產D-氨基酰化酶菌株的篩選及活性菌株A55的初步研究
2014-03-21 06:26:44倪孟祥席煥鴿
化學與生物工程 2014年5期
關鍵詞:生長
倪孟祥,席煥鴿
(中國藥科大學生命科學與技術學院,江蘇南京210009)
人們曾經認為D-氨基酸在生物進程中發揮著相對較少的作用,并稱之為“非天然氨基酸”。然而近期的研究表明,D-氨基酸在細胞結構及信號調控中發揮著重要的作用,科學家們認為闡明D-氨基酸如何調控細胞壁重塑及生物被膜降解的機制不僅能夠對細胞質外的進程有一個新的理解,并且能夠促進新療法的發展[1]。隨著研究的不斷深入,D-氨基酸在醫藥、農業和食品等領域的用途被迅速發掘,如D-氨基酸及其衍生物作為重要的手性原材料被廣泛地應用于半合成抗生素、多肽激素、殺蟲劑和甜味劑等的合成中[2-5]。
目前,隨著應用的推廣,D-氨基酸的工業需求量也逐漸增大。D-氨基酸制備方法主要有:對映體拆分法、化學合成法和生物法。酶法合成作為生物法中應用最多的方法,具有無污染、反應條件溫和、操作簡便、成本低、產物純度高等優點,展現出很好的商業前景[6]。在酶法合成D-氨基酸的過程中,D-氨基酰化酶是主要使用的幾種酶之一。迄今已發現的產D-氨基酰化酶的微生物包括產堿菌屬、假單胞菌屬、貪噬菌屬、狹長平胞屬、擬無枝酸菌屬、博德特菌屬、葡萄糖菌屬、甲基桿菌屬、類諾卡氏菌屬、微桿菌屬、鏈霉菌屬和木霉屬等[7]。其中,來自產堿菌屬的D-氨基酰化酶已經應用于工業化生產[8]。然而,國內對D-氨基酰化酶的研究還處于基礎階段,商品酶仍需進口。
土壤篩選是獲得產酶微生物的重要途徑。作者從氨基酸類化工廠附近土樣中篩選產D-氨基酰化酶的菌株,并對活性較高的菌株進行了鑒定及培養條件的初步優化。
1 實驗
1.1 材料與試劑
土樣采自江蘇省某氨基酸類化工廠排污區附近。
細菌基因組DNA提取試劑盒、質粒小量提取試劑盒、瓊脂糖凝膠回收試劑盒,上海生工生物工程有限公司;250bp DNA Ladder Marker,寶生物工程(大連)有限公司;Taq DNA聚合酶,北京康為世紀有限公司;DNA引物(PAEG純),上海捷瑞公司;其它化學試劑均為分析純或色譜純。
1.2 培養基
分離培養基:N-乙酰-D-甲硫氨酸1%,K2HPO4·3H2O 0.1%,MgSO4·7H2O 0.05%,瓊脂2%,蒸餾水配制,調節pH值至7.0。
發酵培養基:N-乙酰-D-甲硫氨酸1%,K2HPO4·3H2O 0.1%,MgSO4·7H2O 0.05%,酵母浸膏2%,蒸餾水配制,調節pH值至7.0。
1.3 方法
1.3.1 菌株的分離
采用平板稀釋法從土樣中分離菌株。取土壤樣品5g,用無菌生理鹽水逐級稀釋至10-1、10-2、10-3、10-4、10-5數量級,各取0.2mL均勻涂布于分離平板上,每個稀釋度涂3個平板,30℃倒置培養48h。觀察并挑選生長快速、菌落較大的菌株劃線純化,純化的菌株斜面置于4℃冰箱保存[9]。
1.3.2 菌株的篩選
1.3.2.1 初篩
將純化的菌株轉接到發酵培養基中,于30℃、200 r·min-1搖瓶培養48h。低溫離心收集菌體,菌體用適當比例的50mmol·L-1Tris-HCl緩沖溶液(pH值7.8)充分懸浮,加入適量200mmol·L-1的底物N-乙酰-D-甲硫氨酸至濃度為20mmol·L-1,混合均勻,于37℃恒溫水浴振蕩1h,之后立即沸水浴10min終止反應。離心后取上清液1μL點樣進行蛋氨酸的硅膠薄層層析,展開劑為V(正丁醇)∶V(冰乙酸)∶V(水)=4∶1∶1。每次需展開劑25mL,另加入1mL 0.5%茚三酮溶液,混合均勻,展開結束后,電吹風熱風吹干;顯色斑點與0.05mmol·L-1蛋氨酸的標樣斑點比較,檢測蛋氨酸的含量以反映D-氨基酰化酶的活力。為了消除所用試劑和菌體本身所含氨基酸對測定的干擾,將懸浮后的菌體先煮沸10min,然后加入底物,其余操作同上,作為空白對照。保留蛋氨酸斑點較大且無雜斑點的菌株進行復篩[10]。
