青海湖區大型底棲動物群落結構與空間分布格局
2014-03-17 00:38:50孟星亮何玉邦宋卓彥熬鴻毅張虎蔣小明
水生生物學報 2014年5期
孟星亮何玉邦宋卓彥熬鴻毅張 虎蔣小明
(1.中國科學院水生生物研究所, 武漢 430072; 2. 中國科學院大學, 北京 100049; 3. 青海湖國家級自然保護區管理局, 西寧 810008)
青海湖區大型底棲動物群落結構與空間分布格局
孟星亮1,2何玉邦3宋卓彥1,2熬鴻毅1張 虎3蔣小明1
(1.中國科學院水生生物研究所, 武漢 430072; 2. 中國科學院大學, 北京 100049; 3. 青海湖國家級自然保護區管理局, 西寧 810008)
為了解青海湖大型底棲動物的群落結構特征、空間分布格局及關鍵驅動力, 于2011—2012年分4次對青海湖區主體湖、3個子湖和5條入湖河流進行了系統調查。共采集到底棲動物28個分類單元, 隸屬于3門5綱15科, 主體湖區、咸水和淡水子湖及入湖河流分布有9、5、9和15種; 它們的密度分別為455.8、1562.7、741.3和331.2 ind./m2; 生物量為1.18、3.70、2.75和4.53 g/m2。優勢度分析顯示, 喜鹽搖蚊為主體湖區絕對優勢種(占總密度的83.32%)。Surfer模擬顯示, 主體湖區東南部沿岸帶的密度和生物量最高, 湖心區最低; 高峰值主要出現在布哈河、菜擠河等大型河流入湖的河口附近。CCA分析表明: 青海湖區大型底棲動物的分布主要受水體pH、水深、總氮和鹽度等環境因子的驅動。
大型底棲動物; 空間格局; 青海湖; 多因子分析
青海湖是我國最大的高原咸水湖, 由于其獨特的高原環境和豐富的生物多樣性, 在1992年被列入拉姆莎爾保護公約中著名的濕地保護區, 同時分別在1997、2006和2007年被國務院和國家建設部批準為國家級自然保護區、國家自然名錄以及國家級水產種質資源保護區[1]。然而近50年來, 由于受氣候變暖和人類活動影響, 青海湖出現了一系列的生態問題, 如水位下降、沙漠化加劇、草場面積退化、魚類資源驟減等, 這促使國內外學者對青海湖現狀愈加關注[2]。目前為止, 國內有關青海湖的研究主要集中在氣候變化[3,4]、水位狀況[5,6]、魚類資源[7]和水生植物[8]等領域。但迄今為止, 有關該湖底棲動物的研究卻相對較少, 現有研究也大多僅集中于種類描述[1,9], 而罕有關于底棲動物群落結構和格局的研究。
大型底棲動物是湖泊生態系統中的重要生態類群, 在促進水體生態系統的營養物質循環和能量流動、調節底質與水體的物質交換以及加速水體凈化、改善水體微環境等方面起著十分重要的作用[10—13]。此外, 底棲動物作為魚類、鳥類等動物的天然餌料,其群落結構的變化直接影響著高營養級動物的生長和繁殖[14]。鑒于其高原咸水湖泊的特殊性及大型底棲動物的未知現狀, 本研究于2011年6月初和8月底以及2012年6月中旬和8月底, 分4個時間段對青海湖及其周邊水體的底棲動物進行了調查, 以期闡明該湖區底棲動物群落結構和空間分布現狀, 為青海湖湖區管理提供科學依據。
1 材料與方法
1.1 樣點設置與湖泊概況
湖泊概況 青海湖 (N36°32′-37°15′, E99°36′-100°47′) 又名“庫庫淖爾”, 位于青海省東北部青海湖盆地。主體湖水域面積 4340.0 km2(平均高程3193.92 m), 平均湖長為109.0 km, 平均湖寬39.8 km (最大寬67.0 km), 平均水深17.9 m (最大水深27.0 m)。湖區氣候屬高原高寒半干旱氣候, 年均氣溫 1.2 , ℃多年平均降水量336 mm。青海湖于每年11月進入冰封期至次年3月開始解凍, 冰期歷程100—129d。湖水主要依賴地表徑流和湖面降水補給, 主要入湖河流有布哈河、巴戈烏蘭河、沙柳河、黑馬河、甘子河、倒淌河等, 其徑流量約占入湖總徑流量的95%。布哈河最大, 長約286 km, 流域面積14384 km2,其徑流量約占入湖總徑流量的45%[15]。其中金沙灣部分與主體湖區季節性連通, 尕海與主體湖完全隔離且無明顯河流補給, 因而現在這兩個子湖仍然呈現咸水湖泊特征; 洱海與主體湖完全隔離且有倒淌河補給, 如今已演變成為一個淡水子湖。
