利用化學小分子實現人胚胎干細胞多能性狀態的轉化*
2014-03-18 09:22:42歐陽琦楊青青周曉櫻盧光琇胡維新
激光生物學報 2014年4期
關鍵詞:體系
歐陽琦,楊青青,周曉櫻,林 戈,盧光琇,胡維新
(中南大學 a.生命科學學院;b.生殖與干細胞研究所,湖南 長沙410078)
小鼠胚胎干細胞依據其來源的發育階段以及細胞的生物學特性可分為兩類:一類是傳統意義上的胚胎干細胞(mouse embryonic stem cells,mESCs),來源于植入前囊胚的內細胞團,處于原始多能性狀態(naive)[1];一類是上胚層干細胞(mouse epiblast stem cells,mEpiSCs),來源于植入后胚胎的上胚層,處于始發態的多能性狀態(Primed)[2]。這兩類小鼠多能性干細胞從克隆形態、分化能力、維持自我更新的信號通路、表觀遺傳學特征以及單細胞生長能力等方面都存在明顯不同。mESCs克隆形態緊密,呈鳥巢狀隆起,三維生長;依賴LIF/stat3信號通路的活化來維持不分化狀態;雌性mESCs具有兩條活化的X染色體;能夠耐受酶消化進行單細胞傳代,有利于基因定點改造和篩選。而mEpiSCs培養時克隆形態扁平,二維生長;其不分化狀態的維持依賴bFGF和Activin A/Nodal信號通路;雌性mEpiSCs通常失活一條X染色體;單細胞傳代效率低,難以進行基因定點改造操作。此外,mESCs具有更為完整的多能性,將mESCs注射到宿主囊胚腔內能夠整合并參與后者的發育過程,形成嵌合體小鼠,并能分化為生殖細胞而實現生殖系的傳遞。而mEpiSCs只具備部分多能性,雖然能夠向三個胚層的細胞分化,但不具有形成嵌合胚胎的能力,也基本無法實現生殖系的傳遞。以往常規“bFGF+Activin A”培養體系下分離的人胚胎干細胞(human ESCs,hESCs)在生物學特性上更接近于mEpiSCs,即處于始發態多能性[3,4]。因此,單細胞傳代效率低以及嵌合能力低等特性將在一定程度上限制了hESCs在基礎和臨床研究領域更廣泛的應用。
從2007年開始,人們嘗試各種方法建立naive的hESCs。Hanna等[5]過表達多能性基因將現有的hESCs逆轉至原始多能性狀態,但是細胞不穩定,撤除外源性基因的表達,細胞將失去原始態的表型,回復到始發態。Beucker課題組[6]利用在LIF條件下過表達5個重編程基因獲得了原始態的人誘導性多能干細胞,但是這類干細胞不能啟動內源性多能性關鍵基因的表達,分化能力受損。轉入外源性基因的方法只能獲得亞穩態的原始多能性干細胞,且具有安全性隱患,因此建立一種無需轉基因操作、僅通過改變培養條件獲得原始態hESCs的方法是亟待解決的重要問題。目前國外有研究組利用不同的化學小分子和生長因子組合實現了hESCs多能性的轉化,這些化學小分子組合通過激活或者抑制發育過程中不同的關鍵信號通路,使逆轉后的hESCs具有部分原始態多能性特征,但是逆轉后的hESCs之間存在明顯的基因表達差異[7-10]。而建立原始態hESCs的研究在國內尚未見報道。
本實驗利用已經建立的hESCs資源,綜合并改良文獻報道的原始多能性干細胞培養條件,利用化學小分子對已建系的hESCs進行多能性轉化,并評估了轉化后hESCs的原始多能性特征。
1 材料與方法
1.1 試劑
1.1.1 人胚胎干細胞系 chHES-278為本實驗室自主建系,核型為46,XX;已進行相應干細胞特征性鑒定,符合NIH對胚胎干細胞的鑒定標準。
1.1.2 小鼠胚胎成纖維細胞(MEF)從CF1小鼠第12.5天胚胎中分離獲得,經絲裂霉素有絲分裂滅活后,作為hESCs的飼養層細胞。
1.1.3 培養基及配制 始發態hESCs培養基(DFES):基礎培養基DMEM/F12,15%血清替代物,0.1 mmol/Lβ-ME,2 mmol/L L-Glu,4 ng/mL bFGF,1%非必需氨基酸。
擬胚體(EB)培養基:即始發態hESCs培養基不添加bFGF。
