鳙Sox基因克隆及序列進化分析
2014-03-29 01:44:45郭穩杰俞小牧童金茍
水生生物學報 2014年4期
郭穩杰俞小牧童金茍
(1. 中國科學院水生生物研究所, 武漢 430072; 2. 中國科學院大學, 北京 100049)
鳙Sox基因克隆及序列進化分析
郭穩杰1,2俞小牧1童金茍1
(1. 中國科學院水生生物研究所, 武漢 430072; 2. 中國科學院大學, 北京 100049)
利用簡并引物SoxN和Sox9在鳙基因組DNA中進行PCR擴增和產物克隆測序, 并對序列進行同源性比較和系統進化分析。結果表明本文鑒定出鳙15個Sox基因HMG盒序列, 分別屬于SoxB、SoxC和SoxE組, 依據斑馬魚同源基因將其分別命名為Sox1a、Sox1b、Sox2、Sox3、Sox4a、Sox4b、Sox9a、Sox9b、Sox10、Sox11b、Sox12、Sox14a、Sox14b、Sox19和Sox21a。基于鳙和斑馬魚 Sox1、Sox4和Sox9 基因核苷酸序列構建的系統進化樹顯示這3個Sox基因的復制時間發生在鳙和斑馬魚的分化之前, 結果支持了魚類特異的基因組復制假說。以Sox1a、Sox1b和Sox4基因為分子鐘標記構建系統進化樹探討鳙和斑馬魚的分化時間, 結果顯示, 同屬于鯉科魚類的鳙(鯉亞科)和斑馬魚(魚丹亞科)在原始的魚丹亞科魚類中存在一個共同祖先, 大約出現在63.7百萬年前。研究結果為進一步研究魚類Sox基因復制和基因組進化等問題提供了重要參考資料。
鳙; Sox基因; 基因組加倍; 系統進化樹; 分化時間
Sox(Sry-related HMG-box)是一類與哺乳動物性別決定基因Sry (Sex determining region of Y chromosome)相關基因構成的基因家族。該家族成員都含有一個保守的HMG (High mobility group) 盒, 其編碼產物能特異性地識別和結合 DNA序列, 使DNA發生彎曲, 是一類重要的轉錄調控因子[1]。Sox基因廣泛參與神經系統發育、血細胞生成、晶狀體發育以及性別決定和分化等重要的生物學過程。例如, SOX9與SRY共同作用調控睪丸的發育[2]; Sox1、Sox2、Sox3、Sox4和Sox11等參與神經系統的發育調控[3—5]。目前, Bowles, et al.[6]根據HMG盒的相似性, 將Sox基因分成A-J 10個亞族。
由于在進化過程中基因加倍等原因, 魚類和哺乳動物的Sox基因家族成員有所差別。例如, Sox1、Sox4、Sox6、Sox8、Sox9、Sox11、Sox14和 Sox21在人類和小鼠基因組中只存在一個拷貝; 而在斑馬魚中, 這些基因被證實有兩個旁系同源基因[7](Paralogous genes)。關于魚類 Sox基因復制的原因尚無定論, 主要包括全基因組復制和基因片段復制兩種觀點[8]。目前許多研究都支持 Sox基因加倍是全基因組復制的結果[9,10]。
研究表明利用堿基同義突變速率和分子鐘模型構建的系統進化樹, 能夠估計物種間的大致分化時間[11,12]。由于Sox基因廣泛存在于脊椎動物基因組中, 且易于克隆, 成為物種系統進化研究非常重要的材料。鯉科魚類(Cyprinidae)是現有魚類中最大的一個科, 也是生物學家探索水生動物進化極好的代表群體[13]。目前, 已有些研究利用 Sox基因估算鯉科魚類物種間分化時間[9,10]。鳙(Aristichthys nobilis)屬于鯉形目鯉科鰱亞科, 是新近形成的一個物種[14],對其Sox基因HMG盒進行克隆和序列進化分析, 可為探索魚類Sox基因加倍機制和鯉科魚類系統進化研究提供很好的分子資料。
1 材料與方法
1.1 材料和DNA提取
實驗樣本采自武漢漲渡湖漁場, 剪取一尾鳙少量鰭條組織浸泡在 95%乙醇中, 然后置于 4℃下保存備用。采用經典的酚氯仿法[15]提取基因組DNA。
1.2 基因擴增、克隆和測序
針對脊椎動物Sox基因HMG盒保守的氨基酸序列, 設計兩對簡并引物 SoxN和 Sox9[16], 由上海生工生物工程技術服務有限公司合成。引物對SoxN序列為5′-ATGAAYGCNTTYATGGTNTGG-3′和5′-GGNCGRTAYTTRTARTCNGG-3′, 引物對Sox9序列為 5′-ATGAAYGCSTTYATGGTITGG-3′和 5′-GTCIGGGTGRTCYTTCTTRTGYTG-3′。PCR擴增反應體系總體積為50 μL, 包括160—200 ng的基因組DNA,上下游引物(5 μmol/L)各 2 μL, 1.6 μL dNTP (2.5 mmol/L), 0.4 μL Taq DNA酶(5 U/μL)(Takara), 1.25 μL 10×Buffer, 最后補足滅菌雙蒸水至終體積。PCR反應條件如下: 94℃預變性4min; 擴增35個循環, 每個循環 94℃變性 1min、53℃退火 1min、72℃延伸1min; 72℃終延伸10min。PCR擴增產物經1%的瓊脂糖凝膠檢測后, 并用 Biospin膠回收試劑盒(BioFlux, China)純化回收擴增產物。將回收產物連接到 pMD18-T克隆載體上(Takara, Japan), 再轉化到大腸桿菌DH5α, PCR檢測陽性克隆, 交由北京諾賽基因組研究中心有限公司測序。
