傷寒沙門菌線性質粒pBSSB1的p17基因的生物學功能分析
2014-03-21 06:56:53朱云霞吳佳龔明玉侯書寧張綺思陳敏潔張海方
生物技術通報 2014年10期
朱云霞吳佳龔明玉侯書寧張綺思陳敏潔張海方,2
(1.江蘇大學 基礎醫學與醫學技術學院 醫學生物化學系,鎮江 212013;2.蘇州大學 附屬第二醫院 檢驗科,蘇州 215004)
傷寒沙門菌線性質粒pBSSB1的p17基因的生物學功能分析
朱云霞1吳佳1龔明玉1侯書寧1張綺思1陳敏潔1張海方1,2
(1.江蘇大學 基礎醫學與醫學技術學院 醫學生物化學系,鎮江 212013;2.蘇州大學 附屬第二醫院 檢驗科,蘇州 215004)
H:z66陽性傷寒沙門菌含有一特有的介導鞭毛單向相變換的線性質粒pBSSB1,其上第17號基因(p17)編碼一未知功能蛋白(P17)。利用λ-Red系統制備p17基因缺陷變異株,測定變異株和野生株的生長曲線,比較其生長差異,同時通過半固體LB培養基進行動力試驗比較其動力差異。利用PCR技術擴增獲得p17基因,構建其重組表達載體pET28a(+)-p17,通過鎳柱純化目的蛋白P17-His6,純化后蛋白作為抗原免疫兔子以制備多克隆抗體。結果顯示,制備了p17基因缺陷變異株,變異株的生長明顯遲緩于野生株(P<0.05),變異株的動力明顯減弱。構建了重組表達載體pET28a(+)-p17,純化獲得了高純度的目的蛋白P17-His6并制得相應的多克隆抗體。
傷寒沙門菌 線性質粒 pBSSB1 p17
傷寒沙門菌是一種重要的人類病原菌,且已成為原核生物基因表達調控研究的模式菌之一[1,2]。pBSSB1是H∶z66陽性傷寒沙門菌所特有的一線性質粒,該質粒是腸桿菌科中目前為止發現的首個線
性質粒,其介導了H∶z66陽性傷寒沙門菌鞭毛的單向相變換[3-6]。pBSSB1全長27 kb,共編碼33個基因,其中包括二相鞭毛素編碼基因fljBz66和fljA樣基因[3,7,8]。Baker等[3]報道pBSSB1可能的雙向復制起始點位于該質粒編碼的第17號基因(p17)的上游。基因p17經生物信息學分析和預測,其編碼蛋白P17含有特征性同源序列,該序列存在于參與調控DNA復制等生物學功能的DNA結合蛋白如ParA和Soj等蛋白中[9]。另外,蛋白P17具有與RepABC質粒編碼的RepA蛋白同源的ATP酶結構域[10]。但目前為止,關于基因p17的確切生物學功能還不甚清楚。
本研究通過制備p17基因缺陷變異株,測定缺陷株和野生株的生長曲線,對p17基因可能的生物學功能進行初步探討;另外,通過原核表達蛋白P17,并制備其相應的多克隆抗體,利用免疫印跡技術檢測抗體的特異性以及p17基因的表達情況,旨為深入研究p17基因的生物學功能奠定基礎。
