分別升高 3.8% 和 2.8%,細胞器 Zn 含量所占比例降低了 6.6%。添加血紅蛋白后(Cu+SNP+Hb),不同亞細胞的 Zn 含量均顯著降低。
Cu 脅迫處理顯著提高了番茄幼苗葉柄細胞中細胞壁的 Zn 含量,比 CK 提高 8.8%,而細胞器和液泡的 Zn 含量分別降低 56.0%、 18. 6%。與 Cu 脅迫處理相比,添加外源 SNP (Cu+SNP)后顯著增加番茄幼苗根細胞中不同亞細胞結構中的 Zn 含量,細胞壁、 細胞器和液泡的 Zn 含量分別提高了153.8%、 10.7%和71.6%。添加血紅蛋白后,即 Cu+SNP+Hb 處理細胞壁中 Zn 含量比 Cu+SNP 處理降低 3.0%,但差異不顯著,細胞器和液泡 Zn 含量分別提高了14.7% 和 51.3%。
Cu 脅迫處理下,番茄幼苗葉片細胞中細胞壁和液泡的 Zn 含量比 CK 分別提高了18.2% 和 34.8%,而細胞器中 Zn 含量降低 25.5%。添加外源 SNP 后,細胞器和液泡中 Zn 含量比 Cu 脅迫處理分別提高 16.6% 和 86.1%,而細胞壁中 Zn 含量下降 35.9%。添加血紅蛋白后,細胞壁和細胞器的 Zn 含量占葉片細胞 Zn 總含量的比例與 Cu+SNP 處理相比分別提高了 6.0% 和 6.3%,液泡的 Zn 含量所占比例顯著下降,降低幅度為 12.3%。
2.6 外源 NO 對 Cu 脅迫下番茄幼苗 Mn含量及累積量的影響
由表 5 可以看出,番茄幼苗各部位的 Mn 含量和累積量變化幅度較大,番茄幼苗各部位的 Mn 含量和累積量在不同處理間差異顯著。與對照(CK)相比,Cu脅迫處理下,番茄幼苗根系、 葉柄和葉片 Mn 含量分別比對照增加了 15.2%、 20.7%、 29.2%,莖中 Mn 含量減少 39.3%。添加外源 SNP(Cu+SNP)可顯著提高 Cu 脅迫下番茄幼苗根系和莖的 Mn 含量,分別比 Cu 脅迫處理下提高 14.6% 和 40.2%; 但葉柄和葉片 Mn 含量顯著降低了 34.8% 和 30.2%。添加血紅蛋白后(Cu+SNP+Hb),番茄幼苗根系和莖中的 Mn 含量降低了 5.1% 和 18.8%,而葉柄和葉片中的 Mn 含量顯著增加了 72.7% 和27.3%。

表5 番茄幼苗不同部位的錳含量和累積量
與對照相比,Cu 脅迫處理下,番茄幼苗葉柄和葉片的 Mn 累積量顯著增加,根系和莖中的 Mn 累積量顯著降低。與 Cu 脅迫處理相比,添加外源 SNP 顯著提高了番茄根系和莖中 Mn 的累積量,顯著降低葉柄和葉片中 Mn 的累積量。添加血紅蛋白后(Cu+SNP+Hb),番茄幼苗莖中 Mn 累積量顯著降低,根系、 葉柄和葉片中 Mn 累積量顯著增加。
CK 處理的 Mn 轉運因子(TF)為 4.16,Cu 脅迫處理后轉運因子為 6.70,添加外源 SNP 使 TF 降低為 6.01,表明外源 SNP 能顯著降低根系中的 Mn 向地上部轉運。
2.7 外源 NO 對 Cu 脅迫下番茄幼苗 Mn 的亞細胞分布的影響
由表 6 可知,與對照(CK)相比,Cu 脅迫顯著降低番茄幼苗根系細胞中不同亞細胞的 Mn 含量,細胞壁、 細胞器和液泡的 Mn 含量分別降低 65.2%、 47.1%和82.8%。添加外源 SNP(Cu+SNP)可提高細胞器的 Mn 含量,比 Cu 脅迫處理提高 12.7%,而細胞壁和液泡的 Mn 含量分別降低 12.2% 和 6.0%。與 Cu+SNP 處理相比,添加血紅蛋白后(Cu+SNP+Hb),番茄幼苗根系不同亞細胞結構的 Mn 含量顯著提高,細胞壁、 細胞器和液泡的 Mn 含量分別提高了 252.7%、 101.1%和43.6%。

表6 番茄幼苗中錳的亞細胞分布
Cu 脅迫處理下,番茄幼苗莖細胞中細胞壁和細胞器的 Mn 含量比對照分別提高了 6.4% 和 156.0%,而液泡 Mn 含量降低 2.5%。添加外源 SNP 后,細胞壁和液泡 Mn 含量與 Cu 脅迫處理相比差異不顯著,而細胞器 Mn 含量顯著降低 46.2%。添加血紅蛋白后,細胞壁和液泡 Mn 含量比 Cu+SNP 處理分別降低了 15.7% 和 20.5%,細胞器 Mn 含量提高 9.4%。
