不同施氮水平對煙富3/M26/平邑甜茶幼樹當年及翌年氮素吸收、 利用、 分配的影響
2014-03-30 06:13:46姜遠茂魏紹沖王富林李洪娜
植物營養與肥料學報 2014年2期
關鍵詞:生長
李 晶, 姜遠茂, 魏紹沖, 王富林, 周 樂, 李洪娜
(山東農業大學園藝科學與工程學院,作物生物學國家重點實驗室,山東泰安 271018)
蘋果矮化密植栽培的矮化性、 一致性、 早果性、 安全化、 機械化等優點[1]使蘋果矮化密植栽培成為現代蘋果產業的主要發展趨勢,目前主要有自根砧和中間砧兩種應用形式。我國65%的果樹種植在土壤瘠薄的丘陵地[2],土壤有機質普遍較低[3],短期內全面提高較困難,決定了對肥水條件要求較高的矮化自根砧適宜栽植區域有限,這是目前我國蘋果矮化密植仍以矮化中間砧為主的主要原因。氮作為果樹生長所必需的礦質元素之一,氮素營養的吸收、 利用和分配直接或間接影響樹體的器官分化、 形成及建造,與樹體的生長發育、 果實產量和品質形成密切相關。目前蘋果栽培過程中偏施氮肥,施肥不足和超量并存,肥料利用率低,施肥時期不合理,后期脫肥,貯藏營養不足等問題嚴重[2]。根系作為果樹養分和水分的主要吸收器官,植物通過根系形態與生理學反應來適應土壤環境中氮素供應強度的變化[4],作物在生長前期較強的根系生長勢利于快速建立起較大的根系以提高氮素吸收利用效率[5],進而促進作物生長,協調的根系與地上部生長是保障果樹對養分高效吸收,構建豐產、 穩產、 優質樹體結構的根本措施[2]。因此,通過研究施氮不足、 適量施氮肥和過量施氮肥對蘋果矮化中間砧幼樹的生長及氮素營養吸收、 利用、 分配規律的影響,可明確不同氮肥施用量栽培條件下不同生長階段樹體各器官對氮的需求及利用特點,以采取相應措施對樹體各器官的生長節奏加以調節,協調樹體根系與地上部的生長以促進幼樹快速生長縮短營養期,協調營養生長與生殖生長以早果穩產豐產。因此,通過研究不同施氮量對蘋果幼樹矮化中間砧的生長及氮素營養吸收、 利用、 分配規律的影響,可為蘋果矮化中間砧高產高效栽培配套技術提供理論依據。
1 試驗材料與方法
1.1 試材與處理
試驗于2011年在山東農業大學園藝試驗站進行。試材為定植在園藝試驗站的2年生煙富3/M26/平邑甜茶幼樹,不同處理間設置隔離樹。供試土壤為粘壤土,有機質10.13 g/kg,堿解氮76.63 mg/kg,速效磷27.28 mg/kg,速效鉀184.99 mg/kg,硝態氮37.95 mg/kg,銨態氮16.17 mg/kg,pH 6.7。于2011年4月15日各選取9株生長勢基本一致、 無病蟲害的植株,試驗設氮肥施用量: 0 kg/hm2、 100 kg/hm2、 200 kg/hm2(以下簡稱N0、 N100、 N200)三個水平。供試氮肥為尿素(46%),每株幼樹土施1 g15N-尿素(上海化工研究院生產,豐度10.22%),過磷酸鈣43.13 g、 硫酸鉀12.7 g。N0不另施氮肥,N100、 N200分別施用尿素15 g、 30 g,至翌年春梢旺長期不再施肥。施肥后立即澆水,進行常規管理,各處理生長條件和其他栽培管理保持一致。