1 材料與方法
1.1 材料
1.1.1 菌種和質粒 傷寒沙門菌GIFU10007由日本岐阜大學醫學院微生物室惠贈;質粒pKD46(溫度敏感型,含有受阿拉伯糖啟動子調控的gam,bet和exo基因,Apr)由北京軍事醫學科學院微生物流行病研究所惠贈;質粒pET28a(+)、E. coli DH5α、E. coli BL21(DE3)等均由本室保存。
1.1.2 主要試劑和儀器 DNA聚合酶pfu、DNA聚合酶rTaq、限制性核酸內切酶Nco I、Xho I、T4 DNA連接酶均購自寶生物工程有限公司(大連,TaKaRa),質粒提取試劑購自Axygen公司。其他試劑均為進口或國產分析純。Gene Pulsero Ⅱ電轉化儀(Bio-Rad),核酸紫外檢測儀(Eppendorf Biophotometer),PCR儀(AB I 2720),蛋白電泳儀(Bio-Rad),凝膠成像系統(Gene Genius BIO IMAGING SYSTEM),蛋白電泳儀(Bio-Rad),離心機,超聲破碎儀。
1.2 方法
1.2.1 引物設計 根據線性質粒pBSSB1的序列和pET28a(+)的序列信息,設計基因p17缺陷株制備、缺陷株驗證及構建p17編碼蛋白P17的重組表達載體pET28a(+)-p17等引物。制備p17基因缺陷株引物設計:p17基因的ORF全長641 bp,分別設計兩對引物L1-Kl/R1-Kr和L2/R2,擴增含有長度為79 bp的p17ORF上下游同源臂的卡那霉素抗性基因片段,用來敲除p17全部的ORF。p17缺陷株驗證引物V-L/V-R設計:V-L來自基因序列中上游同源臂外側一段16 bp的序列,V-R來自卡那霉素抗性基因中一段16 bp的序列。重組表達載體pET28a(+)-p17引物設計:在p17基因上下游設計引物PA/PB,在5'端分別加上Nco I及Xho I酶切位點用以酶切后連接pET28a(+)載體。引物均由上海生工生物技術服務有限公司合成,序列如表1所示 。
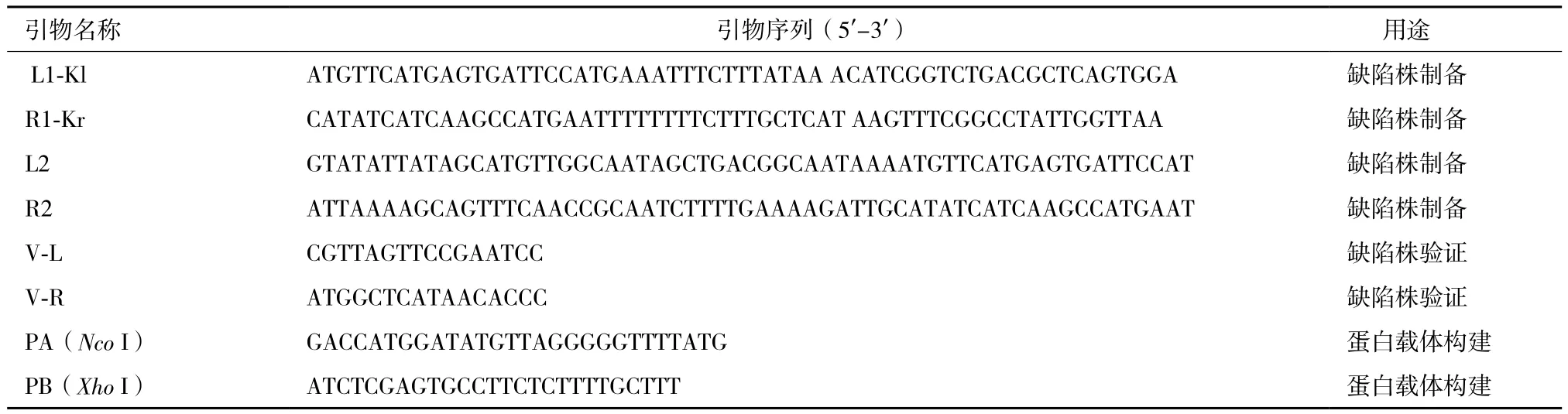
表1 引物序列
1.2.2 感受態細胞的制備及Red重組功能的誘導 將編碼Red重組系統的質粒pKD46電轉至傷寒沙門菌GIFU10007中,篩選陽性克隆,作為下一步轉化的宿主菌,即007-pKD46。挑取007-pKD46單菌落接種于2 mL LB培養基中37℃、250 r/min振搖過夜;然后1∶100轉接至20 mL等滲LB培養基中,
