飼料魚粉、菜粕比例對異育銀鯽生長和飼料利用的影響
2014-03-29 01:44:44劉曉慶朱曉鳴韓冬金俊琰楊云霞解綬啟
水生生物學報 2014年4期
關鍵詞:實驗
劉曉慶朱曉鳴韓 冬金俊琰楊云霞解綬啟
(1. 安徽大學資源與環境工程學院, 合肥230601; 2. 中國科學院水生生物研究所淡水生態與生物技術國家重點實驗室, 武漢 430072)
飼料魚粉、菜粕比例對異育銀鯽生長和飼料利用的影響
劉曉慶1,2朱曉鳴2韓 冬2金俊琰2楊云霞2解綬啟2
(1. 安徽大學資源與環境工程學院, 合肥230601; 2. 中國科學院水生生物研究所淡水生態與生物技術國家重點實驗室, 武漢 430072)
通過在飼料中配制不同魚粉和菜粕比例, 探討利用菜粕蛋白替代魚粉對異育銀鯽(Carssius auratus gibelio)的生長和飼料利用的影響。飼料中菜粕蛋白替代魚粉蛋白的比例分別為 0、20%、40%、60% 和80%。實驗結果表明, 當飼料中菜粕蛋白比例高于 40%時, 異育銀鯽的生長顯著下降(P<0.05), 對攝食率影響不明顯(P>0.05)。在80%替代水平下, 干物質和蛋白質的消化率最低, 能量消化率在40%替代水平最低。飼料轉化效率在20%蛋白替代后明顯下降(P<0.05)。蛋白質和能量儲積率在40%蛋白替代后明顯下降(P<0.05)。在40%菜粕蛋白下, 魚體能量和脂肪含量最高。魚體蛋白質在60% 和80% 菜粕蛋白時最低, 脂肪在20% 菜粕蛋白時最高。因此飼料菜粕蛋白的水平不超過20%(干物質含量約17%)不會影響異育銀鯽的生長和飼料轉化。
菜粕; 魚粉; 生長; 飼料利用; 異育銀鯽
魚粉是優質的飼料蛋白源, 也是飼料原料中價格較高的原料。魚粉資源的緊缺也導致其價格在不斷上升[1]。因此, 飼料企業和營養學家一直致力于尋找降低飼料中魚粉用量的方法, 其中利用一部分植物蛋白是重要的手段之一[2]。豆粕常被作為第一替代蛋白源, 但是隨著中國飼料產業對豆粕的依賴性加強, 豆粕的價格不穩定且隨著養殖對飼料需求量升高逐年上漲。我國油菜種植面積較大, 年產菜粕600余萬噸, 且產量較為穩定, 菜粕含粗蛋白約35%—40%, 還富含維生素和礦物鹽, 是一種重要的植物蛋白源[3,4]。但是因其含有多種抗營養因子和毒素, 限制了菜粕在魚類飼料中的使用量。國內外現有研究表明, 影響魚類對菜粕利用的因素通常包括: (1) 氨基酸不平衡, 尤其是賴氨酸的缺乏[5]; (2) 抗營養因子如單寧酸、芥子油甙等, 可影響氨基酸的利用、消化酶的活性、礦物鹽的利用等[6]; (3) 纖維含量高, 會降低礦物質的利用率, 減少腸道內容物通過腸道的時間[7,8]; (4) 適口性低, 降低魚類攝食[9,10]。大多數研究表明在日糧中添加30%以上的菜粕會降低魚類生長和飼料利用[11—13], 但是也有一些學者報道魚類可以攝食更高水平的飼料菜粕[14,15]。盡管菜粕中的抗營養因子等限制了其在水產飼料中的使用, 并對魚類生長性能和飼料的消化吸收產生不良影響, 但是由于養殖種類、規格、養殖方式、實驗條件等的不同, 導致菜粕在各種魚類飼料中的使用量也不相同。
異育銀鯽(Carassius auratus gibelio)作為我國一種重要的經濟養殖對象, 由于它生長迅速, 肉質鮮美, 深受大眾歡迎, 在中國水產養殖業中具有重要地位[16]。菜粕是異育銀鯽飼料的常用原料, 但是其適宜添加量并沒有報道。我們實驗室此前研究了較高水平的不同植物蛋白源(包括菜粕)替代魚粉對異育銀鯽的生長和飼料利用造成的影響, 結果表明與魚粉對照組相比, 較高含量的菜粕不影響異育銀鯽的攝食但是降低了其生長[17]。本實驗的目的是研究飼料中不同水平菜粕替代魚粉對異育銀鯽生長和飼料利用的影響, 以期為菜粕在水產飼料中的應用提供理論參考。
1 材料與方法
1.1 養殖條件和實驗魚
異育銀鯽來源于中國科學院水生生物研究所關橋漁場。在生長實驗開始 4周之前, 將實驗魚轉入水族箱。在適應階段, 用商品飼料(蛋白質含量35%)每天飽食投喂兩次。在實驗開始前一周, 將實驗飼料按同等比例混合并飼喂實驗魚。
實驗系統為15個平底玻璃鋼水族箱(直徑: 60 cm,容積: 100 L)組成的室內循環水系統。每個水族箱的中央排水管都連有一個用來排除糞便排泄物沉積管。所有水族箱的水都匯集到一個容積為4 m3的生物過濾池。系統中的水保持充氣, 水體交換速率約為 3 L/min。在實驗過程中, 水體溶氧保持在為5 mg/L左右, pH約為6.8, 氨氮含量低于0.5 mg/ L,水溫在26.5—29.5℃(平均溫度為28.5℃), 光照周期是12L︰12D, 光照時間是08:00到20:00。