1.2 取樣與測定
分別于2011年6月1日(春梢旺長期)、 7月1日(春梢停長期)、 8月15日(秋梢旺長期)和2012年6月10日(翌年春梢旺長期)破壞性采樣。每次取樣各處理均取3株。整株解析為葉、 一年生枝條、 接穗韌皮部、 接穗木質部、 中間砧韌皮部、 中間砧木質部、 根砧韌皮部、 根砧木質部。樣品按清水→洗滌劑→清水→1%鹽酸→3次去離子水順序沖洗后,80℃下烘干至恒重,粉碎后過0.25 mm目篩,混勻后裝袋備用。15N豐度在DELTAplusXP型質譜儀中測定。
1.3 計算及統計方法
計算公式如下:
Ndff% =[樣品中的15N豐度%-自然豐度(0.365%)]/[肥料中15N的豐度%-自然豐度(0.365%)]×100;
總N量(g)=干物重(g)×N%;
15N吸收量(mg)=總N量(g)×Ndff%×1000;
氮肥分配率(%)=[各器官從15N吸收N量(mg)/15N總吸收N量(mg)]×100;
氮肥利用率(%)=15N吸收量(g)/施氮量(g)×100。
所有數據均采用DPS7.05軟件進行單因素方差法分析,LSD法進行差異顯著性比較,應用Microsoft Excel 2003進行圖表繪制。
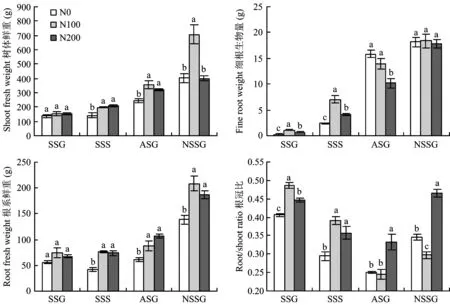
圖1 施氮水平對樹體生長的影響Fig.1 Effects of different N rates on growth of apple tree [注(Note): SSC—春梢生長期 Spring shoot growing; SSS—春梢停止生長期Spring shoot stop growing;ASG—秋梢旺長期Autumn shoot growing; NSSG—翌年春梢生長期Next spring shoot growing. 圖中不同字母表示同一時期處理間差異達到5%顯著水平 Different letters mean significant difference at the same stage at the 5% level.]
2 結果與分析
2.1 施氮水平對樹體生長的影響