Cu 脅迫處理顯著提高了番茄幼苗葉柄細胞中細胞壁和液泡的 Mn 含量,分別比對照提高 27.2% 和 66.8%,而細胞器的 Mn 含量顯著降低 22.2%。與 Cu 脅迫處理相比,添加外源 SNP 后顯著增加番茄幼苗根細胞中不同亞細胞結構的 Mn 含量,細胞壁、 細胞器和液泡 Mn 含量分別提高 18.0%、 2.7%和27.3%。添加血紅蛋白后,細胞壁和液泡 Mn 含量比 Cu+SNP 處理分別降低了 41.3% 和 15.8%,細胞器 Mn 含量升高 12.3%。
Cu 脅迫下,番茄幼苗葉片細胞壁、 細胞器和液泡的 Mn 含量分別比對照提高 50.8%、 69.8%和2.8%。添加外源 SNP 后,液泡中 Mn 含量比 Cu 脅迫處理提高 11.4%,而細胞壁和細胞器 Mn 含量分別下降 43.2% 和 35.1%。添加血紅蛋白后,細胞器的 Mn 含量占葉片細胞 Mn 總含量的比例與 Cu+SNP 處理相比提高了 6.3%,細胞壁和液泡的 Mn 含量所占比例分別下降了 6.0% 和 0.3%。
3 討論與結論
鐵(Fe)是生物體生命活動中必需的微量元素之一,其參與生物體內呼吸作用、 光合作用、 DNA 合成、 氮素同化和固定、 激素合成、 活性氧的形成與消除等重要的生理代謝過程[15]。植物正常生長發育不僅需要從環境中吸收充足的鐵,更需要其在植物體內高效運轉和在器官中合理分布以保證完成生命周期[16]。本試驗中,銅(Cu) 脅迫使番茄幼苗各部位的 Fe 含量和累積量顯著降低。這可能是由于過量 Cu2+存在競爭性抑制根系對 Fe2+的吸收,或者根系積累的大量 Cu2+限制了 Fe2+向地上部運輸。添加外源 SNP 可顯著增加番茄各部位的 Fe 含量,其中根系和葉柄中 Fe 含量和累積量最高。而添加血紅蛋白可使 NO 的緩解效果消除。這可能是因為 SNP 釋放出 NO,調控質膜上 ATPase 等功能蛋白活性,提高了 Cu 脅迫下番茄根系對 K、 Ca、 Fe、 Zn 等離子的吸收,使其在器官間的分配更加趨于合理,以維持胞質的離子穩態,從而緩解過多銅的毒害[17]。
研究表明,鋅(Zn)在玉米細胞內的分布特征與其吸收和富集重金屬能力密切相關[19]。細胞壁固持和液泡區隔化在植物對重金屬的解毒、 耐性和超富集方面起著重要作用[20]。本試驗中,Cu 脅迫使番茄幼苗根系 Zn 含量和累積量顯著降低,葉片 Zn 含量和累積量顯著提高。添加外源 SNP 可顯著促進根系 Zn 的吸收和累積,提高葉片 Zn 含量和累積量。而添加血紅蛋白可使 NO 的這種作用被削弱。說明正是由于 SNP 釋放的 NO 作用于根系細胞質膜的磷脂雙分子層,維持質膜正常的離子選擇性,促進根系對 Zn 的吸收,緩解質膜受過多 Cu 的毒害[21-22]。另外,NO 作為信號分子參與抗逆信號轉導,保護植物細胞免受逆境脅迫的損傷,提高根系和葉片細胞對 Zn 的吸收和富集[23]。
鋅(Zn)在植物細胞的不同部位和細胞器中含量分布不一,胞內區室化分隔有利于保證缺鋅和高鋅脅迫下植物維持正常的生理代謝[24]。Ma等[25]研究顯示,在天藍遏藍菜(Thlaspicaerulescens)的葉片中,77% 的鋅主要儲存在原生質體,其中 91% 的鋅出現在液泡中。本試驗中,鋅主要集中在根系和莖的亞細胞結構中,少量鋅存在于葉片和葉柄中。Cu 脅迫處理后,Zn 的亞細胞分布受到 Cu 脅迫處理的影響,根系各亞細胞結構和各部位細胞器中 Zn 含量顯著降低,而葉片和葉柄細胞壁中 Zn 含量小幅提高,另有部分 Zn 含于莖和葉片的液泡中。添加 SNP 可顯著增加各部位液泡、 細胞器的 Zn 含量,降低根系和葉片細胞壁 Zn 含量。表明外源 NO 能促進 Zn 在番茄體內的吸收和分布,并且影響 Zn 在細胞器和液泡內的積累,提高各部位細胞器對 Zn 的需求以及液泡中有機酸、 氨基酸和花青素等對過量 Zn 的儲存[26]。