1.2 實驗飼料
實驗設計5種等氮(蛋白質水平為38%)等能(能值為14 kJ/g)的飼料, 以菜粕蛋白分別替代0(對照)、20%、40%、60%和80%的飼料蛋白質, 其他部分的蛋白質由魚粉提供, 添加1% Cr2O3作為測定消化率的指示劑, 每種飼料 3個重復。飼料配方及化學成分如表1所示。飼料原料過40目后充分混勻, 經飼料機制成直徑為1 mm的顆粒, 60℃烘干后, 置于–4℃冰箱中備用。
在實驗開始時, 選取經過24h饑餓而體質健壯、規格整齊的個體(平均每尾重3.5 g), 隨機放入15個魚缸, 每缸放養30尾, 進行為期8周的生長實驗。初始取樣為隨機取15尾魚隨機分為3組, 每組5尾。初始樣品在 120℃經過高壓蒸汽處理后搗碎混勻,并在70℃烘干, 用于初始體成分分析。
在實驗期間, 每天飽食投喂兩次(09: 00和 14: 30)。投喂1h后用虹吸法收集殘餌, 60℃烘干稱重。每天兩次從排泄物通道(沉降室)收集外形完整的糞便并60℃烘干。在實驗結束后, 在沒有魚的缸中分別投放對應的已稱重飼料, 1h后回收飼料, 60℃烘干后稱重, 計算飼料的溶失率, 用來校正實際攝食量。
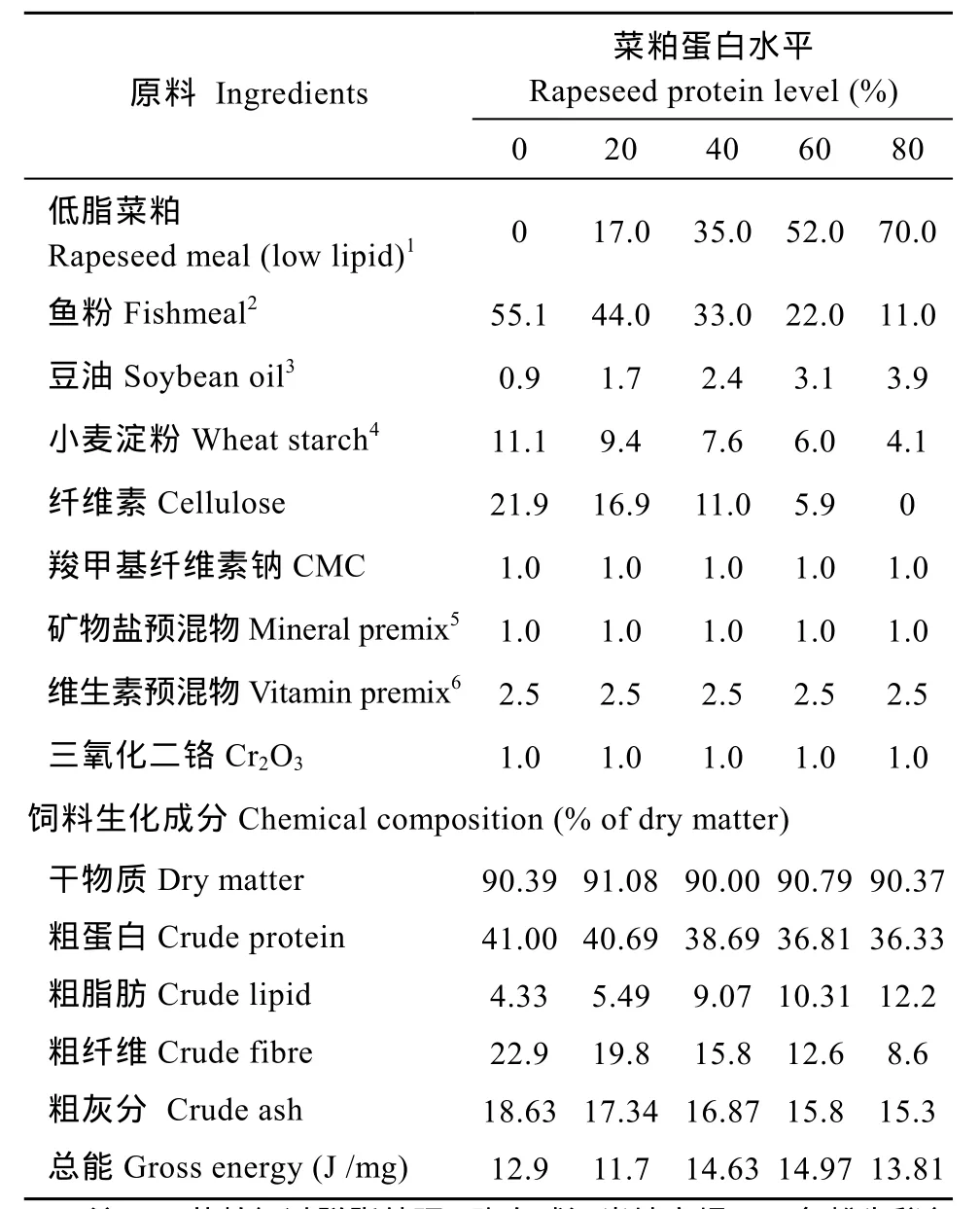
表1 飼料組成與飼料的化學成分(%干物質)Tab. 1 The formulation and chemical composition of the experimental diets (% dry matter)
1.3 實驗取樣和樣品分析
在實驗結束時, 魚體饑餓1d后分別稱重。每缸取5尾魚用于生化分析。在實驗中測定的生化成分包括飼料和實驗魚體樣品的干物質、粗蛋白、粗脂肪、灰分和總能。以上參數的測定均參照 AOAC (1984)的方法。干物質含量通過在105℃下烘烤至恒重測得。粗蛋白含量用凱氏定氮儀(2300Kjeltec Analyzer Unit, FOSS TECATOR, Sweden)測定, 粗脂肪用索氏抽提法(Soxtoc system HT6, Tecator, Haganas, Sweden)以乙醚為溶劑測定, 纖維含量是將樣品用0.5 mol/L H2SO4和0.5 mol/L NaOH抽提后干燥、灰化來測定, 灰分在馬福爐中 550℃下充分灼燒測得, 能值用 Phillipson能量計(Phillipsonmicrobomb calorimeter, Gentry Instruments Inc., Aiken, USA)測定。每個樣品測定兩個平行樣。飼料干物質、粗蛋白和能量的表觀消化率的測定是通過檢測飼料和糞便中三氧化二鉻以及營養物質和能量的含量計算得出[18]。
1.4 數據處理及統計分析
實驗魚的生長性能參數是計算公式的如下: 攝食率(FR, %BW/d)=100×攝食量/[(初始體重+終末體重)/2]/天數
特定生長率(SGR, %/d)=100×(ln魚體末重–ln魚體初重)/天數
飼料效率(FE, %)=100×(魚體末重–魚體初重)/攝食量
蛋白質儲積率(PRE, %)=100×(終末魚體蛋白質重–初始魚體蛋白質重)/蛋白質攝入量
能量儲積率(ERE, %)=100×(終末魚體能量-初始魚體能量)/能量攝入量
干物質表觀消化率(ADCd, %)= 100–100 ×飼料中Cr2O3的含量/ 糞便中Cr2O3的含量
蛋白質表觀消化率(ADCp, %) = 100–100 ×(飼料中Cr2O3含量×糞便蛋白質含量) / (糞便中 Cr2O3含量×飼料蛋白質含量)
能量表觀消化率(ADCe, %) = 100–100 ×(飼料中Cr2O3含量×糞便能量) / (糞便中Cr2O3含量×飼料能量)
使用SPSS 13.0對實驗數據作單因素方差分析(One-way ANOVA), 若差異顯著(P<0.05), 則進行Duncan’s多重比較檢驗組間的差異。
2 結果
2.1 生長性能
如表 2所示, 實驗魚的攝食率不受飼料中菜粕含量的影響(P>0.05), 但是飼料效率卻隨著菜粕的增加而降低, 即使是最低的菜粕替代組(20%), 異育銀鯽的飼料效率也顯著低于魚粉對照組(P<0.05)。當飼料中菜粕含量為 40%或更高時, 異育銀鯽的特定生長率、蛋白質儲積率和能量儲積率顯著降低(P<0.05)。
2.2 表觀消化率
如表 3所示, 與對照組相比, 只有最高替代組(80%替代組)的干物質和粗蛋白質的表觀消化率出現顯著下降(P<0.05)。40%和80%替代組的能量表觀消化率顯著低于其他各組(P<0.05)。
2.3 魚體成分