37℃、250 r/min振蕩培養至對數早期(OD600:0.4);加入L-阿拉伯糖(終濃度為1 mmol/L),誘導1 h后冰上靜置30 min,離心棄上清;沉淀用預冷的滅菌水漂洗3次,再用4 mL預冷的10%甘油洗1次,最后用80 μL預冷的10%甘油懸浮,每40 μL分裝于EP管中放于冰上備用。
1.2.3 基因p17缺陷株的制備 在參考文獻[11]的基礎上稍作改動,主要步驟如下:以環狀質粒pET28a(Kanr)為模板,用特異性引物L1-Kl/R1-Kr擴增出帶有p17基因上下游同源臂的卡那霉素抗性基因片段F1。然后以第1輪PCR產物為模板,用L2/R2做引物擴增出延長了同源臂的片段F2。用酚-氯仿抽提純化PCR產物F2。將007-pKD46的感受態細胞、電極杯和PCR產物均放于冰上,保持低溫狀態。打開電轉儀,調節參數:電阻200 Ω,電容25 μF,電壓2.0 kV。取2 μg PCR產物F2加入40 μL感受態細胞中,輕輕混勻,轉入電極杯中,迅速放到電轉儀上,啟動電擊。聽到“滴”聲后,取出電極杯,加入1 mL 37℃預溫的LB液體培養基輕輕混勻,轉移至EP管中,置于37℃金屬浴1 h。然后25℃、4 000 r/min,離心10 min,棄去900 μL上清液,將剩余液體重懸細菌后涂布于含有卡那霉素的LB平板37℃過夜,挑取長出的若干菌落通過驗證引物V-L/V-R進行PCR篩選,陽性轉化子最后經測序確認,即為p17基因缺陷株,命名為Δp17。
1.2.4 生長曲線測定 分別挑取傷寒沙門菌GIFU10007和Δp17單菌落于2 mL LB培養基中,37℃、250 r/min振蕩培養過夜;過夜細菌以1∶100轉接于新鮮等滲(pH 7.0)LB培養液中(野生株和缺陷株的OD600值一致),37℃、250 r/min振蕩培養,每隔1 h用分光光度計檢測OD600值并記錄,直至12 h。試驗經3次生物學重復后,以OD600值為縱坐標,培養時間為橫坐標制作生長曲線,比較野生株和p17缺陷株的生長情況。
1.2.5 動力試驗 分別挑取傷寒沙門菌GIFU10007和Δp17單菌落進行四區劃線,用滅菌后的牙簽挑取劃線后的大小均勻的野生株和Δp17單菌落接種于0.3%的半固體LB培養板上,置于37℃培養箱中培養18 h后觀察結果。