1.3.2.2 復篩
將初篩保留的菌株轉接到發酵培養基中,于30℃、200r·min-1搖瓶培養48h;采用茚三酮比色法測定酶活[10],方法同1.3.2.1。
酶活力單位(U)定義:在pH值7.8、37℃條件下,1min內酶催化底物水解釋放出1μmol D-甲硫氨酸為1U。
1.3.3 菌株A55的鑒定
1.3.3.1 形態學特征觀察及生理生化特征測定
依據《伯杰氏細菌鑒定手冊》(第八版)[11]和《常見細菌系統鑒定手冊》[12]進行試驗,鑒定菌種。
1.3.3.2 菌株A55的16SrDNA序列測定及系統發育分析
利用細菌基因組DNA提取試劑盒提取菌株A55的基因組DNA。以基因組DNA為模板,用細菌16S rDNA基因通用引物進行PCR擴增。PCR擴增參數:94℃預變性4min;94℃解鏈40s,55℃退火40s,72℃延長1.5min,30個循環;最后72℃延長10 min。PCR產物用1%瓊脂糖凝膠檢測,純化后交南京思普金公司測序。將測序得到的16SrDNA序列通過Blast與GenBank中細菌及古菌的16SrDNA序列進行比對,利用Mega5.2軟件進行系統發育分析。采用鄰接法(neighbor-joining method)構建系統發育樹,并對所構建的系統發育樹進行自舉分析(bootstrap method,1 000次重復),估算其內分支的支持率[13]。
1.3.4 影響D-氨基酰化酶合成的因素考察
以發酵培養基為基礎培養基,考察培養溫度、培養基初始pH值、搖床轉速、搖瓶裝樣量、菌齡、碳源、氮源等對菌體生長及酶活力的影響,優化菌株A55的培養條件[14]。
2 結果與討論
2.1 菌株的分離及活性篩選
本實驗以N-乙酰基甲硫氨酸作為分離培養基中的唯一碳氮源。從來自氨基酸類化工廠附近的8份土樣中,分離了約500株菌,并對其進行了D-氨基酰化酶活力的初篩與復篩,得到了酶活力及穩定性均較好的菌株A55,酶活力達到70U,并對其進行后續研究。
2.2 菌株A55的鑒定結果
2.2.1 形態學特征及生理生化特征(表1)

表1 菌株A55的生理生化特征Tab.1 Biochemical and physiological characteristics of strain A55
菌株A55在LB固體培養基上生長迅速,菌落圓形呈淡黃色,表面濕潤,中心凸起,邊緣整齊;在分離固體培養基上生長相對緩慢,菌落點狀呈乳白色,表面濕潤,邊緣整齊。革蘭氏染色陽性,桿狀,鏈狀排列。
2.2.2 16SrDNA序列分析
PCR擴增得到的菌株A55的16SrDNA序列全長為1 416bp,將該序列與GenBank數據庫中細菌及古菌的16SrDNA序列進行Blast分析。結果表明,菌株A55與耐寒短桿菌相似度最高,達到100%,通過Mega5.2軟件得到了以16SrDNA序列為基礎的系統發育樹(圖1)。
由圖1可知,菌株A55與耐寒短桿菌Brevibacterium frigoritolerans處于同一分支,親緣關系最近,結合形態學及生理生化特征可以確定菌株A55為耐寒短桿菌。
2.3 菌株A55發酵條件的優化
2.3.1 培養條件的確定
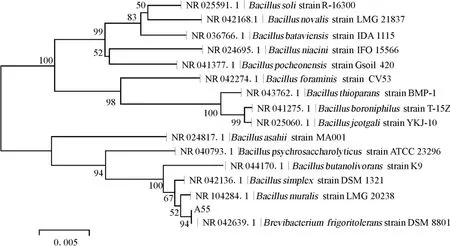
圖1 菌株A55的系統發育樹Fig.1 Phylogenetic tree of strain A55
溫度及培養基pH值對微生物的生長有著非常重要的影響,搖床轉速及搖瓶裝樣量主要影響微生物的供氧,也是發酵中不可忽視的因素。考察了培養溫度、培養基初始pH值、搖床轉速、裝樣量對菌株A55生長及酶活力的影響,結果見圖2。