由表4可知, 40%替代組的全魚干物質含量顯著高于其他處理組(P<0.05), 且其他各組之間沒有顯著性的差異(P>0.05)。當替代水平為60%或更高時, 魚體蛋白質含量呈現出明顯的下降趨勢(P<0.05)。全魚脂肪含量最低組和最高組分別是對照組和 40%替代組, 并與其他組之間存在顯著性差異(P<0.05), 而其他各組之間則沒有顯著差異(P>0.05)。當替代水平達到60%或更高時, 全魚灰分含量明顯下降(P<0.05)。80%替代組的全魚能量顯著低于40%替代組(P<0.05),但是其他各組間沒有顯著性的差異(P>0.05)。
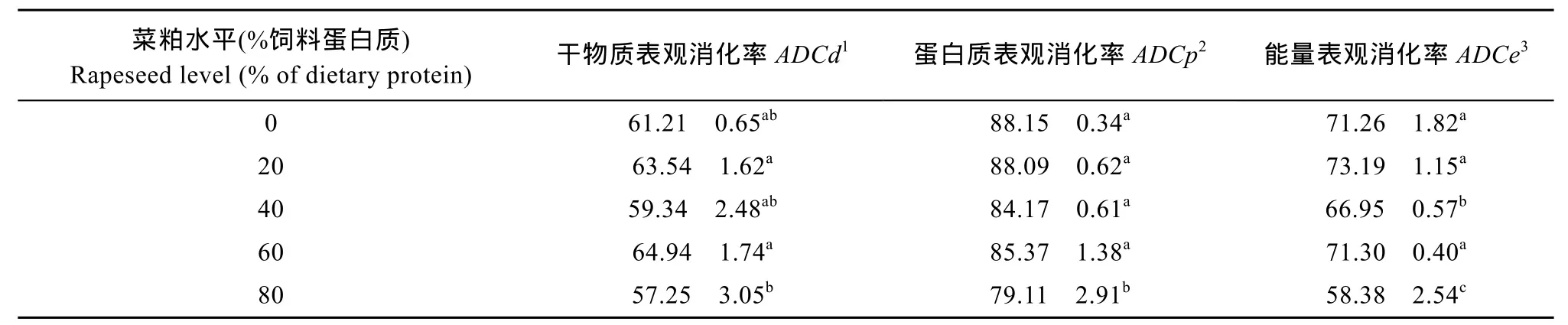
表2 飼料魚粉、菜粕水平對異育銀鯽消化率的影響(平均值±標準誤)Tab. 2 Effect of replacement of fish meal by rapeseed meal on the digestibility of Gibel carp (menas± SE)

表3 飼料魚粉、菜粕水平對異育銀鯽生長和飼料利用的影響(平均值±標準誤)Tab. 3 Effect of replacement of fish meal by rapeseed meal in the diet on the growth and feed utilisation of gibel carp (mean±SE)

表4 飼料魚粉、菜粕水平對異育銀鯽魚體組成的影響(平均值±標準誤)Tab. 4 Effect of replacement of fish meal by rapeseed meal on the body composition of gibel carp (menas± SE)
3 討論
實驗飼料蛋白質水平的設計值和實測值之間存在一些偏差, 引起這些偏差的原因可能是原料魚粉蛋白質含量的預算誤差, 但是設計值和實測值間的偏差對實驗結果結論沒有產生較大的影響。
根據實驗的結果, 飼料中菜粕基于蛋白質的替代水平低于 20%時(干物質中蛋白質水平為 17%),不會對異育銀鯽的生長產生大的負面影響。馬利對草魚(Ctenoparyngodon idellus)[19]、Davies對羅非魚(Oreochromis mossambicus Peters)[3]、Jackson對羅非魚(Sarotherodon mossambicus)[15]、Webster對斑點叉尾(Ictaluru punctatus)[20]以及 Yurkowski對虹鱒(Salmo gairdneri)[9]的研究中發現菜粕的添加水平不能高于 30%(飼料干物質中的含量)。這些研究結果均表明水產飼料中菜粕的添加量不宜過高。
近年來, 油菜籽新品種已經被選育出來并廣泛種植, 該品種油菜籽的籽實中含有低水平的抗甲狀腺復合物或芥子油甙, 油中含有低水平的芥子酸[21]。此外, 新型加工工藝已經開始用于鈍化菜粕中的抗營養因子, 從而降低了芥子油甙、植酸等抗營養因子[10,15]和多種難以消化且降低油菜籽優質蛋白質效價的碳水化合物對養殖魚類的影響, 從而改善的魚類對菜粕的利用。也有一些報道指出, 日糧中可以添加比較高水平的菜粕。Dabrowsski 和 Kozzlowska的研究表明在鯉日糧中菜籽粕替代 50%的魚粉可能不會對鯉產生任何負面影響[14]。Forster, et al.報道虹鱒(Oncorhynchus mykiss)攝食含 41.5%油菜籽蛋白濃縮物的日糧表現出和對照組相似的生長效果[22]。此外, Burel, et al.的實驗表明比目魚(Psetta maxima)攝食含 30%脫殼菜籽粕日糧可以達到與對照組相似的生長性能[23]。這些研究中菜粕添加比例的提高, 可能是由養殖魚類的食性和規格、實驗菜粕本身特性以及加工工藝的差異導致的。
在一些魚類中, 比如虹鱒(Oncorhynchus mykiss)[9]、比目魚(Psetta maxima)[23]和鮭(Oncorhynchus tshawytscha)[24], 菜粕會降低飼料的適口性和魚的攝食量, 最近的研究表明日糧中添加菜粕沒有影響到異育銀鯽的攝食量, 這說明異育銀鯽在適口性方面可以很好地接受菜粕。Reihold, et al.對虹鱒(oncorhynchus mykiss)[25]的研究以及我們對異育銀鯽的最新研究[17]也得出類似的結果。但也有相當多的研究表明菜粕替代魚粉后會顯著降低攝食率[26]。