1.3 序列進化分析
本文分析中所用到的斑馬魚(Danio rerio)、紅鰭東方 鲀(Temmincket Schlegel)、海鱸(Dicentrarchus labrax)一些Sox基因序列的來源見表1。
Sox基因的鑒定 將引物序列從測得的基因序列中去除, 然后用 Clustalx 1.81[17]軟件進行序列比對, 根據比對結果選取有一定變異的一組核酸序列作基因鑒定分析。將篩選出的核酸序列用MEGA 4.0[18]軟件翻譯成氨基酸序列, 在 GenBank數據庫中進行同源性搜索, 鑒定Sox基因。
系統進化樹分析 采用MEGA 4.0軟件, 用斑馬魚Sox基因和所有鑒定出的鳙Sox基因氨基酸序列一起進行最小進化法(Minimum-evolution method, ME)分析, 并以人的TCF7基因(MN_201632)為外類群。
Sox基因復制時間的推算 由于密碼子具有兼并性, 通常氨基酸第 3個密碼子堿基置換不改變氨基酸序列, 其替換速率能全面地反映基因突變率,可用于編碼基因的進化分析[11,19]。本研究以鳙和斑馬魚Sox1、Sox4和Sox9基因核苷酸序列為材料, 根據第3位密碼子堿基置換速率, 利用ME法構建系統進化樹, 推算Sox基因的復制時間。

表1 本研究用于序列分析的Sox基因Tab. 1 Sox genes used for sequence analysis in the study
鳙與斑馬魚分化時間估算 本研究以鳙、斑馬魚、紅鰭東方 鲀和海鱸Sox1a、Sox1b和Sox4基因數據集為分子鐘標記, 根據第 3位密碼子堿基置換速率, 利用非加權組平均法(UPGMA method)構建系統進化樹, 估算鳙和斑馬魚的分化時間。
2 結果
2.1 基因擴增、克隆與測序
在鳙基因組DNA中, SoxN引物擴增出一條約為220 bp的條帶; Sox9引物擴增出大小不等的4條帶, 分別為200、300、700和1200 bp左右。對這些擴增產物進行克隆, 經菌落PCR鑒定, 共挑選60個陽性克隆進行測序。
2.2 序列分析
鳙Sox基因命名及與斑馬魚蛋白的系統進化樹分析 經Clustal序列比對和Blast同源性比較, 共鑒定出15個鳙Sox基因, 分別命名為: Sox1a、Sox1b、Sox2、Sox3、Sox4a、Sox4b、Sox9a、Sox9b、Sox10、Sox11b、Sox12、Sox14a、Sox14b、Sox19和 Sox21a (GenBank登錄號為: KC883624-KC883629, KC867495,KC867494, KC867496, KC883630-KC8836 35)。
以鳙 Sox基因氨基酸序列與斑馬魚相應的 Sox蛋白以ME法構建系統進化樹(圖1)。結果顯示, 本文所得到的15個鳙Sox基因歸屬于SoxB、SoxC和SoxE三個亞族, 其中引物 SoxN擴增所得到的鳙Sox基因屬于SoxB和SoxC, 引物Sox9擴增得到的鳙Sox基因屬于SoxB和SoxE組。
Sox基因復制時間的推算 為了推算Sox基因的復制時間, 以鳙和斑馬魚 Sox1、Sox4和 Sox9基因的核苷酸序列第 3位密碼子堿基置換速率, 利用ME法構建了系統進化樹(圖2)。系統進化樹結果顯示, Sox1、Sox4和Sox9基因的復制時間發生在鳙和斑馬魚的分化之前。
鳙與斑馬魚的分化時間 利用鳙、斑馬魚、紅鰭東方 鲀和海鱸Sox1a、Sox1b和Sox4基因數據集第3位密碼子堿基置換速率, 采用UPGMA法構建系統進化樹, 以期根據這些基因的進化推測物種間的分化時間。以斑馬魚與紅鰭東方 鲀的分化時間(290百萬年前)為標定點[20], 利用以 Sox1a、Sox1b及 Sox4基因為分子鐘標記所構建的系統進化樹(圖 3), 推測鳙(鯉形目, 鯉科, 鳙屬)與斑馬魚(鯉形目,鯉科, 魚丹屬)的分化發生在63.7百萬年前。
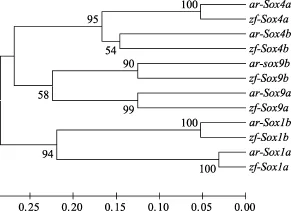
圖2 利用鳙和斑馬魚Sox1、Sox9和Sox11基因第3位密碼子堿基替換速率構建系統進化樹Fig. 2 Phylogenetic tree of Sox1, Sox9 and Sox11 genes from bighead carp and zebrafish using third-codon position substitution rates
3 討論
