骨唇黃河魚耳石早期形態發育和輪紋特征研究
2014-03-29 01:45:23嚴太明胡佳祥趙柳蘭
水生生物學報 2014年4期
關鍵詞:生長
嚴太明 胡佳祥 楊 婷 趙柳蘭 何 智
(四川農業大學動物科技學院, 雅安 625014)
YAN Tai-Ming, HU Jia-Xiang, YANG Ting, ZHAO Liu-Lan and HE Zhi
(College of Animal Science and Technology, Sichuan Agricultural University, Ya’an 625014, China)
骨唇黃河魚耳石早期形態發育和輪紋特征研究
嚴太明 胡佳祥 楊 婷 趙柳蘭 何 智
(四川農業大學動物科技學院, 雅安 625014)
研究了骨唇黃河魚仔稚魚耳石在實驗室養殖條件下的發育過程和生長特點, 確證了輪紋沉積規律。結果表明, 在14.0—17.8℃孵化條件下, 微耳石和矢耳石在受精后96h 30min出現, 星耳石在出膜后第16天出現。仔稚魚生長過程中矢耳石形狀變化較大, 由出膜時的圓形發育到穩定時的箭矢狀。微耳石由近圓形發育成貽貝形, 其中心核位置隨發育明顯偏移。星耳石形狀不規則, 從出現時的心形發育成為星芒狀。微耳石和矢耳石在前后軸方向上后區的生長快于前區(P<0.05); 在背腹軸方向上, 微耳石腹區的生長快于背區(P<0.05),矢耳石背區的生長快于腹區(P<0.05), 兩對耳石的前后區半徑之和與全長均呈線性相關。微耳石和矢耳石的第1個輪紋均在出膜后第2天形成, 新增的輪紋數(微耳石IL, 矢耳石IS)與出膜后的天數(D)表現出顯著的線性相關, 方程分別為: IL=0.9911D-1.0008(R2=0.9971, n=220, P<0.001)和IS=0.9925D-0.10873(R2=0.9919, n=161, P<0.001), 方程的斜率與1均無顯著差異(P>0.05), 表明兩對耳石輪紋沉積均呈日周期性, 生長輪為日輪。研究結果豐富了骨唇黃河魚的發育生物學資料, 可為研究其自然種群早期生活史提供參考。
耳石; 形態發育; 日輪; 生長; 骨唇黃河魚
魚類耳石是頭部半規管中的微小鈣化組織,有探測聲音和感知平衡的功能[1,2]。在早期發育階段, 耳石的生長與發育除受環境因子的影響之外[3],更主要的是由遺傳因素決定, 從而表現出高度的物種特異性[4,5]。因此, 可以利用耳石形態差異來進行魚類分類學研究[6]。然而, 耳石早期形態變化過程的研究還相對較少且詳略不一。
多數硬骨魚類微耳石和矢耳石的輪紋具有日周期性[7—9], 且能記錄早期發育階段所經歷的孵化、營養轉換、卵胎生魚類產出、變態、定居以及環境因子變化等事件[10,11]。近年來, 耳石微結構分析已成功應用于魚類早期生活史研究以及產卵場位置和產卵期的推算[12—14]。但是, 在應用之前均應通過養殖試驗對耳石日輪沉積規律和輪紋特征進行評估和驗證。
骨唇黃河魚(Chuanchia labiosa Herzenstein)是鯉形目(Cypriniformes)裂腹魚亞科(Schizothoracinae)中的單屬單種, 僅分布在我國四川、甘肅和青海省境內黃河上游干流及其支流海拔3000—4300 m的高原寬谷河段和湖泊中, 野生資源已極度匱乏, 被列為易危物種[15], 對它的研究僅見于分類學方面的報道[15,16]。本實驗觀察了骨唇黃河魚耳石早期形態發育過程與生長特點, 并確證了微耳石和矢耳石輪紋形成的規律, 以期為研究其在野外狀況下的早期生活史特點和資源變動規律提供參考。
1 材料與方法
1.1 實驗魚來源與管理
2011年4月5日, 在雅安市蘆山縣寶劍漁業有限公司養殖場通過人工授精方法獲得骨唇黃河魚受精卵, 帶回四川農業大學實驗室進行常溫(14.0—17.8 )℃靜水孵化, 用充氧泵充氧。選擇在1h內先后出膜的仔魚放于白塑料盆(36 cm×28 cm× 12 cm)中作為本實驗材料。每天9:00和21:00將魚分別置于20℃和12℃的生化培養箱培育。以鹵蟲無節幼體作為開口餌料, 隨后投喂足量剁碎并消毒的水蚯蚓, 每日投喂兩次。
1.2 耳石摘取與處理
從出膜至第 26天每天取仔魚 8—10尾, 第 27至第62天每隔3天取樣一次, 每次取樣5尾。實驗魚用鹽酸利多卡因(濃度為 30 mL/L)麻醉后放置在Leica DC500照相系統下拍照, 并用系統自帶軟件測量魚體全長(精確度0.1 mm)。照相后的仔稚魚在顯微鏡下挑出 3對耳石, 無水乙醇清洗后用中性樹膠固定于載玻片上備用。耳石打磨參照Song, et al.[8]的方法進行。因星耳石的輪紋不清晰和無規律性,本實驗未作輪紋數分析。
1.3 輪紋計數和照相
耳石用二甲苯透明后置于光學顯微鏡下計數輪紋, 方法參照 Song, et al.[8]的報道進行。用朱旗等[17]設計的計算機圖像分析系統測量微耳石和矢耳石半徑以及輪紋間距。選取典型樣本在Nikon顯微成像系統下照相。
1.4 數據及圖片處理
用Excel 2007擬合出全長、耳石半徑、耳石輪紋數與日齡以及耳石半徑與體長的關系, 用SPSS Statistics 17.0對上述關系式進行差異顯著性檢驗和方差分析。用Adobe photoshop CS2處理耳石圖片。
2 結果

圖1 骨唇黃河魚耳石形態發育Fig. 1 Otoliths development in C. Labiosa
2.1 耳石形態發育