很多研究表明, 在日糧中添加菜籽粕會降低日糧的消化率, 這可能是由于菜籽粕中高含量的纖維和多種抗營養因子[7,8]導致的。虹鱒對菜籽濃縮蛋白的蛋白質表觀消化率甚至高于低溫鳀魚粉, 干物質和能量表觀消化率和魚粉之間也沒有顯著性差異[8]。Burel, et al.[27]研究表明虹鱒對溶劑萃取型和高溫處理型兩種菜粕的蛋白質表觀消化率都接近魚粉, 大菱鲆對高溫處理型菜粕的蛋白質表觀消化率和魚粉接近, 而對溶劑萃取型菜粕的消化率低于魚粉。在本實驗中異育銀鯽的干物質和蛋白質表觀消化率只在高菜粕替代的飼料組降低。由于使用纖維素作為填充劑, 所以低菜粕水平飼料中粗纖維的含量比較高。這可能是造成異育銀鯽的消化率變化對飼料菜粕含量相對不敏感的原因。
在本實驗中, 異育銀鯽的飼料效率、蛋白質和能量的儲積率隨著飼料中菜粕替代水平的增加都呈現下降趨勢。這種現象和在大部分研究魚類對菜粕利用能力的報道一致[3,9,11,20,28]。這可能是由于菜粕中的氨基酸不平衡并且具有多種抗營養因子或者實驗魚攝食菜粕導致代謝損傷所造成的[29]。盡管異育銀鯽的干物質和蛋白質表觀消化率在中等菜粕替代水平的飼料中并沒有受到顯著地影響, 但一些研究結果表明飼料中較高的纖維含量和抗營養因子可能會降低異育銀鯽對飼料中某些氨基酸[30,31]和礦物質[32]的利用。但在本實驗中并沒有針對這些方面的設想進行具體的驗證。
總之, 在本實驗中菜粕基于蛋白質的替代水平低于 20%時(干物質中蛋白質水平為 17%)不會降低異育銀鯽的生長性能, 可以推薦在異育銀鯽飼料中進行使用。在飼料額外添加必需氨基酸、礦物質和其他微量營養素能否改善異育銀鯽對菜粕的利用能力值得進一步的研究。
[1] Brinker A, Reiter R. Fish meal replacement by plant protein substitution and guar gum addition in trout feed, part i: Effects on feed utilization and fish quality [J]. Aquaculture, 2011, 310(3—4): 350—360
[2] Gatlin D M, Barrows F T, Brown P. Expanding the utilization of sustainable plant products in aquafeeds: A review [J]. Aquaculture Research, 2007, 38(6): 551—579
[3] Davies S J, McConnell S, Bateson R I. Potential of rapeseed meal as an alternative protein source in complete diets for tilapia (Oreochromis mossambicus peters) [J]. Aquaculture, 1990, 87(2): 145—154
[4] Huang Y, Hu Y, Xiao T Y, et al. Influence of dietary canola meal levels on growth and biochemical indices in juvenile Mylopharvngodon piceus [J]. Acta Hydrobiologica Sinica, 2012, 36(1): 41—48 [黃云, 胡毅, 肖調義, 等. 雙低菜粕替代豆粕對青魚幼魚生長及生理生化指標的影響. 水生生物學報, 2012, 36(1): 41—48]
[5] Tacon A G J, Jackson A, Utilization of Conventional and Unconventional Protein Sources in Practical Fish Feeds [M]. In: Cowey C B, Mackie AMM, Bell J G (Eds.), Nutrition and Feeding in Fish. London: Academic Press. 1985, 119—145
[6] Higgs D A, McBride J R, Markert J R, et al. Evaluation of tower and candle rapeseed (canola) meal and bronowski rapeseed protein concentrate as protein supplements in practical dry diets for juvenile chinook salmon (Oncorhynchus tshawytscha) [J]. Aquaculture, 1982, 29(2): 1—31