1.2.6 重組載體的構建與鑒定 參考文獻[12]的方法,用DNA膠回收試劑盒回收PCR 產物,NcoⅠ和Xho Ⅰ同時雙酶切PCR回收產物和表達載體pET28a(+),酶切產物用酚/氯仿抽提、乙醇沉淀純化后,用T4 DNA連接酶4℃過夜連接,連接產物用熱激法轉化至大腸桿菌DH5α。挑取若干連接產物轉化菌單菌落,分別增菌于含有卡那霉素的LB平板上,37℃培養6 h后,收集上述菌體于30 μL 雙蒸水中,加等體積的酚/氯仿漩渦震蕩,離心取上清15 μL上樣,1 %瓊脂糖凝膠電泳觀察質粒大小,篩選可疑的陽性克隆質粒。提取可疑的陽性克隆質粒,采用雙酶切和PCR方法鑒定陽性克隆,再通過DNA測序分析最終確認。陽性克隆質粒熱激法轉化至E. coli BL21(DE3)。
1.2.7 P17蛋白的表達 挑取新鮮的含重組質粒pET28a(+)-p17的大腸桿菌BL21(DE3)單菌落及含空質粒pET28a(+)的對照大腸桿菌BL21(DE3)單菌落分別置于2 mL LB 培養液中(卡那霉素終濃度50 μg/mL),37℃振搖過夜,次日以1∶100加入20 mL LB 培養液中37℃振搖2-3 h后長至OD600nm為0.5-0.8,加入IPTG 至終濃度分別為0.1 mmol/L,于20℃繼續振搖5 h進行誘導,離心取沉淀,4℃超聲破碎后離心取上清液進行SDS-PAGE。
1.2.8 P17蛋白的純化 在上述表達的優化條件下用1 000 mL LB培養基大批量誘導培養,離心收集菌體后,用PBS(pH7.2)洗滌3次,按照每克菌(濕重)加入15 mL 過柱緩沖液的比例重懸菌體,超聲破碎后離心收集上清液。以15 mL/h流速上樣于Ni柱中之后用20 倍柱體積的洗滌緩沖液沖洗Ni柱,移除未結合的蛋白;用洗脫緩沖液洗脫結合于Ni柱的蛋白。收集洗脫液,加入到處理后的透析袋中進行透析,透析后的蛋白取出離心取上清,并進行SDS-PAGE。
1.2.9 兔抗P17蛋白的多克隆抗體的制備 首次免疫取上述純化獲得的目的蛋白P17與弗氏完全佐劑1∶1乳化,按每只兔50 μg/mL抗原劑量,于背部皮下多點注射共計1 mL的方式進行免疫,之后每隔1周用弗氏不完全佐劑與純化獲得的目的蛋白P17以1∶1乳化,共免疫5次,末次免疫1周后頸動脈取血制備抗血清。抗血清經初步純化后獲得兔抗P17蛋白的多克隆抗體,分裝后于-80℃保存備用。用抗體與傷寒沙門菌GIFU10007全菌蛋白進行Western-
blotting以檢測抗體的特異性,具體方法見參考文獻[12]。
1.2.10 統計學分析 數據分析采用 SPSS 16.0 統計軟件。多組間均數比較采用單因素方差分析,以 P <0.05 為差異有統計學意義。
2 結果
2.1 傷寒沙門菌pBSSB1-17基因缺陷變異株的制備