骨唇黃河魚受精卵淡黃色, 弱黏性,吸水后卵徑平均為(3.30±0.12) mm(n=15)。受精后72h聽囊出現, 呈橢圓形。至96h 30min時微耳石和矢耳石出現。孵化歷時 139h開始出膜, 初孵仔魚全長8.5—9.0 mm [平均(8.81±0.24) mm, n=10], 眼灰黑色,鰓弓 3對, 身體透明無色素, 胸鰭原基扇形, 肌節54—56對。星耳石在出膜后第16天出現, 3對耳石形態隨發育逐漸變化直至穩定(圖1)。
微耳石 微耳石由近圓形發育成貽貝形, 整體形態變化不大, 但中心核相對位置隨發育偏移明顯(圖 1)。微耳石在出膜 7d前近似圓形, 中心核居中。第13天時后端基葉生長較后端快, 輪間距較后端寬, 向腹部方向生長快于向背部, 中心核相對位置開始向前端背側方向移動。第 16天耳石橢圓形,背側平緩, 中心核相對位置向前端背側移動明顯,位于前端和背部方向約1/3處。第29天耳石整體輪廓呈扇形, 后端基葉進一步生長, 腹側突起尖圓弧狀, 背側微內凹。第42天時耳石基葉凸起, 腹部外緣尖圓, 背側中部明顯內凹。第52天后外形呈貽貝形, 后端基葉末端略尖, 鄰近基葉的背部微內凹,邊緣光滑, 中心核相對位置向前移至近前端約 3/10處和背側約1/4處。
矢耳石 矢耳石由圓形發育成箭矢狀, 形態變化顯著(圖 1)。出膜后前 13天耳石近圓形, 中心核居中。第16天呈橢圓形, 后端基葉快速生長使中心核相對位置向前側微偏移。第22天外部形態明顯變化, 后端基葉突出成鍥形狀晶狀突, 中心核相對位置向前端偏移, 仍處于背腹中部。第29天呈梭形,背腹部基本對稱, 外緣略圓凸, 后端基葉變薄成略尖的晶狀突, 前端呈薄的橈片狀。第42天耳石明顯向背側方向彎曲, 后端基葉向后快速延伸變得細長且背側平直, 腹側翼葉變寬并出現隆起的嵴。第 52天耳石整體較之前平直, 后端翼葉生長使基葉變寬,末端尖銳, 前端腹側翼葉開始出現缺刻。第 62天及以后, 耳石呈箭矢狀, 前后端腹側翼葉缺刻加深, 后端基葉背側平直, 與小凸起的翼葉之間形成內凹。
星耳石 星耳石形狀總體不規則(圖1)。出膜后第16天星耳石開始出現, 心形, 中心核有明顯的小黑色素團。至第29天耳石變成不規則四邊形, 中心核相對位置明顯偏移。至42d時外形近似半圓形,外緣開始出現微凸的嵴。52d以后近似扇形, 外緣有凸起的嵴3—4個, 隨發育缺刻凹入更深成星芒狀。
2.2 耳石微結構
微耳石和矢耳石的中心核及原基 統計表明,每對耳石左側和右側的中心核和原基數無顯著差異(P>0.05), 左右耳石混合后各組合類型所占比例見表 1。多數耳石上具有一個近圓形的生長中心即中心核, 其內深黑色的圓點為耳石原基(圖2)。在微耳石、矢耳石和星耳石上具有1個中心核和1個原基的比例分別為71.07%、67.18%和74.19%, 具1個中心核和 2個原基的比例分別為 18.95%、15.64%和18.28%, 在微耳石和矢耳石中有1個中心核和3個原基的分別占 6.48%和 11.03%, 星耳石中則僅有1.08%, 其他類型出現幾率則較低。

表1 具有不同中心核和原基數的耳石在3種耳石中的比例Tab.1 The proportions of otoliths with different numbers of nucleus and primordium among sagitta, lapillus and asteriscus

圖2 中心核或原基數目不同的骨唇黃河魚耳石Fig. 2 Otoliths with different numbers of nuclei or primordia in C. labiosa

圖3 骨唇黃河魚仔稚魚耳石輪紋Fig. 3 Increments of otolith of larvae and juvenile in C. labiosa
微耳石中心核面積(162.8±36.51) μm2, 長直徑變幅為11.43—16.65 μm, 平均(13.89±1.46) μm (n=32),短直徑變幅為7.01—13.89 μm, 平均(10.93±1.83) μm (n=32)。矢耳石中心核面積(230.17±40.16) μm2, 長直徑12.38—20.54 μm, 平均(17.50±2.25) μm(n=33),短直徑10.25—17.33 μm, 平均(13.83±1.88) μm(n=33)。
耳石輪紋特征 初孵仔魚微耳石和矢耳石上均未觀察到輪紋, 岀膜后第 2天出現第一個輪紋。29日齡仔魚微耳石出現28輪紋(圖3a), 21日齡仔魚矢耳石出現20輪紋(圖3b)。統計結果發現, 微耳石(Increment number of lapillus, IL)和矢耳石(Increment number of sagitta, IS)的輪紋數與出膜后的天數(Days post hatching, D)呈線性相關(圖4), 回歸方程分別為: IL=0.9911D-1.0008(R2=0.9971, n=220, P<0.001)(圖4a)和IS=0.9925D-0.10873(R2=0.9919, n=161, P<0.001) (圖 4b)。經檢驗, 關系式中的斜率與 1均無顯著差異(P>0.05), 表明兩對耳石的輪紋沉積呈日周期性,生長輪為日輪。
輪間距 對30日齡仔魚微耳石(n=22)和矢耳石(n=20)的輪紋間距進行了測量, 確證了輪紋間距與輪紋增量間(距耳石生長中心的輪紋數)的關系。結果表明, 隨著輪紋沉積, 輪紋間距有增大的趨勢,但在第 6輪附近輪紋間距相對較寬, 其中微耳石最大平均寬度為(2.25±0.11) μm(圖 5a), 矢耳石為(3.12±0.25) μm(圖5b)。
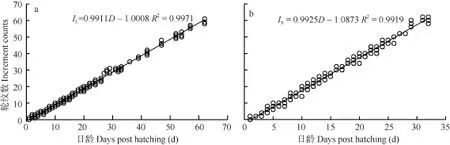