圖2 培養溫度(a)、培養基初始pH值(b)、搖床轉速(c)及裝樣量(d)對菌株A55生長及酶活力的影響Fig.2 Effects of culture temperature(a),culture initial pH value(b),shaker speed(c)and loaded liquid(d)on biomass and enzyme activity of strain A55
由圖2可知:菌株A55的最佳培養溫度為30℃,在此溫度下,菌體的生長情況及酶活力均較好;菌株A55在初始pH值為5的培養基上無法生長,其最適初始pH值為7;菌株A55發酵的最佳搖瓶轉速為200 r·min-1,最佳裝樣量為15%,這與菌株A55嚴格好氧密切相關。因此,確定菌株A55適宜的培養條件為:培養溫度30℃,培養基初始pH值7,搖瓶轉速200r·min-1,搖瓶裝樣量15%。在此條件下繼續考察其它因素的影響。
2.3.2 菌齡對菌株生長及酶活力的影響
菌株A55所產的D-氨基酰化酶屬于誘導酶,只有在培養基中添加N-乙酰基氨基酸作為誘導物時,菌株才能合成D-氨基酰化酶。菌株A55的生長曲線及菌齡對酶活力的影響見圖3。
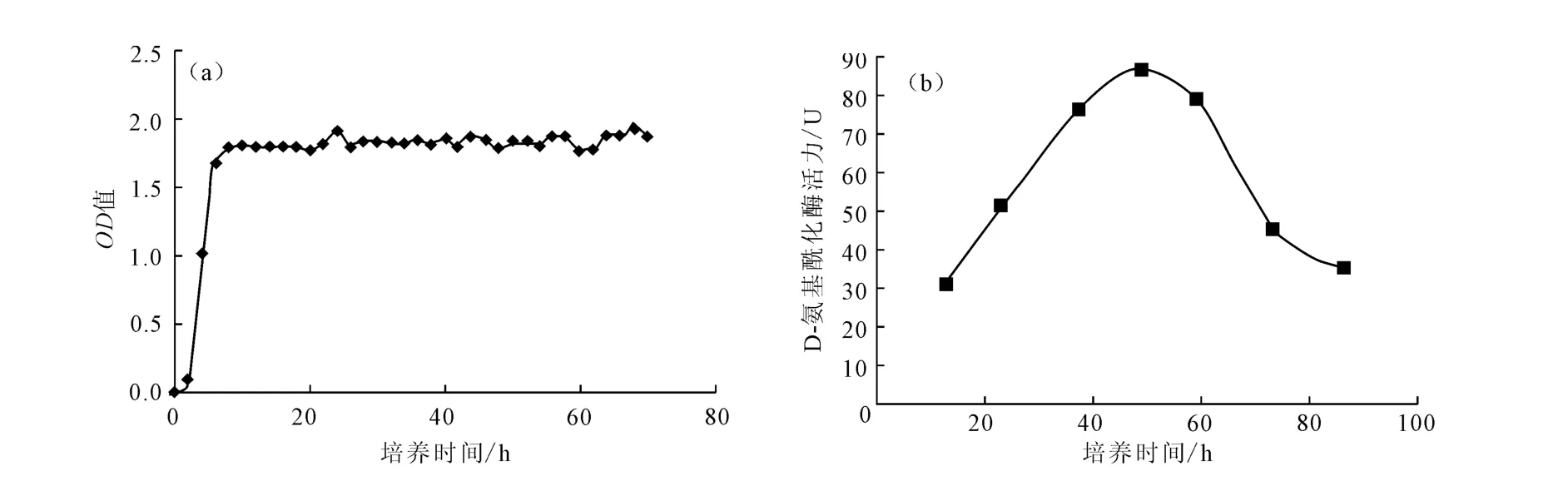
圖3 菌株A55的生長曲線(a)及菌齡對酶活力的影響(b)Fig.3 Growth curve of strain A55(a)and effect of strain age on enzyme activity(b)
由圖3a可知,由于種子培養基與發酵培養基相同,菌株A55調整期非常短暫,很快進入對數期,在第8h即進入穩定期。由圖3b可知,菌株A55在36~60h之間D-氨基酰化酶活力較高,第72h活力明顯降低,可以判斷菌株A55從第60h開始進入衰亡期。因此,確定菌株的最佳培養時間為48h,酶活力達到82.99U。
2.3.3 碳、氮源對菌株生長及酶活力的影響
分別添加1%的葡萄糖、蔗糖、淀粉、甘油,0.5%的硝酸鈉、硫酸銨、尿素、蛋白胨,以發酵培養基為對照,考察不同碳、氮源對菌株A55的生長及酶活力的影響,結果見圖4。

圖4 碳源(a)和氮源(b)對菌株A55生長及酶活力的影響Fig.4 Effects of carbon sources(a)and nitrogen sources(b)on biomass and enzyme activity of strain A55
由圖4a可知,在培養基中分別添加不同碳源后,菌株A55的菌體量均明顯增加,葡萄糖的影響尤其突出,菌體量增加至對照的2.08倍;然而D-氨基酰化酶活力均明顯降低。由圖4b可知,硝酸鈉、硫酸銨及尿素等無機氮源對菌株A55的菌體量影響不大,D-氨基酰化酶活力有所降低;相比之下,蛋白胨的添加使菌株A55的菌體量增加至對照的2.78倍,同時蛋白胨對D-氨基酰化酶活力影響也較大,使其降低至對照的24.67%。表明,培養基營養豐富反而抑制D-氨基酰化酶的合成,這與該酶屬于誘導酶密切相關。
3 結論
從氨基酸類化工廠附近土樣中分離出產D-氨基酰化酶的菌株A55,通過形態學觀察、生理生化特征測定及16SrDNA系統發育分析,鑒定其為短桿菌屬的耐寒短桿菌,具有D-氨基酰化酶活力的耐寒短桿菌此前未見報道。另外,對該菌株的發酵條件進行了初步優化,其最佳產酶發酵條件為:培養溫度30℃,培養基初始pH值7,搖床轉速200r·min-1,搖瓶裝樣量15%,培養時間48h。為實現國內D-氨基酰化酶的商品化及酶法合成D-氨基酸的工業化奠定了基礎。
[1] CAVA F,LAM H,de PEDRO M A,et al.Emerging knowledge of regulatory roles of D-amino acids in bacteria[J].Cell Mol Life Sci,2011,68(5):817-831.