3.1 鳙Sox基因HMG盒序列克隆用于Sox基因家族進化的研究
Sox基因家族成員最顯著特征是含有一個保守的HMG盒, 可以根據HMG盒序列進行Sox基因的鑒定及其家族進化的研究。利用SoxN和Sox9兩對簡并引物對鳙基因組 DNA進行擴增, 為了保證測序結果的可信度, 本文鑒定出的每個 Sox基因序列至少是兩個不同克隆測序結果。測得的序列經blast比對分析, 我們共鑒定出15個鳙Sox基因, 分屬于SoxB、SoxC和SoxE。有趣的是, 歸屬于SoxB亞族的Sox19基因, 在哺乳動物中不存在其同源基因[21],而在斑馬魚[22]和 紅鰭東方 鲀[23]等真骨魚中則存在其同源基因。Sox19基因在哺乳動物基因組中的缺失可能是在進化過程中, 哺乳動物丟失了該基因,也可能是真骨魚類通過基因復制逐漸由某一基因的復制子演化成Sox19基因。Koopman, et al.[23]認為Sox19基因很可能是由Sox3基因的一個復制子進化而來, 圖1中Sox3和Sox19的進化關系可能是這種觀點的一個佐證。鳙這些Sox基因的鑒定將有助于硬骨魚類Sox基因家族的進化研究。
3.2 Sox基因的復制及其復制時間
在魚類基因組研究中已經發現基因復制是非常常見的現象, 但重復基因在不同的物種中有不同的命運, 主要包括選擇性丟失或協同進化產生新的功能[24,25]。Blast同源性比較和進化樹分析表明, Sox1、Sox4、Sox9和Sox14在鳙基因組中發生了基因復制。其中Sox1、Sox4和Sox9的復制并沒有改變其氨基酸序列, 說明這3個Sox基因在進化過程中受到很大的選擇壓力, 可能在鳙個體發育過程中發揮重要的調控作用。如果在輻鰭魚進化歷程中魚類特異的全基因組復制事件存立的話, 那么這次事件一定發生在鳙和斑馬魚分化之前。然而, 關于魚類特異性的Sox基因復制, 是全基因組復制還是Sox基因片段復制的結果, 迄今還沒有定論。本文的Sox基因復制時間估算結果表明, Sox1、Sox4和Sox11基因的復制時間確實發生在鳙和斑馬魚的分化之前(圖 2), 這也在一定程度上支持了“魚類特異性的基因組復制”學說[26]。
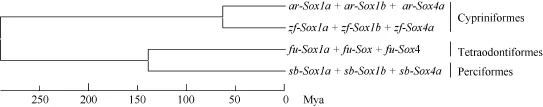
圖3 利用鳙、斑馬魚、紅鰭東方 鲀和海鱸Sox1a、Sox1b和Sox4基因數據集第3位密碼子堿基替換速率構建系統進化樹Fig. 3 Phylogenetic tree of concatenated dataset of Sox1a, Sox1b and Sox4 genes from bighead carp, zebrafish, fugu and sea bass using third-codon position substitution rates
3.3 鳙和斑馬魚的分化時間
鳙是鯉科魚類進化過程中新近形成的一個物種,對其Sox基因HMG盒進行克隆和序列分析, 可為研究魚類Sox基因進化歷程提供較好的分子信息。陳宜瑜等[14]?通過對現有魚類化石資料的觀察和鑒定認為這些化石大都屬于原始的 鲃亞科和魚丹亞科, 且都出現在距今65百萬年至距今26百萬年的老第三紀。由于地質的一系列變化帶來溫暖地帶向南退縮,生活在熱帶和亞熱帶的原始魚丹亞科不適應氣候變化在北方幾近滅絕, 從而派生出適應寒冷氣候的雅羅魚亞科魚類成為鯉科魚類重要的組成成分。隨著青藏高原急劇隆升, 形成了長江中下游河流縱橫交錯的復合生態系統, 由適應較冷氣候環境的原始雅羅魚亞科, 逐步派生出 鲌亞科和鰱亞科。綜上所述,鳙、草魚和斑馬魚在原始魚丹亞科中存在一個共同的祖先。本研究以Sox1a、Sox1b及Sox4基因為分子鐘標記, 推測鳙(鯉形目, 鯉科, 鰱亞科, 鳙屬)與斑馬魚(鯉形目, 鯉科, 魚丹亞科, 魚丹屬)共同的祖先大約存在于63.7百萬年前, 正好處于老第三紀的古新世(Paleocene)(距今65百萬年至53百萬年)。本文的這個推算結果與Zhong, et al.[10]研究草魚(鯉形目, 鯉科, 雅羅魚亞科, 草魚屬)和斑馬魚的分化時間(63百萬年前左右)一致, 從而從基因進化的角度證實了陳宜瑜等[14]對鯉科魚類物種進化的推斷。
[1] Rehberg S, Lischka P, Glaser G, et al. Sox10 is an active nucleo-cytoplasmic shuttle protein and shuttling is crucial for Sox10-mediated transactivation [J]. Molecular and Cellular Biology, 2002, 22: 5826—5834
[2] Zanaria E, Muscatelli F, Bardoni B, et al. An unusual member of the nuclear hormone receptor superfamily responsible for X-linked adrenal hypoplasia congenita [J]. Nature, 1994, 372: 635—641