如圖1所示,PCR擴增得到1 165 bp的F1片段,以F1片段為模板,PCR擴增獲得含同源臂的1 243 bp的片段F2;將F2片段電擊轉化入含有pKD46的傷寒沙門菌野生株感受態細胞中,在含卡那霉素(終濃度為50 μg/mL)的LB平板篩選到了陽性克隆,用V-L/V-R作引物通過PCR驗證8個陽性克隆,均獲得了272 bp的DNA片段,與預期結果一致,證明p17基因缺陷株制備成功,命名為Δp17。
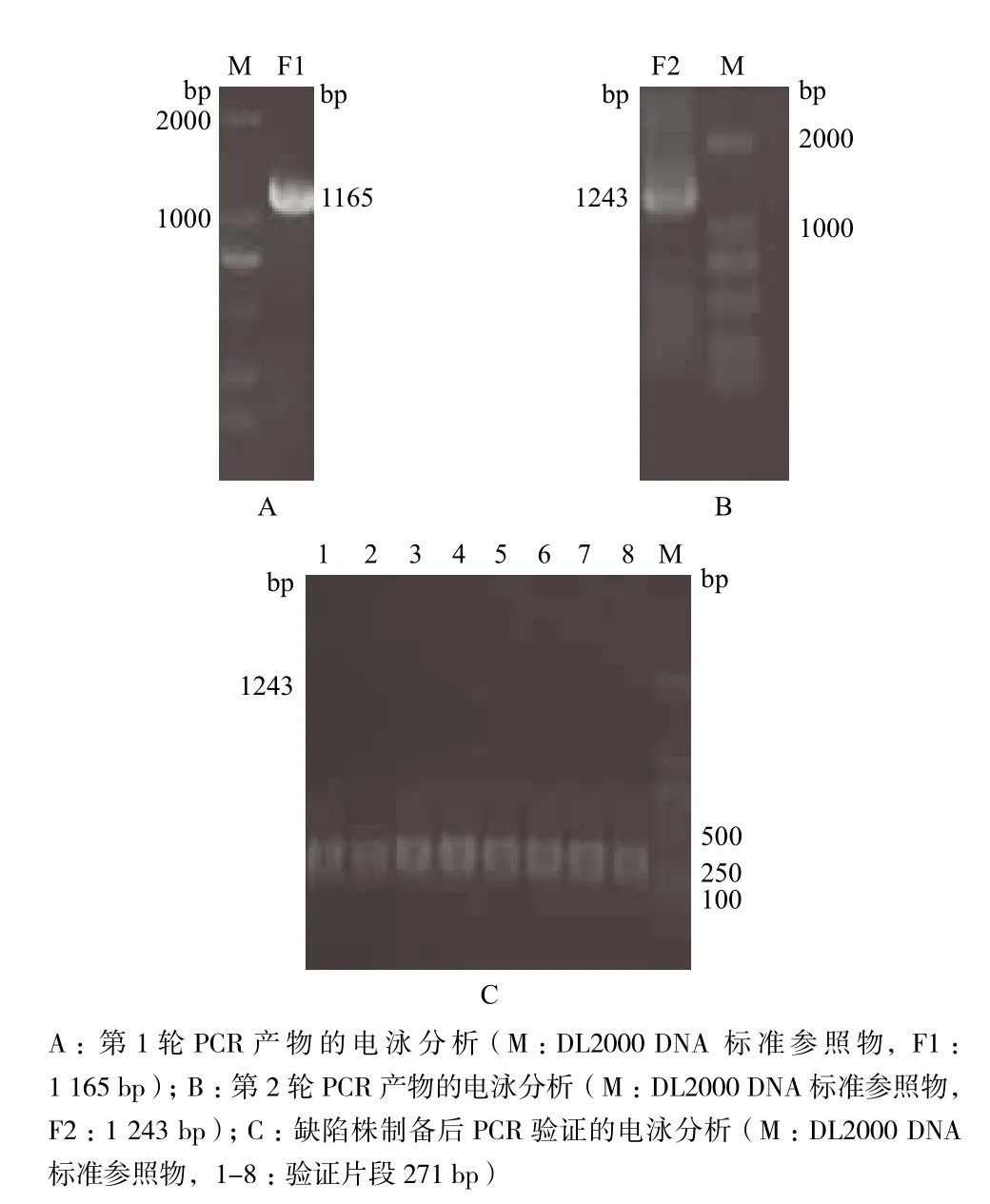
圖1 傷寒沙門菌pBSSB1的p17基因缺陷變異株的制備
2.2 p17基因缺陷株生長特性分析
p17基因缺陷株和傷寒沙門菌GIFU10007野生株在普通培養條件下的生長曲線如圖2所示,結果表明Δp17的生長能力明顯遲緩于野生株(P<0.05),提示p17基因可能參與調控細菌的生長。
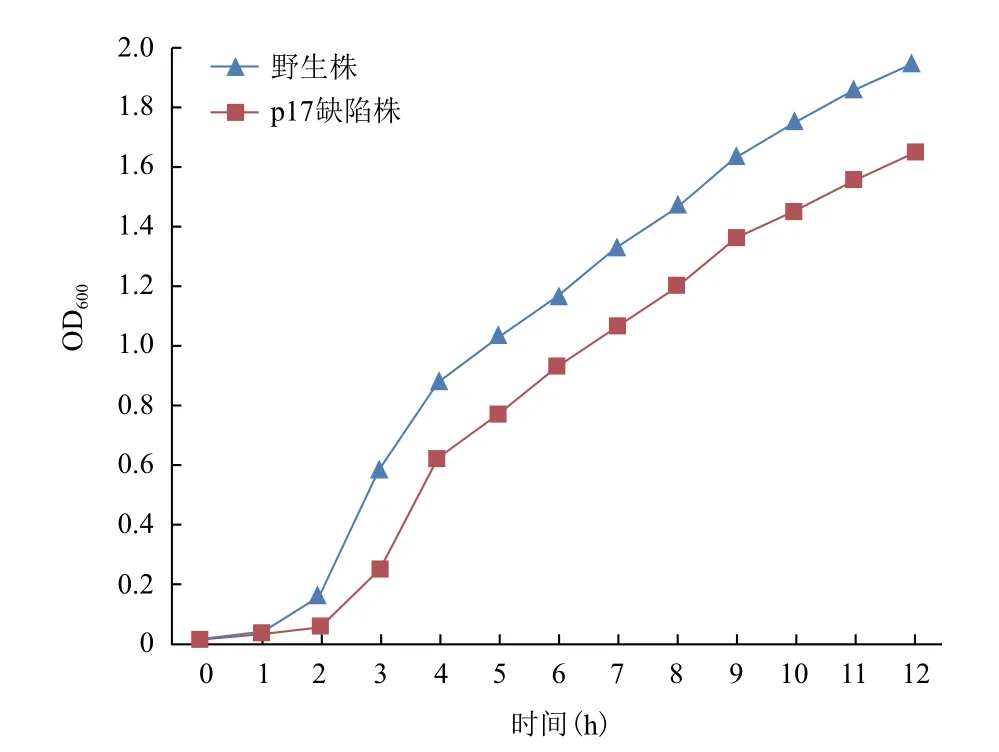
圖2 傷寒沙門菌GIFU10007野生株和p17基因缺陷株的生長曲線
2.3 野生株和缺陷株的動力比較
傷寒沙門菌GIFU10007野生株和p17基因缺陷株在0.3%的半固體培養基上的動力比較如圖3所示,p17基因缺失后其動力明顯減弱,說明p17基因參與調控細菌的動力。

圖3 傷寒沙門菌GIFU10007野生株和p17基因缺陷株的動力圖
2.4 傷寒沙門菌P17蛋白表達載體的構建
如圖4所示,PCR擴增獲得了與預期大小相符的646 bp的p17基因片段。該片段經NcoⅠ和XhoⅠ雙酶切后克隆到表達載體pET28a(+)上,將重組質粒pET28a(+)-p17轉化至大腸桿菌DH5α得到了陽性克隆 。提取陽性克隆的質粒,經Nco I和Xho Ⅰ雙酶切獲得約646 bp的片段,大小與PCR 產物一致,證明P17蛋白表達載體構建成功。

圖4 重組表達載體ET28a(+)-p17的鑒定
2.5 P17-His6重組蛋白的表達與純化
在最佳的條件即20℃,IPTG終濃度為0.1 mmol/L進行誘導5 h,上清經Ni柱純化獲得P17-His6蛋白,結果如圖5所示。
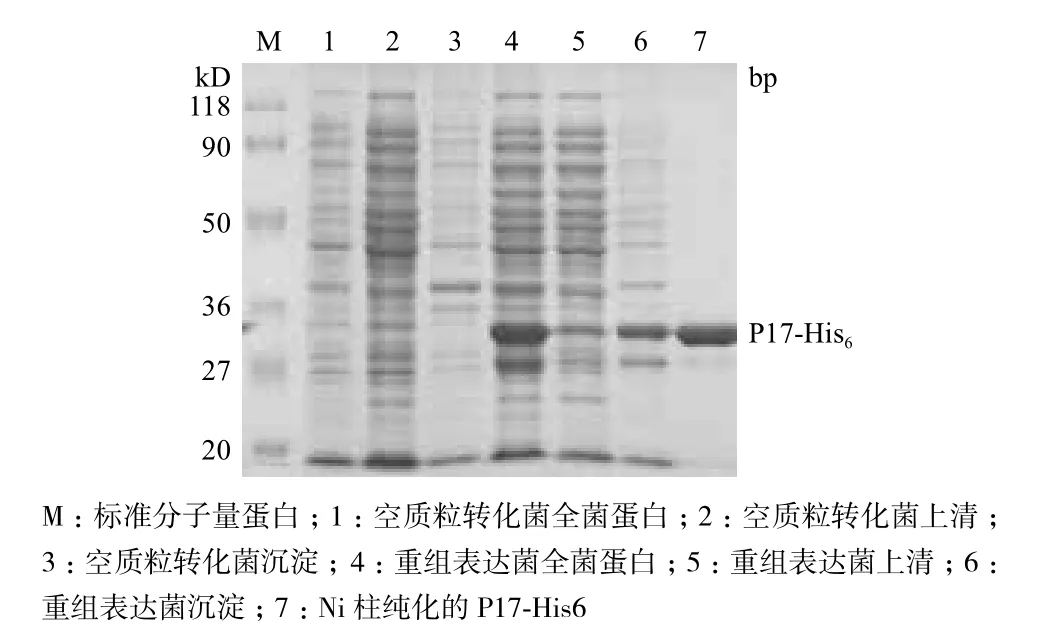
圖5 重組蛋白P17-His6的表達與純化
2.6 兔抗P17-His6多克隆抗體的鑒定及應用
將重組蛋白P17-His6免疫兔子制備了兔抗P17的多克隆抗血清,純化后的多克隆抗體與傷寒沙門菌GIFU10007野生株和p17基因缺陷株的全菌蛋白經Western blotting鑒定,如圖6所示,在缺陷株中未鑒定到p17基因的編碼蛋白P17,說明所制備的抗體與抗原反應的特異性良好,同時也說明p17基因在正常生長條件下有一定的表達,提示該基因具有一定的生物學功能。
3 討論