作為植物生命活動所必需的微量元素之一,錳(Mn)過量或缺乏,導致葉綠素合成減少、 光合作用受阻、 活性氧累積和體內激素失衡,導致生長受阻,尤其是葉部和根尖分生區的生長受到明顯的抑制[27]。Xu等[28]利用同步輻射X-射線熒光光譜(synchrotron radiation X-ray fluorescence,SRXRF)微探針對商陸(PhytolaccaacinosaRoxb)根系、 莖和葉片中錳分布的分析結果表明,葉片是錳累積的主要器官,而根系、 莖和葉片的維管組織中存在高濃度的錳,有利于其長距離運輸和向葉片不同組織中分配。根系主要通過有機酸的螯合作用促進植物對 Mn2+的轉運解毒,同時能夠將過量的 Mn2+分隔在細胞壁中; 葉片可通過酚類物質或有機酸螯合 Mn2+,并將其分隔在葉片表皮細胞和葉肉細胞的液泡中[29]。本試驗中,Cu 脅迫使番茄幼苗葉柄和葉片的 Mn 含量和累積量顯著提高,莖中 Mn 含量和累積量顯著降低,根系 Mn 含量顯著升高。添加 SNP 可顯著增加根系和莖中 Mn 含量和累積量,降低葉柄和葉片中 Mn 含量和累積量。添加血紅蛋白后 NO 的這種作用被減弱。表明銅脅迫下,外源 NO 可能介導了抗氧化酶保護系統,清除根尖細胞活性氧(ROS)引起的氧化損傷,保護細胞膜免受重金屬脅迫的傷害,增強根系中有機酸(尤其是草酸、 檸檬酸)對過量 Mn2+的螯合,同時,促進葉柄和葉片中 Mn 向莖中轉運,提高莖中 Mn 的補給,緩解 Cu 毒害[30-31]。
從番茄幼苗各部位及亞細胞中Fe、 Zn、 Mn含量和累積量的分布來看,同正常營養液處理相比,外源NO在Cu脅迫逆境下可以顯著提高Fe、 Zn、 Mn的選擇性吸收,調控番茄幼苗各部位及亞細胞中Fe、 Zn、 Mn的合理分布,維持胞質離子穩態和礦質營養元素平衡,緩解銅脅迫,保證番茄幼苗正常的生理代謝。
參考文獻:
[1] 王曉蘭, 周守標, 楊集輝, 等. 銅脅迫對彎囊苔草(Carexdispalata)生長和生理特性的影響[J]. 農業環境科學學報, 2010, 29(2): 264-269.
Wang X L, Zhou S B, Yang J Hetal. Copper stress on the growth and some physiological parameters ofCarexdispalata[J]. Journal of Agro-Environment Science, 2010, 29(2): 264-269.
[2] 謝建春, 趙娟, 楊世勇. 銅尾礦對油菜生長和生理功能的影響[J]. 生態與農村環境學報, 2009, 25(2): 74-79.
Xie J C, Zhao J, Yang S Y. Effects of copper mine tailings on growth and physiological functions ofBrassicacampestris[J]. Journal of Ecology and Rural Environment, 2009, 25(2): 74-79.
[3] Sharma S S, Dietz K J. The relationship between metal toxicity and cellular redox imbalance[J]. Trends in Plant Science, 2009, 14(1): 43-50.
[4] Wojcik M, Tukiendorf A. Response of wild type ofArabidopsisthalianato copper stress[J]. Biologia Plantarum, 2003, 46(1): 79-84.
[5] 高曉寧. 土壤重金屬污染現狀及修復技術研究進展[J]. 現代農業科技, 2013, (9): 229-231.
Gao X N. Research advances on soil heavy metal pollution and remediation technology[J]. Modern Agricultural Science and Technology, 2013, (9): 229-231.
[6] Beligni M V, Lamattina L. Nitric oxide: a non-traditional regulator of plant growth[J]. Trends in Plant Science, 2001, 6(11): 508-509.