2i(ERK1/2抑制劑及GSK3β抑制劑)培養基:基礎培養基DMEM/F12和Neurobasal,1%N2,2%B27,0.1 mmol/Lβ-ME,2 mmol/L L-Glu,3μmol/L CHIR99021(GSK3βi,Stemgent公 司),1μmol/L PD0325901(ERK1/2i,Tocris公司),1 000 U/L LIF(Millipore公司)。
原始態hESCs培養基(NHSM):基礎培養基DMEM/F12,2 g/L AlbuMAX,1%N2,1%非必需氨基酸,0.1 mmol/Lβ-ME,1 mmol/L L-Glu,12.5 mg/mL insulin(sigma公司),8 ng/mL bFGF,1 ng/mL TGF-β1(Peprotech公 司),3μmol/L CHIR99021(GSK3βi,Stemgent公司),0.5μmol/L PD0325901(ERK1/2i,Tocris公司),10μmol/L SP600125(JNKi,Tocris公司),5μmol/L SB202190(p38i,Axon Medchem公司),5μmol/L Go6983(PKCi,Tocris公司),5μmol/L Y-27632(ROCKi,Millipore公 司),1 000 U/L LIF(Millipore公司)。
1.2 胚胎干細胞培養
以小鼠胚胎成纖維細胞為飼養層細胞,分別采用hESCs培養基、2i培養基和NHSM培養基。機械或者酶消化傳代,置于37℃,5%CO2培養箱中,每5-7天傳代一次。
1.3 hESCs的相關檢測
1.3.1 堿性磷酸酶(AKP)的檢測 采用Zymed Laboratories Inc公司提供的試劑盒對未分化的hESCs克隆進行檢測,操作步驟按試劑盒說明進行,陽性染色結果為紫色。周邊的MEF不著色,為陰性對照。
1.3.2 細胞特異抗原的檢測 多能性干細胞特異性抗原,包括:TRA-1-60、TRA-1-81、OCT-4;三胚層特異性標記,包括:AFP(內胚層)、β-tubulin(外胚層)、SMA(中胚層);其他抗體:H3K27me3、E-cadherin。均采用間接免疫熒光法來檢測,步驟如下:4%的多聚甲醛固定20 min;0.1%Triton-X-100透膜10 min(針對核內抗原,胞膜抗原不需要透膜);正常驢血清室溫封閉30 min;一抗4℃孵育過夜;加入Alexa Flour 488-連接二抗,室溫避光孵育1 h;DAPI復染核;熒光顯微鏡下觀察檢測結果并照相。
1.3.3 多能性相關基因的檢測 收集未分化的hESCs,用Trizol抽提總RNA,用Roche公司提供的逆轉錄試劑盒將1μg RNA反轉錄為cDNA,再以cDNA為模板進行Real-time PCR分析,以檢測其中多能性相關基因的表達,28S為反應體系陽性對照。Realtime PCR反應條件:95℃5 min;45個循環(95℃10 s,58℃10 s,72℃15 s)。
1.3.4 單克隆形成率實驗 未分化的hESCs經胰酶消化成單細胞后計數,按照200個細胞/10 cm培養皿的密度種植,根據消化前hESCs的培養條件分別采用原始態或者始發態hESCs培養基進行培養,6天后計數克隆數目,克隆數目與接種細胞數目之比即為單克隆形成率,每組實驗重復3次。
2 實驗結果
2.1 hESCs無法在2i體系下長期培養
mESCs既可以在LIF+2i體系下成功分離建系,也可以長期穩定培養。但是將已建立的具有始發態多能性特征的hESCs轉至2i體系不能保持未分化狀態,難以傳代培養,往往3代之內即完全分化,AKP染色呈陰性(圖1)。
2.2 2i聯合JNKi、P38i可以獲得類mESCs的原始態hESCs