[3] Uwanogho D, Rex M, Cartwright E J, et al. Embryonic expression of the chicken Sox2, Sox3 and Sox11 genes suggests an interactive role in neuronal development [J]. Mechanisms of Development, 1995, 49(1): 23—36
[4] Collignon J, Socknathan S, Hacker A, et al. A comparison of the properties of Sox3 with Sry and two related genes Sox1 and Sox2 [J]. Development, 1996, 122(2): 509
[5] Cheung M, Abu-Elmagd M, Clevers H, et al. Roles of Sox4 in central nervous system development [J]. Molecular Brain Research, 2000, 79(1): 180—191
[6] Bowles J, Schepers G, Koopman P. Phylogeny of the Sox family of developmental transcription factors based on sequence and structural indicators [J]. Development Biology, 2000, 227(2): 239—255
[7] Okuda Y, Yoda H, Uchikawa M, et al. Comparative genomic and expression analysis of group B1 sox genes in zebrafish indicates their diversification during vertebrate evolution [J]. Developmental Dynamics, 2006, 235(3): 811—825
[8] Vandepoele K, De Vos W, Taylor J S, et al. Major events in the genome evolution of vertebrates: paranome age and size differ considerably between ray-finned fishes and land vertebrates [J]. Proceedings of the National Academy of the Sciences ot the United States of America, 2004, 101(6): 1638—1643
[9] Zheng J, Long L, Hu J, et al. Isolation and sequence analysis of Sox genes in the red crucian carp (Carassius carassius, red variety) [J]. Russian Journal of Genetics, 2008, 44(11): 1325—1330
[10] Zhong L, Yu X M, Tong J G. Sox genes in grass carp (Ctenopharyngodon idella) with their implications forgenome duplication and evolution [J]. Genetics Selection Evolution, 2006, 38: 673—687
[11] Nei M, Rogozin L B, Piontkivska H, et al. Purifying selection and birth-and-death evolution in the ubiquitin gene family [J]. Proceeding of the National Academy of the Sciences, 2000, 97(20): 10866—10871
[12] Nei M, Kumar S. Molecular Evolution and Phylogenetics [M]. New York: Oxford University Press. 2000, 203—206