施氮水平對樹體生長影響顯著。圖1可見,在春梢旺長期,隨著施氮肥量增加樹體生物量有增加趨勢,但差異不顯著;隨著生長期的延長至春梢停長期,N100和N200處理對樹體生長的促進作用逐漸顯著,N100處理(196.23 g)和N200處理(209.82 g)樹體生物量比N0處理(151.63 g)顯著提高了25.17%和33.84%,表明N100和N200施氮水平均對樹體生長有顯著促進作用;至秋梢旺長期,N100處理(351.38 g)和N200處理(322.94 g)仍保持較高樹體生物量,比N0處理分別提高了44.00%和32.34%,且N100比N200處理對樹體生長有明顯促進作用;至翌年春梢旺長期,N100樹體生物量升高至N0的175.83%和N200的176.41%,N0和N200樹體生物量無顯著差異,表明對樹體生長抑制作用較強的M26中間砧蘋果幼樹的生長沒有隨著施氮水平增加而提高,而是在適宜氮肥水平下表現出較大生物量,這一結果在年周期中是逐漸顯現出來,且適宜施氮對樹體翌年生長的促進作用較當年更為顯著,與宇萬太[6]和韓振海[7]等的研究結果一致。 在春梢旺長期和春梢停長期,N100處理細根生長量最高,其次是N200處理,N0處理最低,三者之間差異顯著,表明氮肥施入在較長時間內對細根生長量的增加有一定促進作用,且N100施氮水平相對于N200處理更利于細根生長。至秋梢旺長期,N0處理細根生物量迅速升高至最高且顯著高于N200處理,與N100處理無顯著差異,這與長期低氮條件下蘋果幼樹產生低氮脅迫響應,誘導細根大量發生有關。N200處理細根生長呈現先高后低趨勢,與羅雪華[8]等人在巴西橡膠樹上的研究結果一致。已往研究發現低氮條件下植物可刺激根的伸長[9],高氮抑制細根生長[10, 11],從試驗結果來看,到秋梢旺長期才在細根數量上與這個結論吻合,表明蘋果作為多年生植物由于貯藏營養等的影響,施氮水平對根系生長的影響有一定滯后性和特殊性。
氮肥施入初期,即春梢旺長期施氮水平對根系總量的影響由高到低表現為: N100>N200>N0,但差異不顯著。隨著氮肥施入時間和生長期的延長,施氮對根系總量的影響逐漸顯著,至春梢停長期N100和N200處理根系鮮重均顯著高于N0,N100和N200處理無顯著差異,而翌年以適宜施氮處理最高,表明適宜施氮肥對根系總量有促進作用。
在春梢旺長期,N100和N200處理根冠比升高至N0處理的119.04%和119.66%;至春梢停長期,各處理根冠比逐漸降低,但N100和N200施氮處理根冠比仍顯著高于N0,表明適宜施氮肥N100和過量施氮肥N200處理在前期對根冠比有顯著的促進作用;至秋梢旺長期,隨著N100處理地上部生物量的顯著升高,根冠比持續降低至與N0無顯著差異,N200處理仍保持較高根冠比;至翌年春梢旺長期,各施氮處理樹體根冠比均顯著高于其秋梢旺長期,且N0、 N100和N200根冠比分別升高至秋梢旺長期的137.62%、 121.22%和139.71%,由高到低為N200>N0>N100,與秋梢旺長期變化趨勢一致。
2.2 施氮水平對15N利用率的影響