在2i體系中進一步添加JNK抑制劑(SP600125)、p38抑制劑(SB202190)并聯合生長因子bFGF、TGFβ,可以獲得形態上類似mESCs的hESCs,細胞形態表現為緊密、圓形、隆起的克隆。培養過程中,hESCs克隆周邊始終有少量分化細胞,將ERKi(PD0325901)的濃度從1μmol/L降低至0.5μmol/L,則細胞基本無分化。(圖2)
2.3 轉化后的hESCs具有naive多能性特征
將NHSM培養體系下培養的hESCs進行多能性特征性鑒定,結果提示轉化后的hESCs具有原始多能性的特征。始發態培養體系的hESCs進行H3K27me3染色,細胞內有凝集點,說明一條X染色體失活,轉移至naive體系下培養,H3K27me3凝集點消失,說明X染色體重新活化,再將轉化后的hESCs形成EB自發分化后,H3K27me3凝集點再次出現,說明X染色體又重新啟動失活。將兩種體系下的hESCs進行Xist基因表達的對比分析,結果也提示后者的Xist表達水平明顯偏低,與H3K27me3染色結果吻合。DNA甲基轉移酶(DNMT)主要參與基因組從頭甲基化,在哺乳動物的基因印記和X染色體失活的調節中起關鍵作用,與始發態培養體系相比,naive培養體系下hESCs的DNMT3A和DNMT3B表達水平明顯升高。此外,轉化后的hESCs單克隆形成率明顯增高。始發態hESCs單克隆形成率約3%,添加具有抗凋亡作用的ROCKi,單克隆形成率提高到19%;轉化的hESCs單克隆形成率為38%,添加ROCKi后,單克隆形成率上升到75.6%,明顯高于始發態hESCs。(圖3)

圖1 2i體系和bFGF體系下hESCs培養的形態變化及AKP染色情況Fig.1 The morphologic change and AKP staining of hESCs cultured in the culture system containing 2i or bFGF
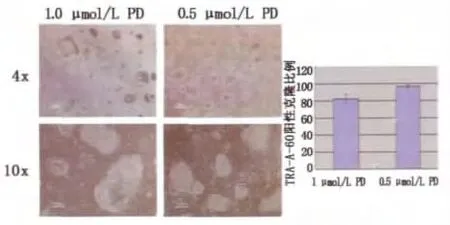
圖2 原始態hESCs培養體系中不同ERKi濃度培養下的hESCs形態圖Fig.2 The morphologic change of hESCs cultured in the naive ESCs culture system containing ERKi with different concentration.
2.4 轉化后的hESCs不具備mESCs多能性基因表達模式
將NHSM體系轉化后的hESCs與轉化前的hESCs進行多能性基因的表達對比分析,除LIMCH1和TBX3以外,mEpiSCs中明確高表達的多能性基因OTX2以及mESCs高表達的多能性相關基因KLF4、PRDM14、REX1等在兩者之間均無明顯差別,ESRRB、FGF5、FOXA2不表達(圖4)。
3 討論
小鼠上胚層干細胞(mEpiSCs)的成功分離使人們意識到胚胎干細胞由于組織來源不同而具有不同的多能性狀態,早期原始多能性階段主要指植入前囊胚內細胞團,以mESCs為代表,晚期始發態多能性階段主要指植入后胚胎的上胚層,以mEpiSCs和既往bFGF體系下培養的hESCs為代表。這兩類多能性干細胞在生物學特性上存在明顯不同,因此在應用前景上也有明顯差別。mESCs具有更廣泛的發育潛能,能夠整合到宿主胚胎參與各器官組織的發育,形成嵌合體,因此可以用于制備轉基因動物或者在宿主體內形成完全由供者細胞來源的器官,能夠生殖系傳遞,在未來器官組織工程方面具有很大的發展前景。mEpiSCs和hESCs的發育潛能被進一步限定,雖然能夠向三個胚層各種組織細胞分化,但是不能整合到宿主胚胎形成嵌合體,且不具備生殖系傳遞的能力,因此在器官組織工程方面用途受限。而轉基因往往需要進行克隆篩選,mESCs能夠耐受單細胞傳代,是目前研究基因功能最常用的的哺乳動物,而mEpiSCs和hESCs在胰酶處理后單克隆形成能力很差,很大程度上限制了hESCs的基因操作。
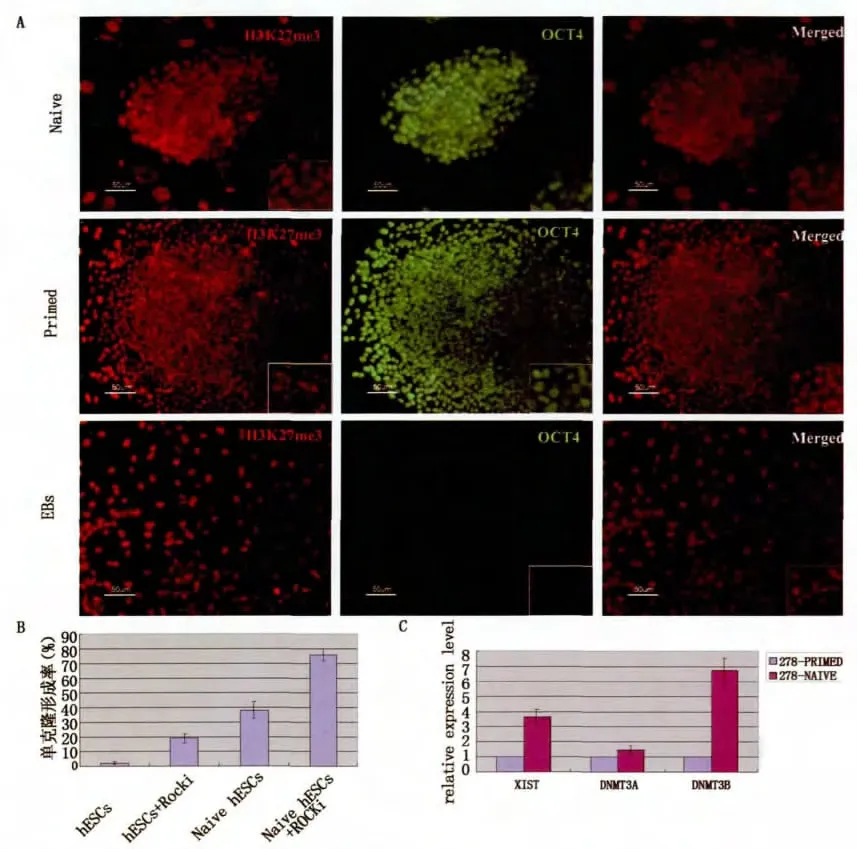