[13] 韓亞偉, 童超波, 何舜平, 等. 鯉科魚類系統進化過程中SINEs的插人事件. 自然科學進展, 2007, 17(10): 1345—1351
[14] Chen Y Y, Cao W X, Zheng C Y. Ichthyofauna of the Zhujiang river with a discussion on zoogeographical divisions for freshwater fishes [J]. Acta Hydrobiologica Sinica, 1986, 10(3): 228—236 [陳宜瑜, 曹文宣, 鄭慈英.珠江的魚類區系及其動物地理區劃的討論. 水生生物學報, 1986, 10(3): 228—236]
[15] Taggart J B, Hynes R A, Prod?uhl P A, et al. A simplified protocol for routine total DNA isolation from salmonid fishes [J]. Journal of Fish Biology, 1992, 40(6): 963—965
[16] Galay-Burgos M, Llewellyn L, Mylonas C C, et al. Analysis of the Sox gene family in the European sea bass (Dicentrarchus labrax) [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2004, 137(2): 279—284
[17] Thompson J D, Gibson T J, Plewniak F, et al. The Clustal_X Windows interface: Flexible strategies for multiple sequences alignment aided by quality analysis Tools [J]. Nucleic Acids Research, 1997, 25(24): 4876—4882
[18] Tamura K, Dudley J, Nei M, et al. Mega 4: Molecular evolutionary genetics analysis (Mega) software version 4.0 [J]. Molecular Biology and Evolution, 2007, 24(8): 1596—1599
[19] Hughes A L. Adaptive Evolution of Genes and Genomes [M]. New York: Oxford University Press. 1999, 15—37
[20] Kumazawa Y, Yamaguchi M, Nishida M. The Biology of Biodiversity [M]. Hong Kong: Springer-Verlag. 1999, 35—52
[21] Hett A K, Ludwig A. SRY-related (Sox) genes in the genome of European Atlantic sturgeon (Acipenser sturio) [J]. Genome, 2005, 48(2): 181—186
[22] Vriz S, Lovell-Badge R. The zebrafish Zf-Sox 19 protein: A novel member of the Sox family which reveals highly conserved motifs outside of the DNA-binding domain [J]. Gene, 1995, 153(2): 275—276
[23] Koopman P, Schepers G, Brenner S, et al. Origin and diversity of the Sox transcription factor gene family: Genome-wide analysis in Fugu rubripes [J]. Gene, 2004, 328: 177—186
[24] Postlethwait J, Amores A, Cresko W, et al. Subfunction partitioning, the teleost radiation and the annotation of the human genome [J]. Trends in Genetics, 2004, 20(10): 481—490