圖2可見,春梢旺長期至秋梢旺長期,不同施氮水平樹體15N利用率均隨著施肥時間的延長而提高,與趙林[12]等人的研究結果認為春季土施氮肥利用率隨物候期的推移而提高的結論一致。不同施氮水平對樹體在不同生長期15N利用率的影響存在顯著差異,春梢旺長期N200和N100處理15N利用率差異不顯著,由高到低為N0(1.29%)>N100(0.34%)>N200(0.25%),與巨曉堂[13]、 鐘茜[14]等人的研究認為氮肥利用率隨著氮肥施用量的增加而降低的結論一致。隨著氮肥施入時間和生長期的延長,春梢停長期至秋梢旺長期N100處理15N利用率均顯著高于N0和N200,氮肥利用率并未隨著氮肥施用量的增加呈現直線下降的趨勢,與劉新宇[15]等在冬小麥上的研究結果一致。至翌年春梢旺長期,各處理15N利用率由高到低仍表現為 N100>N0>N200,表明氮肥適量施用不僅利于樹體當年氮肥利用率的提高,更利于翌年氮肥再利用率的提高。
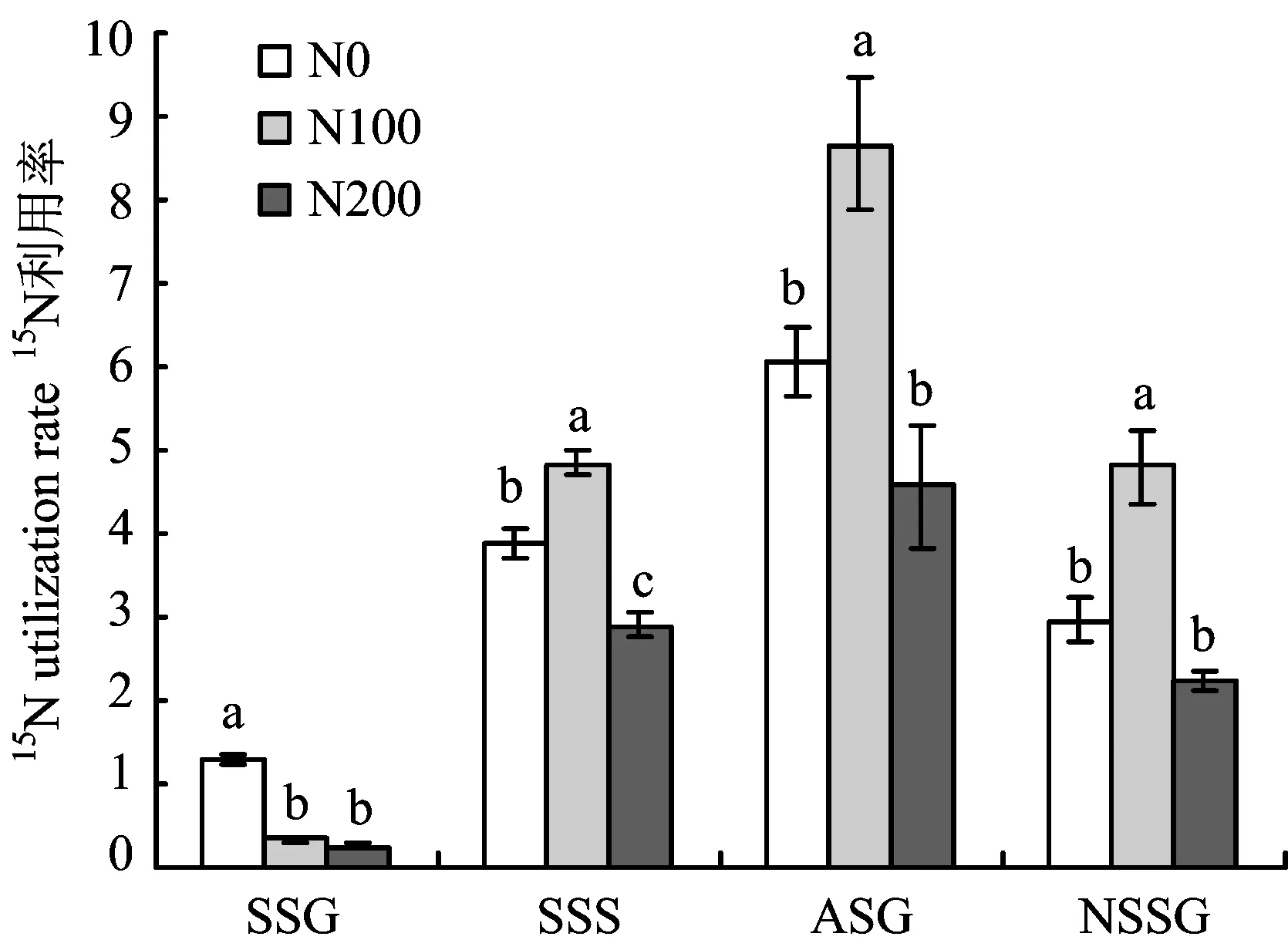
圖2 施氮水平對不同生長期15N利用率的影響Fig.2 Effects of different N rates on15N utilization rate[注(Note): SSC—春梢生長期Spring shoot growing; SSS—春梢停止生長期Spring shoot stop growing;ASG—秋梢旺長期Autumn shoot growing; NSSG—翌年春梢生長期Next spring shoot growing. 圖中不同字母表示同一時期處理間差異達到5%顯著水平Different letters mean significant difference at the same stage at the 5% level.]
2.3 施氮水平對15N分配率的影響
器官中15N占全株15N總量的百分率反映了肥料在樹體內的分布及在各器官遷移的規律[16]。本次試驗結果表明,不同氮肥施用量條件下蘋果幼樹均表現為春梢旺長期至秋梢旺長期地上部15N分配率的逐漸升高和根系15N分配率的逐漸降低,但各物候期不同處理的葉片和根系15N分配率均存在顯著差異,與趙營[17]等認為不同施氮量只影響夏玉米不同生育時期養分的階段積累量,但對累積趨勢基本無影響的結論一致。
氮肥施入至春梢旺長期,葉片15N分配率由高到低為: N0>N200>N100,根系15N分配率由高到低為: N100>N200>N0。不施氮肥N0處理樹體葉片15N分配率為根系15N分配率的113.49%,葉片為最大的15N利用器官;適宜施氮肥N100處理樹體葉片15N分配率為根系15N分配率的為49.15%,N200樹體為74.37%,最大的15N利用器官為根系,表明低氮條件下樹體新吸收的氮素營養可更為快速的運轉至地上部用于新生葉片的構建,而N100和N200施氮條件下,氮肥施入初期樹體生長中心為根系,新吸收的氮素營養更多分配至根系,用于根系生長和根冠比的提高。