圖4 骨唇黃河魚微耳石(a)和矢耳石(b)輪紋數與日齡的關系Fig. 4 Relationship between increment counts and days post hatching of lapillus (a) or sagitta (b) in C. labiosa
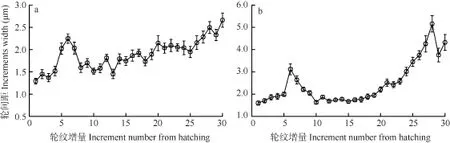
圖5 骨唇黃河魚微耳石(a)和矢耳石(b)輪紋間距與輪紋增量間的關系Fig. 5 Relationship between increment width and increment number of lapillus (a) and sagittal (b) in C. labiosa. Bars indicate standard deviation
2.3 耳石的生長特征
耳石的生長 以耳石核為中心分別測量了微耳石和矢耳石前區(Otolith radius of anterior in lapillus, OAL, Otolith radius of anterior in sagitta, OAS)、后區(OPL, OPS)、背區(ODL, ODS)和腹區(OVL, OVS)的半徑。結果表明, 微耳石和矢耳石前區半徑(OAL, OAS)和后區半徑(OPL, OPS)與出膜時間(D)呈指數相關(圖6a、c), 而背區半徑(ODL, ODS)和腹區半徑(OVL, OVS)與出膜時間(D)呈冪函數相關(圖6b、d)。研究也發現, 兩對耳石前后軸方向后區的生長快于前區(P<0.05); 在背腹軸方向上微耳石腹區的生長快于背區(P<0.05), 矢耳石背區的生長快于腹區(P<0.05)。
耳石長度與全長的關系 微耳石(Otolith length of lapillus, OLL)和矢耳石(Otolith length of sagitta, OLS)長度(前區和后區半徑之和)與全長(Total length, TL)均呈線性相關(如圖 7), 回歸方程分別為: OLL=118.29TL+6.9919(R2=0.9683, n=212, P<0.001) (圖 7a)和 OLS=71.656TL+8.3292(R2=0.9654, n=212, P<0.001)(圖7b)。
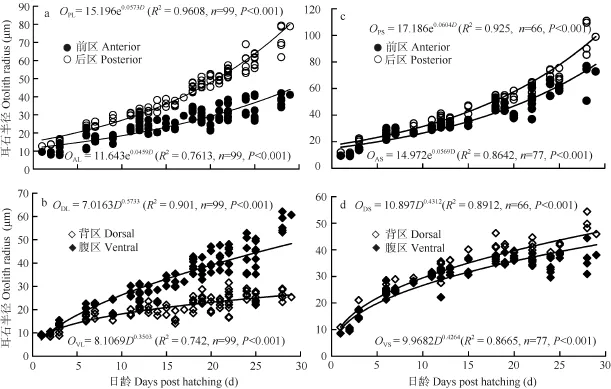
圖6 骨唇黃河魚耳石微耳石(a, b)和矢耳石(c, d)半徑與出膜時間的關系Fig. 6 Relationship between otolith radius of different area in lapillus (a, b) or sagitta (c, d) and days post hatching in C. labiosas
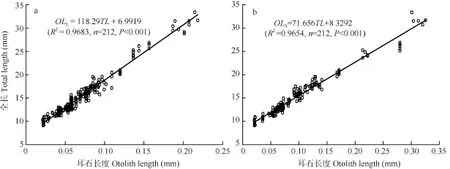
圖7 骨唇黃河魚微耳石(a)和矢耳石(b)長度(前區和后區半徑之和)與全長的關系Fig. 7 Relationship between length (the sum length of radius anterior and posterior in otolith ) of lapillus (a) or sagitta (b) and total length in C. labiosa
3 討論
3.1 耳石的形態發育
本研究中骨唇黃河魚的微耳石由近圓形發育成貽貝形, 整體形態變化不大。而叉尾斗魚(Macropodus opercularis)的微耳石剛出現時為不規則的小型結晶體, 然后逐漸沉積為圓盤狀, 孵出后7d轉變為近橢圓形, 孵出后19d轉變為中部圓凸兩端較尖的菱形[18]。出膜前唐魚(Tanichthys albonubes)的微耳石形狀不規則, 含有一些結晶狀物質, 孵出時仔魚的微耳石通常為圓盤狀, 第 16天后轉變為近似橢圓形,30d后轉變為梨形[19]。鰱(Hypophthalmichthys molitrix)、草魚(Ctenopharynodon idellus)和鳙(Aristichthys nobilis)的微耳石形態相對穩定, 經歷圓形、豆形、長豆形到腰部形成一縊部的鍥形[20]。這些結果表明,魚類微耳石早期形態相對穩定, 其發育受生理活動和環境變化等的影響不大。
研究也表明, 骨唇黃河魚的矢耳石由初孵仔魚的圓形或近圓形, 經過橢圓形、菱形, 最后漸變為前端長尖后端圓鈍的長箭矢形, 形態變化明顯。唐魚仔魚矢耳石發育經歷了前端較尖、后端圓鈍、中間呈卡腰狀的發育階段[19], 鴨綠江香魚(Flecoglossus altivelis)矢耳石則經歷了圓錐形的發育過程[21]。鳙的矢耳石由前端伸長變尖, 后端圓鈍的菱形, 最后形成中部兩側有薄片狀突出的箭矢形; 鰱經歷了一端長尖, 另一端長圓, 中部兩側有薄片狀突出的長菱形狀的發育過程[22]。胭脂魚(Myxocyprinus asiaticus)矢耳石的中間發育過程包括紡錘形, 后端梨形, 前端鈍圓等幾個階段[8]。這些研究結果表明, 淡水鯉形目魚類的矢耳石形態發育過程大致相似, 但也表現出一定的種類特異性。
3.2 耳石輪紋出現及日增量