圖3 NHSM培養條件下的hESCs表達原始態多能性的生物學特性Fig.3 naive Characteristics of transformed hESCs cultured in NHSM media
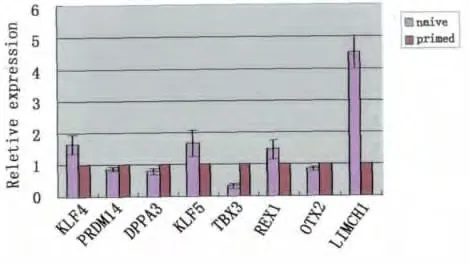
圖4 轉化后原始態與始發態hESCs多能性基因表達的對比分析Fig.4 The expression analysis of pluripotency-related gene between naive and primed hESCs
意識到胚胎源性干細胞具有兩種不同的多能性狀態,人們開始尋找建立naive狀態hESCs的方法。2008年以前,mESCs依賴LIF+BMP4體系來維持自我更新,2008年,Ying創建了2i體系,ERK1/2i能夠抑制mESCs自身分泌的FGF4,而GSK3βi能夠活化Wnt信號通路,均可以加強mESCs的自我更新能力,能夠更好地維持原始多能性狀態[11]。許多在LIF+BMP體系下難以建系的物種都能夠通過2i體系建立原始多能性干細胞[12-15]。本研究將始發態的hESCs轉入2i體系中培養,細胞很快分化,說明2i不足以抗衡人類原始多能性狀態維持所需要克服的內因障礙。Hanna等借助外源性基因(例如:Oct4、Klf2、Klf4)的導入,幫助hESCs在2i體系中獲得原始多能性,但是這種逆轉不穩定,撤除外源性基因的表達,細胞將迅速回復到始發態多能性水平[5]。Buecker等過表達5個重編程因子雖然能夠獲得原始多能性的人誘導性多能干細胞(hiPSCs),但這類細胞以犧牲多能性為代價,細胞的分化能力受損[6]。上述研究證明轉入外源性基因的方法只能獲得亞穩態的原始多能性干細胞,且具有安全性隱患。如果能夠不依賴基因操作,僅通過改變培養條件就實現干細胞系多能性狀態的轉化,則有益于推動這類干細胞在再生醫學中的應用。2013年12月,Gafni[7]和Chan[8]兩個課題組分別報道利用培養條件實現了hESCs的多能性狀態轉化,其培養體系能將現有的多能性干細胞從primed狀態逆轉到naive狀態。2014年3月,Ware等利用2i+bFGF也獲得了具有原始多能性特征的hESCs,但是該方法并且依賴bFGF,撤除bFGF后細胞培養過程中始終存在背景分化[10]。這些研究組所采用的naive hESCs培養條件不盡相同,通過分析利用這些不同培養體系獲得的“naive”多能性干細胞的基因表達譜,結果提示它們之間基因表達存在明顯差異,并且部分mESCs高表達的原始多能性相關基因在轉化后的hESCs中無明顯差別。我們參考并改良了naive hESCs培養體系,也獲得了具有原始多能性特征的hESCs,細胞重新獲得兩條活化的X染色體,單克隆形成率明顯增高,DNA甲基轉移酶活性增高等。但與mESCs比較,多能性基因表達模式也不盡相同,因此這些具有naive特征的hESCs是否為真正意義上的原始多能性干細胞還有待考證,而上述培養條件中哪些是naive多能性維持的關鍵因素、它們發揮作用的分子機制以及不同培養體系造成的基因表達差別是否影響其生物學功能和安全性等問題還需要進一步研究。
目前的研究結果表明人類原始態和始發態這兩種多能性狀態維持的分子機制存在差異,與已知小鼠相應多能性狀態維持的分子機制也不盡相同。如果能更好地認識上述問題,不僅可以幫助我們了解人類早期胚胎發育規律、多能性狀態建立的過程和分子機制,還能夠幫助我們建立原始多能性的hESCs,拓展hESCs在再生醫學以及遺傳性疾病機理研究上的應用。
[1] EVANS J,KAUFMAN H.Establishment in culture of pluripotential cells from mouse embryos[J].Nature,1981,(5819)292:154-156.