[7] Hilton J W, Atkinson J L, Slinger S J. Effect of increased dietary fiber on the growth of rainbow trout (Salmo gairdneri) [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1983, 40(1): 81—85
[8] Hilton J W, Slinger S J. Digestibility and utilization of canola meal in practical-type diets for rainbow trout (Salmo gairdneri) [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1986, 43(6): 1149—1155
[9] Yurkowski M, Bailey J K, Evans R E, et al. Acceptability of rapeseed proteins in diets of rainbow trout (Salmo gairdneri) [J]. Journal of the Fisheries Board of Canada, 1978, 35(7): 951—962
[10] Eales J G. Effect of extrusion processing on the nutritive value of canola meal for chinook salmon (Oncorhynchus tshawytscha) in seawater [J]. Aquaculture nutrition, 1998, 4(2): 115—122
[11] Teskered?i? Z, Higgs D A, Dosanjh B S, et al. Assessment of undephytinized and dephytinized rapeseed protein concentrate as sources of dietary protein for juvenile rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture, 1995, 131(3): 261—277
[12] Takii K, Kita E, Nakamura M, et al. Evaluation of rapeseed protein concentration as protein source of diet for red sea bream [J]. Fisheriesscience-Tokyo, 1999, 65(2): 150—154
[13] Kissil G W, Lupatsch I, Higgs D A, et al. Dietary substitution of soy and rapeseed protein concentrates for fish meal, and their effects on growth and nutrient utilization in gilthead seabream Sparus aurata [J]. Aquaculture Research, 2000, 31(7): 595—601
[14] Dabrowski K, Krasnicki K, Kozlowska H. Rapeseed meal in the diet for common carp reared in heated waters [J]. Zeitschrift für Tierphysiologie Tierern?hrung und Futtermittelkunde, 1981, 46(1—5): 273—282
[15] Jackson A J, Capper B S, Matty A J. Evaluation of someplant proteins in complete diets for the tilapia sarotherodon mossambicus [J]. Aquaculture, 1982, 27(2): 97—109