魚類耳石上第一日輪的形成時間存在種間差異,與環境條件有一定的關系。如藍羅非魚(Oreochromis aureus)[23]和白鮭(Coregonidae coregonus)[24]矢耳石第一日輪在孵化前形成, 但大多數魚類在孵化后出現。黑鯛(Acanthopagrus schlegeli)[25]、狹鱈(Theragra chalcogramma)[26]以及黑鱸(Micropterus salmoides)[27]孵化時形成, 南方鲇(Silurus meridionalis)[28]和白仔鰻(Temmincket Schlegel)[29]于孵化后24h內出現, 尼羅羅非魚(Oreochromis niloticus)[30]、草魚和大銀魚(Protosalanx hyalocranius)[31]在出膜后第2天可見第一日輪, 梭魚(Chelon haematocheilus)卻在第3天出現[32]。此外, 耳石第一條日輪的形成受攝食等因素的影響[33,34], 如遮目魚(Chanos chanos)[35]第一日輪形成與內源性營養轉換有關, 竹莢魚(Trachurus japonicus)則與開口攝食有關[36]。由此可見, 耳石第一日輪出現與魚類早期發育階段的一些重要事件相關聯。本實驗中骨唇黃河魚仔魚微耳石和矢耳石在剛孵化出膜時均未出現輪紋, 第 2天才出現第一個輪紋。通過對耳石輪紋數的統計發現, 微耳石和矢耳石的輪紋數與日齡呈線性相關, 其斜率與 1均無顯著差異(P>0.05), 表明其耳石輪紋沉積呈日周期性, 生長輪為日輪。這與胭脂魚[8]、唐魚[19]和南方鲇[28]等魚類的觀察結果一致。
3.3 耳石生長特征
耳石的形態發育和輪紋特征可以和個體發育過程以及棲息環境變化等事件相對應[14]。因此, 在建立魚類早期生長發育過程與耳石形態和微結構關系基礎上, 可以通過耳石形態結構分析研究個體的早期生長發育特點、環境狀況以及種群補充等關鍵問題[36,37]。本實驗表明, 30日齡前骨唇黃河魚微耳石和矢耳石在前后軸和背腹軸方向的生長速度存在明顯差異, 其中前后區呈指數增長, 背腹區為冪函數生長。總體來看, 耳石前后區半徑之和與全長均呈顯著的線性相關, 說明微耳石和矢耳石形態變化與仔稚魚的生長發育狀況關系密切。
本研究發現, 與安邦雀鯛(Pomacentrus amboinensis)[38]和綠刺尾鯛(Acanthurus triostegus)[39]等輪紋寬度呈現了由窄到寬的規律不同, 骨唇黃河魚微耳石和矢耳石的輪紋寬度在出膜后 6d左右突然增大, 這剛好與仔魚營養轉換的時間點相吻合, 由此推斷二者之間可能存在一定的關聯[19,40]。這表明耳石的輪紋間距等微結構特點能反映魚類早期生活史中的一些發育事件。
[1] Degens E T, Deuser W G, Haedrich R L. Molecular structure and composition of fish otoliths [J]. Marine Biology, 1969, 2(2): 105—113
[2] Popper A N, Fay R R, Platt C. Sound detection mechanisms and capabilities of teleost fishes [A]. In: Collin S P, Marshall N J (Eds.), Sensory Processing in Aquatic Environments [C]. New York. Springer-Verlag. 2003, 3—38
[3] Capoccioni F, Costa C, Aguzzi J, et al. Ontogenetic and environmental effects on otolith shape variability in three Mediterranean European eel (Anguilla anguilla, L.) local stocks [J]. Journal of Experimental Marine Biology and Ecology, 2010, 397(1): 1—7
[4] Vignon M, Morat F. Environmental and genetic determinant of otolith shape revealed by a non-indigenous tropical fish [J]. Marine Ecology Progress Series, 2010, 411: 231—241
[5] Parmentier E, Vandewalle P, Lagardère F. Morpho-anatomy of the otic region in carapid fishes: eco-morphological study of their otoliths [J]. Journal of Fish Biology, 2001, 58(4): 1046—1061
[6] Lord C, Morat F, Lecomte-Finiger R, et al. Otolith shape analysis for three Sicyopterus (Teleostei: Gobioidei: Sicydiinae)species from New Caledonia and Vanuatu [J]. Environmental Biology of Fishes, 2012, 93(2): 209—222
[7] Pannella G. Fish otoliths: daily growth layers and periodical patterns [J]. Science, 1971, 173(4002): 1124—1127