[2] BRONSG,SMITHERS E,TROTTER W,et al.Derivation of pluripotent epiblast stem cells from mammalian embryos[J].Nature,2007,448(7150):191-195.
[3] GINISI,LUO Y,MIURA T,et al.Differences between human and mouse embryonic stem cells[J].Developmental Biology,2004,269(2):360-380.
[4] KOESTENBAUER S,ZECH H,JUCH H,et al.Embryonic stem cells:similarities and differences between human and murine embryonic stem cells[J].American Journal of Reproductive Immunology,2006,55(3):169-180.
[5] HANNA J,CHENG W,SAHA K,et al.Human embryonic stem cells with biological and epigenetic characteristics similar to those of mouse ESCs[J].Proc Natl Acad Sci,USA 2010,107(20):9222-9227.
[6] BUECHER C,CHEN H,POLO M,et al.A murine ESC-like state facilitates transgenesis and homologous recombination in human pluripotent stem cells[J].Cell Stem Cell,2010,6(6):535-546.
[7] GAFNI O,WEINBERGER L,MANSOUR A,et al.Derivation of novel human ground state naive pluripotent stem cells[J].Nature,2013,504(7479):282-286.
[8] CHAN S,GOKE J,NG H,et al.Induction of a human pluripotent state with distinct regulatory circuitry that resembles preimplantation epiblast[J].Cell Stem Cell,2013,13(6):663-675.
[9] XU Y,ZHU X,HAHM S,et al.Revealing a core signaling regulatory mechanism for pluripotent stem cell survival and self-renewal by small molecules[J].Proc Natl Acad Sci USA,2010,107(18):8129-8134.
[10] WARE B,NELSON A,MECHAM B,et al.Derivation of naive human embryonic stem cells[J].Proc Natl Acad Sci USA,2014,111(12):4484-4489.
[11] YING L,WRAY J,NICHOLS J,et al.The ground state of embryonic stem cell self-renewal[J].Nature,2008,453(7194):519-523.
[12] NICHOLSJ,JONES K,PHILLIPS M,et al.Validated germline-competent embryonic stem cell lines from nonobese diabetic mice[J].Nature Medicine,2009,15(7):814-818.
[13] HANNA J,MARKOULAKI S,MITALIPOVA M,et al.Metastable pluripotent states in NOD mouse derived ESCs[J].Cell Stem Cell,2009,(6):513-524.
[14] BUEHR M,MEEK S,BLAIR K,et al.Capture of authentic embryonic stem cells from rat blastocysts[J].Cell,2008,135(7):1287-1298.
[15] LI P,TONG C,MEHRIAN-SHAI R,et al.Germline competent embryonic stem cells derived from rat blastocysts[J].Cell,2008,135(7):1299-1310.
猜你喜歡
商品與質量(2021年43期)2022-01-18 05:31:22
杭州(2020年23期)2021-01-11 00:54:42
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
中國衛生(2015年12期)2015-11-10 05:13:40
現代企業(2015年1期)2015-02-28 18:43:18
汽車零部件(2014年5期)2014-11-11 12:24:28
新高考·高一物理(2014年1期)2014-09-18 01:26:07
浙江人大(2014年1期)2014-03-20 16:19:53
終身教育研究(2012年4期)2012-03-25 10:41:11

- 激光生物學報的其它文章
- 流式細胞儀分析癌細胞對肥大細胞募集的影響*
- 去泛素化酶與腫瘤*