[16] Xue M, Cui Y. Effect of several feeding stimulants on diet preference by juvenile gibel carp (Carassius auratus gibelio), fed diets with or without partial replacement of fish meal by meat and bone meal [J]. Aquaculture, 2001, 198(3): 281—292
[17] Xie S, Zhu X, Cui Y, et al. Utilization of several plant proteins by gibel carp (Carassius auratus gibelio) [J]. Journal of Applied Ichthyology, 2001, 17(2): 70—76
[18] Xie S, Cui Y, Yang Y, et al. Energy budget of nile tilapia (Oreochromis niloticus) in relation to ration size [J]. Aquaculture, 1997, 154(1): 57—68
[19] Ma L, Huang F, Wu J K, et al. Effects of different rapeseed meal levels on growth, serum biochemical indices and toxins residues in Ctenoparyngodon idellus [J]. Journal of Fisheries of China, 2005, 29(6): 798—803 [馬利, 黃峰, 吳建開, 等.不同菜粕水平對草魚生長、血清生化指標和毒素殘留的影響. 水產學報, 2005, 29(6): 798—803]
[20] Webster C D, Tiu L G, Tidwell J H, et al. Growth and body composition of channel catfish (Ictalurus punctatus) fed diets containing various percentages of canola meal [J]. Aquaculture, 1997, 150(1): 103—112
[21] Higgs D A, Fagerlund U H, McBride J R, et al. Protein quality of altex canola meal for juvenile chinook salmon (Oncorhynchus tshawytscha) considering dietary protein and 3, 5, 3′-triiodo-l-thyronine content [J]. Aquaculture, 1983, 34(3): 213—238
[22] Forster I, Higgs D A, Dosanjh B S, et al. Potential for dietary phytase to improve the nutritive value of canola protein concentrate and decrease phosphorus output in rainbow trout (Oncorhynchus mykiss) held in 11℃ fresh water [J]. Aquaculture, 1999, 179(1): 109—125
[23] Burel C, Boujard T, Kaushik S J, et al. Potential of plant-protein sources as fish meal substitutes in diets for turbot (Psetta maxima): Growth, nutrient utilisation and thyroid status [J]. Aquaculture, 2000, 188(3): 363—382
[24] Higgs D A, McBride J R, Markert J R, et al. Evaluation of tower and candle rapeseed (canola) meal and bronowski rapeseed protein concentrate as protein supplements in practical dry diets for juvenile chinook salmon (Oncorhynchus tshawytscha) [J]. Aquaculture, 1982, 29(1): 1—31