春梢停長期各施氮處理葉片均成為樹體最大的15N利用器官,根系15N分配率顯著降低,表明至春梢停長期,樹體的生長中心均轉移至地上部,吸收的氮素營養多轉運至葉片用于新生器官建造。根系15N分配率由高到低分別為: N100>N200>N0,表明N100和N200樹體根系仍為較強的生長中心,且N100處理對根系生長的促進作用仍顯著高于N200。至秋梢旺長期,N200樹體葉片15N分配率顯著低于N0和N100,根系15N分配率顯著高于N0和N100,表明N200施氮水平對根系生長的促進作用較N100處理更為顯著,N100和N0樹體各器官15N分配率未表現出顯著差異。至翌年春梢旺長期,不同施氮處理樹體的葉片均為最大的氮利用器官,其次為根系和枝條。N200樹體葉片15N分配率低于N100和N0,根系15N分配率由高到低為: N200>N0> N100,與根冠比變化規律一致,表明至樹體氮素營養在翌年各器官的再利用與分配亦表現為隨著生長中心的轉移而轉移。各處理翌年春梢旺長期葉片15N分配率均顯著高于當年春梢旺長期葉片15N分配率的結果表明,回流的氮素營養相對于當年吸收的氮素營養更優先用于春梢葉片生長,與曾驤[18]的研究認為回流的15N在翌年優先用于葉、 花和新梢生長的結論一致。
不同施氮水平對樹體在氮肥施入當年和翌年不同生長期各器官的15N分配率變化規律的影響表明,施氮水平只改變樹體各器官的15N分配率大小,對不同生長期15N在各器官的分配趨勢無顯著影響,均表現為隨著生長點的轉移而轉移,隨著生長期由春梢旺長期延長至秋梢旺長期,地上部生長勢逐漸增強,地上部15N分配率逐漸提高,根系15N分配率逐漸降低,翌年春梢旺長期根冠比顯著高于氮肥施入當年秋梢旺長期。
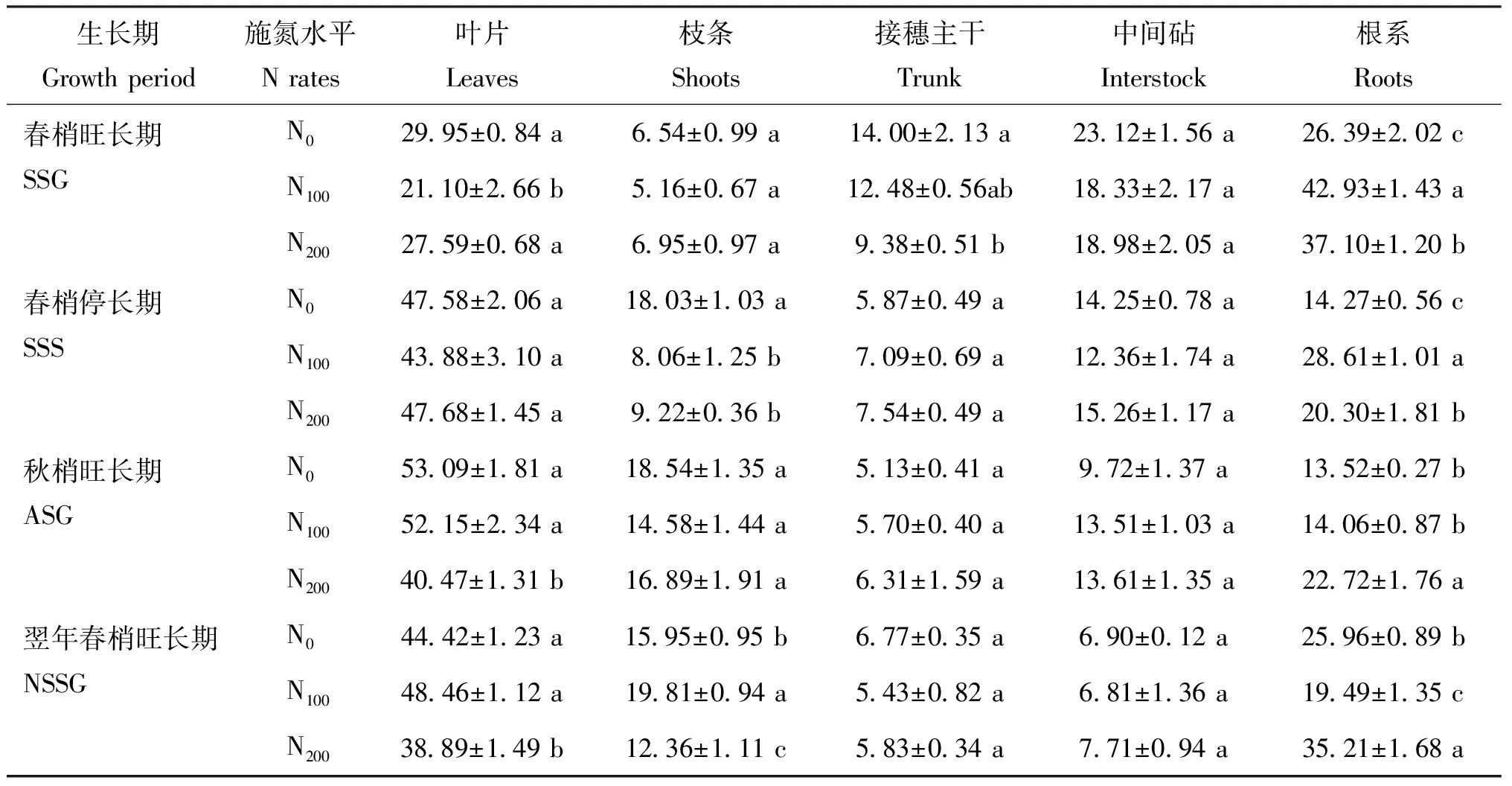
表1 施氮水平對不同生長期各器官15N分配率的影響Table 1 Effects of different N rates on ratio of15N-urea partition of different organs
注(Note): SSC—春梢生長期Spring shoot growing; SSS—春梢停止生長期Spring shoot stop growing; ASG—秋梢旺長期Autumn shoot growing; NSSG—翌年春梢生長期Next spring shoot growing. 不同小寫字母表示同一列中不同施氮處理在同一時期處理間差異達到5%顯著水平Different letters mean significant difference at the same stage at the 5% level.