[8] Song Z B, Fu Z D, Li J, et al. Validation of daily otolith increments in larval and juvenile Chinese sucker, Myxocyprinus asiaticus [J]. Environmental Biology of Fishes, 2008, 82(2): 165—171
[9] Yamada H, Chimura M, Asami K, et al. Otolith development and daily increment formation in laboratory-reared larval and juvenile black-spot tuskfish Choerodon schoenleinii [J]. Fisheries Science, 2009, 75(5): 1141—1146
[10] Joh M, Matsuda T, Satoh N, et al. Otolith microstructure of brown sole Pseudopleuronectes herzensteini: validation of daily ring formation and the occurrence of microstructure denoting metamorphosis [J]. Fisheries Science, 2011, 77(5): 773—783
[11] Vignon M. Ontogenetic trajectories of otolith shape during shift in habitat use: Interaction between otolith growth and environment [J]. Journal of Experimental Marine Biology and Ecology, 2012, 420—421: 26—32
[12] Correia A T, Able K, W Antunes C, et al. Early life history of the American conger eel (Conger oceanicus) as revealed by otolith microstructure and microchemistry of metamorphosing leptocephali [J]. Marine Biology, 2004, 145(3): 477—488
[13] La Mesa M. The utility of otolith microstructure in determining the timing and position of the first annulus in juvenile Antarctic toothfish (Dissostichus mawsoni) from the South Shetland Islands [J]. Polar Biology, 2007, 30(10): 1219—1226
[14] Campana S E. Otolith science entering the 21st century [J]. Marine and Freshwater Research, 2005, 56(5): 485—495
[15] Yue P Q, Shan X H, Lin R D, et al. Fauna Sinica: Osteichthyes Cypriniformes Ш [M]. Beijing: Science Press. 2000, 385—386 [樂佩琪, 單鄉紅, 林人端, 等. 中國動物志硬骨魚綱鯉形目(下卷). 北京: 科學出版社. 2000, 385—386]
[16] Ding R H. The Fishes of Sichuan, China [M]. Chengdu: Sichuan Publishing House of Science & Technology. 1994, 370—371 [丁瑞華. 四川魚類志. 成都: 四川科學技術出版社. 1994, 370—371]
[17] Zhu Q, Xia L Q, Chang J B. Computer identification on otolith microstructure of fish [J]. Acta Hydrobiologica Sinica, 2002, 26(6): 600—604 [朱旗, 夏立啟, 常劍波. 魚類耳石微結構的計算機自動識別. 水生生物學報, 2002, 26(6): 600—604]
[18] Zhao T, Chen G Z, Lin X T. Otolith ontogeny and increment formation of larval Macropodus opercularis [J]. Journal of Fishery Sciences of China, 2010. 17(6): 1364—1370 [趙天,陳國柱, 林小濤. 叉尾斗魚仔魚耳石形態發育與日輪形成特征. 中國水產科學, 2010, 17(6): 1364—1370]
[19] Shi F, Sun J, Lin X T, et al. Otolith ontogeny and increment formation in larval Tanichthys albonubes [J]. Chinese Journal of Zoology, 2006, 41(4): 10—16 [史方, 孫軍, 林小濤, 等. 唐魚仔魚耳石的形態發育及日輪. 動物學雜志, 2006, 41(4): 10—16]