[25] Gomes E F, Corraze G, Kaushik S. Effects of dietary incorporation of a co-extruded plant protein (rapeseed and peas) on growth, nutrient utilization and muscle fatty acid composition of rainbow trout (Oncorhynchus mykiss) [J]. Aquaculture, 1993, 113(4): 339—353
[26] McNeill L, Bernard K, MacLeod M G. Food intake, growth rate, food conversion and food choice in broilers fed on diets high in rapeseed meal and pea meal, with observations on sensory evaluation of the resulting poultry meat [J]. British Poultry Science, 2004, 45(4): 519—523
[27] Burel C, Boujard T, Tulli F, et al. Digestibility of extruded peas, extruded lupin, and rapeseed meal in rainbow trout (Oncorhynchus mykiss) and turbot (Psetta maxima) [J]. Aquaculture, 2000, 188(3): 285—298
[28] Slawski H, Nagel F, Wysujack K, et al. Total fish meal replacement with canola protein isolate in diets fed to rainbow trout (Oncorhynchus mykiss W.) [J]. Aquaculture Nutrition, 2013,18(6): 10—18
[29] Tripath M K, Mishra A S. Glucosinolates in animal nutrition: a review [J]. Animal Feed Science and Technology, 2007, 132(2): 1—27
[30] Leslie A J, Summers J D. Amino acid balance of rapeseed meal [J]. Poultry Science, 1975, 54(2): 532—538
[31] Enami H R. A review of using canola/rapeseed meal in aquaculture feeding [J]. Fisheries and Aquatic Science, 2011, 6(1): 22—36
[32] Oberleas D, Harland B F. Nutritional agents which affect metabolic zinc status [J]. Progress in Clinical and Biological Research, 1977, 14(3): 11
EFFECTS OF DIETARY RATIO OF FISH MEAL AND RAPESEED MEAL ON THE GROWTH AND FEED UTILIZATION IN GIBEL CARP (CARASSIUS AURATUS GIBELIO)
LIU Xiao-Qing1,2, ZHU Xiao-Ming2, HAN Dong2, JIN Jun-Yan2, YANG Yun-Xia2and XIE Shou-Qi2
(1. College of Natural Resource and Environment Engineering, Anhui University, Hefei 230601, China; 2. State Key Laboratory of Freshwater Ecology and Biotechnology; Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China)