3 討論
氮肥施入至春梢旺長期適宜施氮肥N100處理和過量施氮肥N200處理根冠比顯著升高,樹體生物量有增加趨勢但未表現出顯著差異,隨著氮肥施入時間的延長,至春梢停長期施氮對樹體生長的促進作用逐漸顯著,至翌年春梢旺長期方表現為適宜施氮肥對樹體的促進作用較過量施氮肥更為顯著,表明施氮水平對蘋果矮化中間砧幼樹生長的影響有一定滯后性,可能與蘋果中間砧樹體作為多年生植物具有貯藏營養可循環供給特性和春梢旺長期新生器官的生長主要依靠貯藏營養有關,還可能與植物氮吸收系統對氮脅迫的主要適應性響應為根冠比變化,在不同氮水平下首先通過調節根系生長及其生理學反應(如養分轉運蛋白的表達及活性等)去應對環境養分供應強度的變化[4],進而促進地上部生長有關。
對樹體生長抑制作用較強的M26中間砧蘋果幼樹的生長并不隨著施氮水平增加而提高,只在適宜施氮水平下表現出較大生物量和較高氮肥利用率可能與適宜施氮肥對根系生長的促進作用有關。施氮并不會迅速促進中間砧蘋果幼樹地上部快速生長,而是通過促進根系Zr的大量合成[19,20,21],對氮素營養的運輸和分配產生加速和定向作用,促使樹體將更多的同化氮分配到根部(表1)用于根系生長[22](圖1),較大根系的快速建立和細根的大量分生,有效提高了氮吸收效率可能是適宜施氮肥N100處理對樹體生長的促進作用較過量施氮肥N200處理更為顯著的最主要原因之一。過量施氮肥N200處理在氮肥施入初期根系15N分配率的顯著升高(表1)對根系生長也產生一定促進作用(圖1),但過量施氮肥條件下土壤中氮素營養超出蘋果幼樹生長需要量,至生長后期大量盈余氮素除以有機態、 生物態氮形式存在以外,還有較高比例以NO3--N形式存在[13],對主根生長仍有顯著促進作用,但高濃度的NO3--N長期持續刺激細胞分裂素含量增加,拮抗生長素,顯著降低根中3-吲哚乙酸(IAA)含量[23,24]和促進乙烯的產生,抑制細根生長[10,11](圖1),雖未對樹體當年生長產生顯著影響,但生長后期較低的細根生物量可能是N200處理翌年春梢旺長期15N再利用率和樹體生物量低于適宜施氮肥N100處理的主要原因之一。不施氮肥N0處理在秋梢旺長期新生器官生長轉為依靠當年吸收氮素營養,土壤中氮素耗竭,誘導根中生長素含量增高,且通過一氧化氮介導的信號途徑刺激根的伸長[9],促使樹體將更多的同化氮分配到根部用于根系生長[22],因此表現為生長后期不施氮肥N0處理逐漸升高的根系15N分配率和根冠比,以及細根的大量分生,但土壤中較低的有效氮濃度仍限制了樹體的快速生長。

本次試驗表明,施氮對樹體生長抑制作用較強的M26中間砧幼樹生長的促進較為緩慢,且低氮和高氮均不利于樹體矮化中間砧蘋果幼樹生長,而傳統肥料一次性大量施用和氮素營養容易淋失、 揮發和反硝化大量損失、 肥效期短的特點容易導致施氮前期形成高氮環境而施氮后期脫肥,已不適應矮砧蘋果對施氮濃度要求較高、 氮濃度過低或過高均不利于其生長的特點,可能也是矮砧蘋果易早衰的主要原因,因此應根據中間砧對樹體生長和氮響應影響的顯著性,改變其栽培過程中的氮肥施用方式。而緩控釋肥料逐漸緩慢釋放養分、 使土壤速效養分的濃度可保持相對穩定,一次施用可滿足植物整個生長期需要和水肥一體化可按照作物生長需求,進行全生育期的水分和養分需求實現水分和養分定量、 定時、 按比例直接提供給作物的優點,可保證蘋果矮化密植集約化栽培中氮肥的充足和均衡供應,對于維持矮化中間砧蘋果健壯樹勢、 提高產量和改善品質,實現豐產、 穩產將具有重要意義。
4 結論
1)不同供氮水平對矮化中間砧蘋果生長及氮素利用率影響顯著,適宜供氮促進矮化中間砧蘋果生長以及氮素利用率提高,這種促進作用是隨著氮肥施入時間的延長,至春稍停長期以后逐漸顯現出來。
2)氮肥對幼樹生長的促進作用與其可有效促進細根的發生有關。
參考文獻:
[1] 陳學森, 韓明玉, 蘇桂林, 等. 當今世界蘋果產業發展趨勢及我國蘋果產業優質高效發展意見[J]. 果樹學報, 2010, 27(4): 598-604.