[2] SHINADA T,ISHIDA T,HAYASHI K,et al.Synthesis of leucine-enkephalin analogs containingα-amino squaric acid[J].Tetrahedron Letters,2007,48(43):7614-7617.
[3] NAGATA Y,HIGASHI M,ISHII Y,et al.The presence of high concentrations of free D-amino acids in human saliva[J].Life Sciences,2006,78(15):1677-1681.
[4] CIACCIO C,TUNDO G R,GRASSO G,et al.Somatostatin:A novel substrate and a modulator of insulin-degrading enzyme activity[J].Journal of Molecular Biology,2009,385(5):1556-1567.
[5] MIYOSHI Y,KOGA R,OYAMA T,et al.HPLC Analysis of naturally occurring free D-amino acids in mammals[J].Journal of Pharmaceutical and Biomedical Analysis,2012,69:42-49.
[6] 于平.生物轉化和手性拆分技術制備D-氨基酸研究進展[J].生物學通報,2005,40(9):3-5.
[7] LIU J,ASANO Y,IKOMA K,et al.Purification,characterization,and primary structure of a novel N-acyl-D-amino acid amidohydrolase from Microbacterium natoriense TNJL143-2[J].Journal of Bioscience and Bioengineering,2012,114(4):391-397.
[8] WAKAYAMA M,YOSHIMUNE K,HIROSE Y,et al.Production of D-amino acids by N-acyl-D-amino acid amidohydrolase and its structure and function[J].Journal of Molecular Catalysis B:Enzymatic,2003,23(2-6):71-85.
[9] 倪孟祥,胡穎.北極海泥來源抗菌活性真菌的篩選及菌株H5的初步研究[J].化學與生物工程,2013,30(5):61-64.
[10] 劉偉.產D-氨基酰化酶菌株的選育[D].杭州:浙江大學,2007.
[11] R E布坎南,N E吉本斯.伯杰氏細菌鑒定手冊[M].第八版,北京:科學出版社,1984.
[12] 東秀珠,蔡妙英.常見細菌系統鑒定手冊[M].北京:科學出版社,2001.
[13] 趙章鎖,郝曉潔,魏聰,等.苯酚降解菌SW34的鑒定[J].安徽農業科學,2012,40(31):15372-15374.
[14] 滿永博,范季瀛,蘇剛,等.培養條件及培養基組分對粘質沙雷氏菌生長及產D-乳酸的影響[J].化學與生物工程,2013,30(7):56-60.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