[25] Maere S, De Bodt S, Raes J, et al. Modeling gene and genome duplication in eukaryotes [J]. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(10): 5454—5459
[26] Zhou L, Wang Y, Gui J F. Fish-specific genome duplication [J]. Zoological Research, 2006, 27(5): 525—532 [周莉, 汪洋, 桂建芳. 魚類特異的基因組復制. 動物學研究, 2006, 27(5): 525—532]
CLONING AND SEQUENCE EVOLUTION ANALYSIS OF SOX GENES IN BIGHEAD CARP (ARISTICHTHYS NOBILIS)
GUO Wen-Jie1,2, YU Xiao-Mu1and TONG Jin-Gou1
(1. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China)
To amplify Sox genes from bighead carp genome, two pairs of degenerate primers (SoxN and Sox9) were designed for PCR amplification that were utilized for sequencing and analysis of sequence homology and phylogenetic relationships. The results showed that 15 distinct Sox genes encoding the HMG domains were identified in bighead carp, which were assigned to group B, C and E. According to their homology to orthologs of zebrafish, 15 Sox genes were designated as Sox1a, Sox1b, Sox2, Sox3, Sox4a, Sox4b, Sox9a, Sox9b, Sox10, Sox11b, Sox12, Sox14a, Sox14b, Sox19 and Sox21a, respectively. Phylogenetic tree of Sox1, Sox4 and Sox9 nucleotide sequences indicated that the duplication of these three Sox genes occurred before the divergence of bighead carp and zebrafish, and this observation supported the“fish-specific whole-genome duplication” theory. Using Sox1a, Sox1b and Sox4 as molecular clock markers in phylogenetic analysis, the estimation of the divergence time between bighead carp and zebrafish demonstrated that in original Danioninae, a common ancestor appeared approximately 63.7 million years ago for bighead carp and zebrafish, both of which belong to Cyprinidae. This study would provide important information for further studies of Sox gene replication and genome evolution in fish.
Aristichthys nobilis; Sox gene; Genome duplication; Phylogenetic tree; Divergenc time
Q781
A
1000-3207(2014)04-0664-05
10.7541/2014.94
2013-04-23;
2014-01-09
公益性(農業)行業科研項目(200903045); 國家自然科學基金項目(31272647)資助
郭穩杰(1985—), 男, 湖北黃梅縣人; 博士研究生; 主要從事魚類遺傳與基因組學研究。E-mail: chaliangle12@163. com
童金茍, E-mail: jgtong@ihb.ac.cn