Chen X S, Han M Y, Su G Letal. Discussion on today’s world apple industry trends and the suggestions on sustainable and efficient development of apple industry in China[J]. J. Fruit Sci., 2010, 27(4): 598-604.
[2] 姜遠茂, 張宏彥, 張福鎖. 北方落葉果樹養分資源綜合管理理論與實踐[M]. 北京: 中國農業大學出版社, 2007.
Jiang Y M, Zhang H Y, Zhang F S. Concepts & practice of integrate nutrient resource management in northern deciduous fruit[M]. Beijing: China Agricultural Press, 2007.
[3] 姜遠茂, 彭福田, 張宏彥. 山東省蘋果園土壤有機質及養分狀況研究[J]. 土壤通報, 2001, 32(4): 167-169.
Jiang Y M, Peng F T, Zhang H Y. Status of organic matter and nutrients in Shandong orchard soils[J]. Chin.J. Soil Sci., 2001, 32(4): 167-169.
[4] 米國華, 陳范駿, 張福鎖. 作物養分高效的生理基礎與遺傳改良[M]. 北京: 中國農業大學出版社, 2012.
Mi G H, Chen F J, Zhang F S. Physiological basis and genetic improvement of nutrient use efficiency in crops[M]. Beijing: China Agricultural Press, 2012.
[5] Liao M T, Palta J A, Fillery I R. Root characteristics of vigorous wheat improve early nitrogen uptake[J]. Australian J. Agric. Res., 2006, 57: 1097-1107.
[6] 宇萬太, 周 樺, 徐永剛, 等. 追施氮肥當年與翌年對桉樹生長及各部位氮貯量的影響[J]. 生態學雜志, 2010, 29(9): 1703-1708.
Yu W T, Zhou H, Xu Y Getal. Eucalypt growth and nitrogen storage in the current year and next year of nitrogen top dressing[J]. Chin. J. Ecol., 2010, 29(9): 1703- 1708.
[7] 韓振海, 曾 驤, 王福鈞. 晚秋葉施尿素和生長調節劑對富士蘋果幼樹貯藏氮素的影響[J]. 園藝學報, 1992, 19(1): 15-21.
Han Z H, Zheng X, Wang F J. Effects of autumn foliar application of 15N-urea and several pgr on N storage and reuse in apple[J]. Acta Hort. Sin., 1992, 19(1): 15-21.
[8] 羅雪華, 鄒碧霞, 吳菊群, 等. 氮水平和形態配比對巴西橡膠樹花藥苗生長及氮代謝、 光合作用的影響[J]. 植物營養與肥料學報, 2011, 17(3): 693-701.
Luo X H, Zou B X, Wu J Qetal. Effects of different nitrogen levels and NH4+/NO3-ratios on the growth, nitrogen metabolism and photosynthesis of anther-derived somatic seedlings ofHeveabrasiliensis[J]. Plant Nutr. Fert. Sci., 2011, 17(3): 693-701.
[9] Zhao D Y, Tian Q Y, Li L H, Zhang W H. Nitric oxide is involved in nitrate-induced inhibition of root elongation in Zea mays L[J]. Ann Bot, 2007(100): 497-503.
[10] Tian Q Y, Chen F J, Liu J Xetal. Inhibition of maize root growth by high nitrate supply is correlated with reduced IAA levels in roots[J]. J. Plant Physiol, 2008 (165): 942-951.
[11] Tian Q Y, Sun P, Zhang W H. Ethylene is involved in nitrate-dependent root growth and branching in Arabidopsis thaliana[J]. New Phytol, 2009 (184): 918-931.