質粒是指細菌、酵母菌和放線菌等生物中染色體以外的DNA分子,它們能為宿主提供抗藥性等一些額外的功能[13]。絕大多數的細菌質粒都是閉合環狀DNA分子,1979年在婁徹霉菌中發現了第一例原核生物線性質粒,該發現糾正了所有細菌質粒都是環狀結構這一傳統觀點[14]。

圖6 抗體特異性的免疫印跡鑒定
線性質粒pBSSB1介導H∶z66陽性傷寒沙門菌的鞭毛單向相變換,該質粒全長27 kb,共編碼33個基因,其表達特性已由本實驗室報道,特別是其中二相鞭毛素編碼基因fljBz66和fljA樣基因功能已明確[3-8,15,16]。另外,我們發現 Fis蛋白可以影響pBSSB1的穩定性[17]。通過對其序列比對分析和GC含量偏移分析發現,緊鄰基因p17的上游可能是該質粒的一個雙向復制起始位點。基因p17編碼蛋白(P17),具有ParA和Soj蛋白的高度同源結構域序列。研究表明,ParA和Soj蛋白參與許多質粒、噬菌體和細菌基因組染色體DNA的復制和分配[9]。另外,P17具有與RepABC質粒編碼的RepA蛋白同源的ATP酶結構域,在17號基因的編碼區上游含有GANTC序列,該序列是RepABC質粒編碼的repABC操縱子所具有的特征性序列[10]。另外,Zhang等[18]在研究鏈霉菌屬線性質粒pFRL1時發現編碼的rlr基因可以幫助啟動該質粒的復制,由于rlr基因也緊鄰線性質粒pFRL1的復制起始區,我們推測線性質粒pBSSB1編碼的17號基因可能和鏈霉菌線性質粒pFRL1的rlr基因具有類似的功能。但是,P17蛋白在傷寒沙門菌中發揮作用的分子機制還有待于深入研究。
4 結論
本研究首次制備了傷寒沙門菌pBSSB1的p17基因缺陷株,發現缺陷株的生長明顯遲緩于野生株,且p17基因缺失后細菌的動力明顯減弱,這提示基因p17可能具有調控細菌的生長和動力等功能;另外,克隆表達了基因p17,純化獲得了重組蛋白P17-His6及其多克隆抗體,為進一步深入研究基因p17的生物學功能及其作用機制奠定了堅實的基礎。
[1]Everest P, Wain J, Roberts M, et al. The molecular mechanisms of severe typhoid fever[J]. Trends Microbiol, 2001, 9(7):316-
320.
[2]張海方. 沙門菌黏附和侵襲腸黏膜細胞的分子基礎[J]. 生命的化學, 2011, 31(1):124-127.
[3]Baker S, Hardy J, Sanderson KE, et al. A novel linear plasmid mediates flagellar variation in Salmonella typhi[J]. PLoS Pathog, 2007, 3(5):e59.