[20] Zeng X B. Studies on otolith mophology in larval-young silver carp, big head, and grass carp with the application in species discrimination [D]. Thesis for Master of Science, Huazhong Agricultural University, Wuhan. 2001 [曾祥波.鰱、鳙和草魚仔幼魚耳石形態及種類鑒別中的應用. 碩士學位論文, 華中農業大學, 武漢. 2001]
[21] Xie Y H, Li B, Fu L J, et al. On daily-growth increment of otolith and growth of ayu fish from Yalüjiang River [J]. Acta Zoologica Sinica, 1995, 41(2): 125—133 [解玉浩, 李勃, 富麗靜, 等. 鴨綠江香魚耳石日輪與生長的研究. 動物學報, 1995, 41(2): 125—133]
[22] Xie Y H, Li B, Fu L J, et al. Daily growth increment in otolith and growth for fry-young fish of bighead (Aristichthys nobilis) [J]. Journal of Fishery Sciences of China, 1995, 2(2): 34—42 [解玉浩, 李勃, 富麗靜, 等. 鳙仔-幼魚耳石日輪與生長的研究. 中國水產科學, 1995, 2(2): 34—42]
[23] Karakiri M, Hammer C. Preliminary notes on the formation of daily increments in otoliths of Oreochromis aureus [J]. Journal of Applied Ichthyology, 1989, 5(2): 53—60
[24] Eckmann R, Rey P. Daily increments on the otoliths of larval and juvenile Coregonus spp., and their modification by environmental factors [J]. Hydrobiology, 1987, 148(2): 137—143
[25] Huang W B, Chiu T S. Daily increments in otoliths and growth equation of black porgy, Acanthopagrus schlegeli, larvae [J]. Acta Zoologica Taiwanica, 1997, 8(2): 121—131
[26] Nishimura A, Yamada J. Age and growth of larval and juvenile walleye pollock, Theragra chalcogramma (Pallas), as determined by otolith daily growth increments [J]. Journal of Experimental Marine Biology and Ecology, 1984, 82(2—3): 191—205
[27] Miller S J, Storck T. Daily growth rings in otoliths of young-of-the-year largemouth bass [J]. Transactions of the American Fisheries Society, 1982, 111(4): 527—530
[28] Yang F, Peng W H. A study of the daily growth increment of otoliths in the larvae and juvenile of Silurus slidatovi meridionalls Chen [J]. Journal of southwest Agricultural University, 2001, 23(4): 340—342 [楊帆, 彭文輝. 南方鲇仔幼魚耳石日輪的研究. 西南農業大學學報, 2001, 23(4): 340—342]
[29] Li C H, Sha X S. Periodicity of increment formation in the otoliths of laboratory-reared Anguilla Japonica larvae [J]. Oceanologia et Limnologia Sinica, 1995, 26(4): 408—413 [李城華, 沙學紳. 日本鰻鱺早期階段耳石日生長輪形成的周期. 海洋與湖沼, 1995, 26(4): 408—413]
[30] Zhang Z, Runham N W. Otolith microstructure pattern in Oreochromis niloticus (L.) [J]. Journal of Fish Biology, 1992, 40(3): 325—332