[7] Tan Y W, Xu Y F, Zhou H. Effects of nitric oxide on seed germination and seedling growth of tall fescue under salt stress[J]. Acta Agrestia Sinica, 2010, 18(3): 394-398.
[8] 張敏, 姜春輝, 李曉云, 崔秀敏. 外源 NO 對銅脅迫下番茄幼苗根系構型及其超微結構的影響[J]. 植物生理學報, 2012, 48(11): 1105-1112.
Zhang M, Jiang C H, Li X Y, Cui X M. Effects of exogenous nitric oxide on tomato seedlings root architecture and ultrastructure under copper stress[J]. Plant Physiology Journal, 2012, 48(11): 1105-1112.
[9] 張敏, 姜春輝, 崔秀敏. 外源 NO 供體硝普鈉(SNP)對銅脅迫下番茄幼苗生理生化指標的影響[J]. 植物生理學報, 2013, 49(2): 144-152.
Zhang M, Jiang C H, Cui X M. Effects of exogenous nitric oxide donor SNP on physiological and biochemical indexes in tomato seedlings under copper stress[J]. Plant Physiology Journal, 2013, 49(2): 144-152.
[10] 李興華, 方子森, 牛俊義. 大量及微量元素對胡麻幼苗生長發育的影響[J]. 甘肅農業大學學報, 2013, 48(1): 42-48.
Li X H, Fang Z S, Niu J Y. Effects of major and trace elements on growth and development of flax seedlings[J]. Journal of Gansu Agricultural University, 2013, 48(1): 42-48.
[11] 姜春輝, 王秀峰, 尹博, 等. 外源 NO 對 Cu 脅迫下番茄 Cu 的亞細胞分布和化學形態的影響[J]. 應用生態學報, 2012, 23(11): 3033-3039.
Jang C H, Wang X F, Yin Betal. Effects of exogenous nitric oxide on the subcellular distribution and chemical forms of copper in tomato seedlings under copper stress[J]. Chinese Journal of Applied Ecology, 2012, 23(11): 3033-3039.
[12] Weigel H J, J?ger H J. Subcellular distribution and chemical form of cadmium in bean plant[J]. Plant Physiology, 1980, 65: 480-482.
[13] Rathore V S, Baiai Y P, Wittwer S H. Subcellular localization of zinc and calcium in bean (PhaseolusvulgarisL.) tissues[J]. Plant Physiology, 1972, 49: 207-211.
[14] 段曼莉, 付冬冬, 王松山, 等. 亞硒酸鹽對四種蔬菜生長、 吸收及轉運硒的影響[J]. 環境科學學報, 2011, 31(3): 658-665.
Duan M L, Fu D D, Wang S Setal. Effects of different selenite concentrations on plant growth, absorption and transportation of selenium in four different vegetables[J]. Acta Scientiae Circumstantiae, 2011, 31(3): 658-665.
[15] 申紅蕓, 熊宏春, 郭笑彤, 左元梅. 植物吸收和轉運鐵的分子生理機制研究進展[J]. 植物營養與肥料學報, 2011, 17(6): 1522-1530.
Sheng H Y, Xiong H C, Guo X T, Zuo Y M. Progress of molecular and physiological mechanism of iron uptake and translocation in plants[J]. Plant Nutrition and Fertilizer Science, 2011, 17(6): 1522-1530.
[16] Morrissey J, Guerinot M L. Iron uptake and transport in plants: the good, the bad, and the ionome[J]. Chemical Reviews, 2009, 109(10): 4553-4567.
[17] Zhang Y K, Han X J, Chen X Letal.Exogenous nitric oxide on antioxidative system and ATPase activities from tomato seedlings under copper[J]. Scientia Horticulturae, 2009, 123(2): 217-223.
[18] Graziano M, Beligni M V, Lamattina L. Nitric oxide improves internal iron availability in plants[J]. Plant Physiology, 2002, 130: 1852-1859.
[19] 周建民, 黨志, 陶雪琴,周永章. NTA 對玉米體內 Cu、 Zn 的積累及亞細胞分布的影響[J]. 環境科學, 2005, 26(6): 126-130.
Zhou J M, Dang Z, Tao X Q, Zhou Y Z. Influence of NTA on accumulation and subcellular distribution of copper and zinc in corn (Zeamays)[J]. Environmental Science, 2005, 26(6): 126-130.
[20] Küpper H, Lombi E, Zhao F Jetal. Cellular compartmentation of cadmium and zinc in relation to other elements in the hyperaccumulatorArabidopsishalleri[J]. Planta, 2000, 212(1): 75-84.