[4]Baker S, Holt K, Whitehead S, et al. A linear plasmid truncation induces unidirectional flagellar phase change in H:z66 positive Salmonella typhi[J]. Mol Microbiol, 2007, 66(5):1207-1218.
[5]張海方, 黃新祥, 張曉磊, 等. 抗H:z66抗體誘導z66+傷寒沙門菌鞭毛相變換特點的探討[J]. 蘇州大學學報:醫學版, 2010, 30(6):1171-1175.
[6]張海方. 沙門菌鞭毛基因表達的相變換研究進展[J]. 安徽農業科學, 2010, 38(34):19254-19255, 19265.
[7]Zou X, Huang X, Xu S, et al. Identification of fljA located on a linear plasmid as a repressor gene of fliC in Salmonella enterica serovar Typhi[J]. Microbiol Immunol, 2009, 53(4):191-197.
[8]Xu S, Zou X, Sheng X et al. Expression of fljB:z66 on a novel linear plasmid of Salmonella enterica serovar Typhi is dependent on FliA and FlhDC and regulated by OmpR[J]. Braz J Microbiol, 2010, 41(3):729-740.
[9]Hester CM, Lutkenhaus J. Soj(ParA)DNA binding is mediated by conserved arginines and is essential for plasmid segregation[J]. Proc Natl Acad Sci USA, 1999, 96(1):73-78.
[10]Pinto UM, Pappas KM, Winans SC. The ABCs of plasmid replication and segregation[J]. Nat Rev Microbiol, 2012, 10(11):755-765.