[31] Fu L J, Xie Y H, Li B, et al. Studies on daily-growth increment of otolith and growth of Lare Icefish [J]. Journalof Fishery Sciences of China, 1997, 4(2): 21—27 [富麗靜,解玉浩, 李勃, 等. 大銀魚耳石日輪與生長的研究. 中國水產科學, 1997, 4(2): 21—27]
[32] Li C H, Sha X H, You F, et al. Daily growth increments in otoliths of Mullet larvae Mugil so-iuy basilewsky and determination from field-collected ones [J]. Oceanologia et Limnologia Sinica, 1993, 24(4):345—349 [李城華, 沙學紳,尤鋒, 等. 梭魚仔耳石日輪形成及自然種群日齡的鑒定.海洋與湖沼, 1993, 24(4): 345—349]
[33] Morley S A, Belchier M, Dickson J, et al. Daily otolith increment validation in larval mackerel icefish, Champsocephalus gunnari [J]. Fisheries Research, 2005, 75(1—3): 200—203
[34] H?ie H, Folkvord A, Johannessen A. Maternal, paternal and temperature effects on otolith size of young herring (Clupea harengus L.) larvae [J]. Journal of Experimental Marine Biology and Ecology, 1999, 234(2): 167—184
[35] Tzeng W N, Yu S Y. Daily growth increments in otoliths of milkfish, Chanos chanos (Forssk?l), larvae [J]. Journal of Fish Biology, 1988, 32(4): 495—504
[36] Xie S G, Watanabe Y. Transport-determined early growth and development of jack mackerel Trachurus japonicus juveniles immigrating into Sagami Bay, Japan [J]. Marine & Freshwater Research, 2007, 58(11): 1048—1055
[37] Ma Q Y, Xue Y, Xu B D, et al. Relationships between otolith size and fish size for twelve prey fish species from Jiaozhou Bay [J]. Acta Hydrobiologica Sinica, 2013, 37(3): 481—487 [麻秋云, 薛瑩, 徐賓鐸, 等. 膠州灣12種餌料魚類耳石大小與體長的關系. 水生生物學報, 2013, 37(3): 481—487]
[38] Wilson D T, McCormick M I. Microstructure of settlementmarks in the otoliths of tropical reef fishes [J]. Marine Biology, 1999, 134(1): 29—41
[39] McCormick M I. Delayed metamorphosis of a tropical reef fish (Acanthurus triostegus): a field experiment [J]. Marine Ecology Progress Series, 1999, 176: 25—38
[40] Song Z B, Chang J B, Cao W X. Natural checks and clarity of increments in larval grass carp (Ctenop haryngodon idellus) otoliths [J]. Acta Zoologica Sinica, 2003, 49(4): 508—513 [宋昭彬, 常劍波, 曹文宣. 草魚仔魚耳石的自然標記和生長輪的清晰度. 動物學報, 2003, 49(4): 508—513]
STUDY ON THE OTOLITH DEVELOPMENT AND THE FORMATION OF INCREMENTS IN LARVAE AND JUVENILE OF CHUANCHIA LABIOSA