[12] 趙 林, 姜遠茂, 彭福田, 等. 嘎拉蘋果對春施15N-尿素的吸收、 利用與分配特性[J]. 植物營養與肥料學報, 2009, 15(6): 1439- 1443.
Zhao L, Jiang Y M, Peng F Tetal. Characteristics of absorption, utilization and distribution of spring soil15N-urea application for Gala/Malushupehensis[J]. Plant Nutr. Fert. Sci., 2009, 15 (6): 1439-1443.
[13] 巨曉棠, 潘家榮, 劉學軍, 等. 北京郊區冬小麥/夏玉米輪作體系中氮肥去向研究[J].植物營養與肥料學報, 2003, 9(3): 264-270.
Ju X T, Pan J R, Liu X Jetal. Study on the fate of nitrogen fertilizer in winter wheat/summer maize rotation system in Beijing suburban[J]. Plant Nutr. Fert. Sci., 2003, 9(3): 264- 270.
[14] 鐘茜, 巨曉棠, 張福鎖. 華北平原冬小麥/夏玉米輪作體系對氮素環境承受力分析[J]. 植物營養與肥料學報, 2006, 12(3): 285-293.
Zhong Q, Ju X T, Zhang F S. Analysis of environmental endurance of winter wheat/summer maize rotation system to nitrogen in North China Plain[J]. Plant Nutr. Fert. Sci., 2006, 12 (3): 285-293.
[15] 劉新宇, 巨曉棠, 張麗娟, 等. 不同施氮水平對冬小麥季化肥氮去向及土壤氮素平衡的影響[J]. 植物營養與肥料學報, 2010, 16(2): 296-303.
Liu X Y, Ju X T, Zhang L Jetal. Effects of different N rates on fate of N fertilizer and balance of soil N of winter wheat[J]. Plant Nutr. Fert. Sci., 2010, 16 (2): 296-303.
[16] 顧曼如, 束懷瑞, 周宏偉. 蘋果氮素營養研究Ⅳ貯藏15N的運轉、 分配特性[J].園藝學報, 1986, 13 (1): 25-30.
Gu M R, Shu H R, Zhou H W. A study on the nitrogen nutrition of apple trees IV. The characters of translocation and distribution of the reserved15N[J]. Acta Hort. Sin., 1986, 13 (1): 25-30.
[17] 趙營, 同延安, 趙護兵.不同供氮水平對夏玉米養分積累、 轉運及產量的影響[J]. 植物營養與肥料學報, 2006, 12(5): 622-627.
Zhao Y, Tong Y A, Zhao H B. Effects of different N rates on nutrients accumulation, transformation and yield of summer maize[J]. Plant Nutr. Fert. Sci., 2006, 12(5): 622-627.
[18] 曾驤. 棗樹葉片內氮素貯藏和循環利用的研究[J]. 核農學報, 1991, 5(1): 37-43.
Zeng X. Storage and recycling utilization of leaf-nitrogen of jujube tree[J]. J. Nucl. Agric. Sci., 1991, 5(1): 37-43.
[19] Beck E H. Regulation of shoot/root ratio by cytokinins from roots in Urtica dioica: opinion[J]. Plant Soil, 1996(185): 3-12.
[20] Takei K, Sakakibara H, Sugiyama T. Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana[J]. J. Biol. Chem., 2001, 276(28): 26405-26410.
[21] 周鵬, 彭福田, 魏紹沖, 等. 氮素形態對平邑甜茶細胞分裂素水平和葉片生長的影響[J]. 園藝學報, 2007, 34(2): 269-274.
Zhou P, Peng F T, Wei S Cetal. Effects of rhizosphere nitrate and ammonium on the level of cytokinins and leaf growth ofMalushupenensisRehd[J]. Acta Hort. Sin., 2007, 34(2): 269-274.
[22] 李燕婷, 米國華, 陳范駿, 等. 玉米幼苗地上部/根間氮的循環及其基因差異[J].植物生理學報, 2001, 27(3): 226-230.
Li Y T, Mi G H, Chen F Jetal. Genotypic difference of nitrogen recycling between root and shoot of maize seedlings[J]. Acta Phytoph. Sin., 2001, 27(3): 226-230.
[23] Walch L P, Ivanov, I I, Filleur Setal. Nitrogen regulation of root branching[J]. Ann. Bot., 2006, 97: 875-881.
[24] Caba J M, Centeno M L, Fernandez Betal.Inoculation and nitrate alter phytohormone levels in soybean roots: differences between a supernodulating mutant and the wild type[J]. Planta, 2000, 211: 98-104.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