[11]詹莉芳, 張曉磊, 翁曉琴, 等.OxyR蛋白不同應激條件下傷寒沙門菌生存能力的影響[J]. 江蘇大學學報:醫學版. 2013, 23(2):93-98.
[12]張海方, 高宇琳, 黃新祥, 等. 傷寒沙門菌鞭毛素基因fljB:z66的克隆表達及其多克隆抗體的制備[J]. 江蘇大學學報:醫學版, 2009, 19(5):376-379.
[13]朱云霞, 趙昕, 張綺思, 張海方. 細菌線性質粒的復制研究[J]. 生物技術通報, 2014(5):32-36.
[14]Hayakawa T, Otake N, Yonehara H, et al. Isolation and characterization of plasmids from Streptomyces[J]. J Antibiot, 1979, 32(12):1348-1350.
[15]Xu S, Zhang H, Sheng X, et al. Transcriptional expression of fljB:z66, a flagellin gene located on a novel linear plasmid of Salmonella enterica serovar Typhi under environmental stresses[J]. New Microbiol, 2008, 31(2):241-247.
[16]Zhao X, Zhu Y, Zhang H, et al. Transcriptional expression of 6 genes located on pBSSB1 of Salmonella enterica serovar Typhi in different growth phases and environmental stresses[J]. Curr Microbiol, 2014, 69(3):252-257.
[17]Zhang H, Ni B, Zhao X, et al. Fis is essential for the stability of linear plasmid pBSSB1 and affects the motility of Salmonella enterica serovar Typhi[J]. PLoS One, 2012, 7:e37462.
[18]Zhang R, Peng S, Qin Z. Two internal origins of replication in Streptomyces linear plasmid pFRL1[J]. Appl Environ Microbiol, 2010, 76(17):5676-5683.
(責任編輯 李楠)
A Preliminary Study on the Biological Function of a Gene p17 Located on a Linear Plasmid pBSSB1 of Salmonella enterica serovar Typhi
Zhu Yunxia1Wu Jia1Gong Mingyu1Hou Shuning1Zhang Qisi1Chen Minjie1Zhang Haifang1,2
(1. Department of Biochemistry and Molecular Biology,School of Medical Science and Laboratory Medicine,Jiangsu University,Zhenjiang 212013;2. Department of Clinical Laboratory,the Second Affiliated Hospital of Soochow University,Suzhou 215004)
pBSSB1 is a linear plasmid which mediates the flagellar phase variation in H:z66 positive Salmonella enterica serovar Typhi(S. Typhi). The gene named p17 is located on pBSSB1 and it encodes the protein P17 whose function is unknown. The p17 deleted mutant of S. Typhi was prepared through the λ-Red recombination system. The growth curves of wild type and p17 mutant strain were detected to compare their growth ability. The motility of wild type and p17 mutant strains was compared through the experiments on the semi-solid LB plates. The specific primers were designed to amplify the gene p17 by PCR. The amplicon was inserted into the expression vector pET28a(+)to construct recombinant vector pET28a(+)-p17, which was then transferred to E. coli BL21(DE3)to be expressed. The recombinant protein P17-His6was purified with Ni-TED packed column and was used as immunogen to prepare the rabbit anti-P17 polyclonal antibody. The results showed that p17 deleted mutant of S. Typhi was constructed successfully. It was showed that the growth of p17 mutant strain was significantly slower compared with the wild type strain(P<0.05). The motility of p17 mutant strain was decreased obviously compared to the wild type strain. The gene p17 of S. Typhi was inserted into the vector pET-28a(+)and was expressed in E. coli BL21(DE3). The rabbit anti-P17 polyclonal antibody was prepared.
Salmonella enterica serovar Typhi Linear plasmid pBSSB1 p17
2014-02-25
國家自然科學基金項目(31000076),中國博士后科學基金項目(2013M531278),江蘇省博士后科研資助計劃項目(1202011B),江蘇大學大學生科研立項資助項目(12A123,13A239)
朱云霞,女,碩士研究生,研究方向:病原菌分子致病機制;E-mail:zhuyunxia3032@163.com
張海方,男,博士,副教授,研究方向:分子細菌學;E-mail:haifangzhang@sina.com