Daily observations were made on the morphological development and the growth in increments of the otolith in larvae and juvenile of Chuanchia labiosa in rearing conditions. The sagitta and the lapillus appeared at 96h and 30min after the fertilization, and the asteriscus first appeared at 16 days post hatching. The shape of the sagitta was rounded at hatching and gradually changed into long elliptical with a long pointy posterior in the end. The lapillus was also rounded at hatching, and changed into a mussel shape at the end of the experiment. Compared to the sagitta and the lapillus, the shape of the asteriscus remained mussel-like and was relatively stable during the development. We compared the data of the radius of the dorsal, ventral, anterior and posterior areas in the sagitta and the lapillus. It showed that in the lapillus the growth of the posterior was faster than that of the anterior (P<0.05), and the ventral grew faster than the dorsal (P<0.05); in the contrast the dorsal grew faster than the ventral in the sagitta (P<0.05). The combined length of the radius of the anterior and the posterior of the sagitta was linearly related to the total length of the larvae and the juvenile, and the same was observed in the lapillus. The first increment on the sagitta and the lapillus appeared on the second day post hatching and from then on there was one new increment formed every day. The number of increments on the lapillus (IL) and the sagitta (IS) showed a linear relationship with the number of days post hatching (D), with the equations of IL=0.9911D–1.0008 (R2=0.9971, n=220, P<0.001) and IS=0.9925D–0.10873 (R2=0.9919, n=161, P<0.001), respectively. The slopes of the equations were statistically close to 1.0 (P>0.05). These results suggested that the number of the otolith increments increased by 1 on a daily basis. Our study contributed to the understanding in the development of C. labiosa, as well as in its early life history in the natural environment.
Otoliths; Morphology development; Daily increments; Growth; Chuanchia labiosa
YAN Tai-Ming, HU Jia-Xiang, YANG Ting, ZHAO Liu-Lan and HE Zhi
(College of Animal Science and Technology, Sichuan Agricultural University, Ya’an 625014, China)
Q174
A
1000-3207(2014)04-0764-08
10.7541/2014.107
2013-12-18;
2014-02-19
農業部淡水魚類種質資源與生物技術重點開放實驗室開放課題LFB20070609資助
嚴太明(1972—), 男, 四川萬源人; 理學博士; 主要研究方向為魚類生態學; E-mail: yantaiming@sina.com
何智, E-mail: hezhisicau@aliyun.com
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