[21] 張遠兵, 劉愛榮, 方蓉. 外源一氧化氮對鎘脅迫下黑麥草生長和抗氧化酶活性的影響[J]. 草業學報, 2008, 17(4): 57-64.
Zhang Y B, Liu A R, Fang R. Effect of exogenous nitric oxide on the growth and antioxidant enzyme activities of Lolium perenne under Cd stress[J]. Acta Prataculturae Sinica, 2008, 17(4): 57-64.
[22] 張義凱, 韓小嬌, 金洪, 等. 外源 NO 對銅脅迫下番茄光合、 生物發光特性及礦質元素吸收的影響[J]. 植物營養與肥料學報, 2010, 16(1): 172-178.
Zhang Y K, Han X J, Jin Hetal. Effects of exogenous nitric oxide on photosynthetic, bioluminescent characteristics and mineral element contents in tomato under copper stress[J]. Plant Nutrition and Fertilizer Science, 2010, 16(1): 172-178.
[23] 彭金英, 黃勇平. 植物防御反應的兩種信號轉導途徑及其相互作用[J]. 植物生理與分子生物學報, 2005, 31(4): 347-353.
Peng J Y, Huang Y P. The signaling pathways of plant defense response and their interaction[J]. Journal of Plant Physiology and Molecular Biology, 2005, 31(4): 347-353.
[24] Broadley R, White P J, Hammond J Petal. Zinc in plants[J]. New Phytologist, 2007, 173(4): 677-702.
[25] Ma J F, Ueno D, Zhao F Jetal. Subcellular localization of Cd and Zn in the leaves of a Cd-hyperaccumulating ecotype ofThlaspicaerulescens[J]. Planta, 2005, 220(5): 731-736.
[26] Cosio C, Martinoia E, Keller C. Hyperaccumulation of cadmium and zinc inThlaspicaerulescensandArabidopsishalleriat the leaf cellular level[J]. Plant Physiology, 2004, 134(2): 716-725.
[27] Santandrea G, Schiff S, Bennici A. Effects of manganese on Nicotiana species cultivated in vitro and characterization of regenerated Mn-tolerant tobacco plants[J]. Plant Science, 1998, 132(1): 71-82.
[28] Xu X H, Shi J Y, Chen Y Xetal. Distribution and mobility of manganese in the hyperaccumulator plantPhytolaccaacinosaRoxb. (Phytolaccaceae)[J]. Plant and Soil, 2006, 285(1-2): 323-331.
[29] 張玉秀, 李林峰, 柴團耀, 等. 錳對植物毒害及植物耐錳機理研究進展[J]. 植物學通報, 2010, 45(4): 506-520.
Zhang Y X, Li L F, Chai T Yetal. Mechanisms of manganese toxicity and manganese tolerance in plants[J]. Chinese Bulletin of Botany, 2010, 45(4): 506-520.
[30] Mora M L, Rosas A, Ribera Aetal. Differential tolerance to Mn toxicity in perennial ryegrass genotypes: involvement of antioxidative enzymes and root exudation of carboxylates[J]. Plant and Soil, 2009, 32(1-2): 79-89.
[31] 趙寶泉, 萬宇, 楊世湖, 等. 外源 NO 供體硝普鈉(SNP)對重金屬 Cd 脅迫下水稻幼苗膜脂過氧化及抗氧化酶的影響[J]. 江蘇農業學報, 2010, 26(3): 468-475.
Zhao B Q, Wan Y, Yang S Hetal. Effects of exogenous nitric oxide donor SNP on membrane lipid peroxidation and activity of antioxidative enzyme in rice seedlings under Cd stress[J]. Jiangsu Journal of Agricultural Sciences, 2010, 26(3): 468-475.
[32] 徐向華, 施積炎, 陳新才, 等. 錳在商陸葉片的細胞分布及化學形態分析[J]. 農業環境科學學報, 2008, 27(2): 515-520.
Xu X H, Shi J Y, Chen X Cetal. Subcellular distribution and chemical fractions of manganese in leaves of hyper accumulator phytolacca acinosa roxb[J]. Journal of Agro-Environment Science, 2008, 27(2): 515-520.
[33] 倪才英, 劉永厚, 趙振紀. 贛東北土壤中銅的毒害作用研究[J]. 江西師范大學學報(自然科學版), 1999, 23(2): 170-173.
Ni C Y, Liu Y H, Zhao Z J. A study on toxicity of copper in soil of northeast Jiangxi province[J]. Journal of Jiangxi Normal University(Natural Sciences Edition), 1999, 23(2): 170-173.