Effects of replacement of fish meal with rapeseed meal on the growth and feed utilisation were investigated in juvenile gibel carp (Carssius auratus gibelio). Three groups of fish were fed with each of five groups of diets supplemented with different concentrations of rapeseed: 0, 20%, 40%, 60% and 80% of total dietary protein. The experiment was carried out in a recirculation system at 26.5—29.5℃ (mean 28.5℃) for 8 weeks. The results showed that the growth rate of the fish decreased significantly when the rapeseed protein accounted for 40%, or higher, of the total dietary proteins. The feed intake was not affected by the concentration of rapeseed. The digestion rate of the dry matter and protein was significantly lower only when the concentration of rapeseed protein was 80% of the total dietary proteins. The digestion rate of energy was the lowest when rapeseed protein accounted for 40% of the total. Feed conversion efficiency decreased significantly when the dietary rapeseed protein concentration was 20% or higher. The protein and energy retention efficiency decreased when dietary rapeseed protein concentration was 40% or higher. The body energy and fat content reached the highest values when the fish was fed with the diet containing 40% rapeseed protein. The body protein level was lowest at the rapeseed protein concentrations of 60% and 80%, while the fat content was highest at 20%. Based on the data we suggest that the optimal concentration of rapeseed protein should be 20% in the diet for gibel carp.
Rapeseed meal; Fish meal; Growth; Feed utilization; Gibel carp
S965. 1
A
1000-3207(2014)04-0657-07
10.7541/2014.93
2013-04-16;
2013-12-20
飼料高效低耗加工技術研究與示范(201203015); 大宗淡水魚類產業技術體系-鯽魚營養與飼料崗位 CARS-46-19; 替代魚用飼料中魚粉的新蛋白源開發利用技術研究與示范(201303053)資助
劉曉慶 (1988—), 女, 安徽合肥人; 碩士; 主要從事魚類生理生態學研究。E-mail: sally.xiaoqing@163.com
朱曉鳴(1964—), 副研究員, 碩士生導師; E-mail: xmzhu@ihb.ac.cn
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