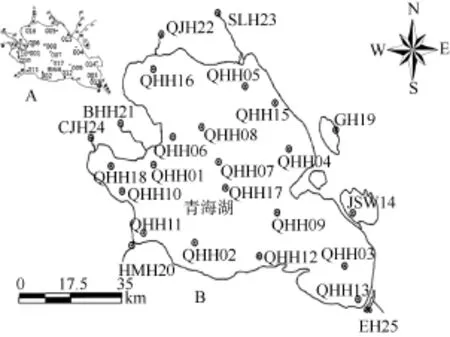
圖1 青海湖及其附近水體底棲動物和水體理化參數的樣點分布Fig. 1 Sampling sites of benthic organisms and physical-chemical parameters in Qinghai Lake
樣點設置 樣點布設上綜合考慮了主體湖區、2 個咸水子湖、1 個淡水子湖及入湖河流4 類不同鹽度類型生態區域。主體湖區大體沿用了1962年樣點 (圖 1A), 布設 17個采樣點(QHH01-18, 除JSW14外)。入湖河流設置了5個采樣點, 其中黑馬河、布哈河、泉吉河、沙柳河和菜擠河樣點分別對應HMH20、BHH21、QJH22、SLH23和CJH24。子湖設置了3個采樣點, JSW14、GH19和EH25分別對應金沙灣、尕海和洱海3個子湖 (圖1B)。
1.2 樣品的采集與處理
主體湖區使用 1/16 m2的改良 Peterson 采泥器進行底棲動物樣品采集, 每個樣點采集2次, 泥樣經40目的篩網篩洗后, 置于密封袋內帶于室內, 當天置入白色解剖盤中分揀。隨后置入50 mL樣品瓶中,并用 10%的福爾馬林固定, 帶回實驗室進行種類鑒定、計數和稱濕重(其中軟體動物為帶殼濕重)。入湖河流和子湖則使用0.09 m2的Surber網進行, 將采得樣品置于40目的分樣篩中篩洗, 此后處理方法同上。
1.3 水體理化指標的測定
用5 L采水器采取表層水樣(0.5 m), 當天帶回實驗室測定。堿度(ALK)、化學需氧(COD)、總氮(TN)、氨氮(NH4-N)、亞硝酸氮(NO2-N)、硝氮(NO3-N)、總磷(TP)、磷酸鹽(PO4-P)測定方法參照《水和廢水監測分析方法》[16]。其他指標溫度(T)、pH、溶解氧(DO)、水深(H)、電導率(EC)、鹽度(SAL)在現場使用便攜式水質分析儀測定, 透明度(SD)用Secchi盤測定。
1.4 數據處理
以底棲動物相對密度≥5%作為確定優勢種的標準[17, 18]。
使用單因素方差分析(One-way ANOVA)比較4類水體的理化因子、底棲動物密度、生物量、物種數和多樣性指數間的差異。為提高數據的正態性和方差齊性, 理化因子(pH、溶解氧和氨氮除外)與Shannon-Wiener 和Margalef指數分別進行lg(x + 1)轉換。若單因素方差分析的組間差異顯著, 則進行多重比較; 如果方差同質, 則進行 LSD (the least significant difference)檢驗; 如果方差不具同質性, 就選用Games-Howell檢驗[19], 以上處理均在SPSS 13.0軟件中完成。
底棲動物 Shannon-Wiener和 Margalef 多樣性指數計算采用以下公式:

Margalef 多樣性指數: Margalef index (d) = (S–1)/lnN,其中, H ′: Shannon-Wiener 多樣性指數; S: 樣本中總分類單元數; Pi: 物種 i在樣本中的相對豐度; N:樣本中總個體數。
本次調查主要在主體湖區, 因而作者著重分析了主體湖區底棲動物的密度和生物量空間分布格局,空間分布圖使用普通克里格插值法, 在Surfer 8.0軟件中完成[20]。
探討底棲動物分布和水環境因子的關系采用 R軟件(2.15版本), “vegan” 軟件包完成。首先, 對25個樣點4次水體理化因子和物種數據分別進行平均,然后分別對非正態分布的環境因子(pH、氨氮和礦化度除外)分別進行 lg (x+1)轉換。接著利用函數permutest( )進行 Monte Carlo 轉置, 檢驗環境因子與物種分布間的相關性。其次使用envfit( )涵數檢驗每個環境因子的相關性, 然后將物種豐富度與環境因子不相關的(P>0.05)環境因子剔除, 同時利用vif.cca( )函數剔除較高變異波動系數(>20)的變量。隨后分別對物種數據進行 lg (x+1) 轉換, 保留在各樣點出現的頻度>10%且相對密度≥1%的物種, 以消除稀有種的影響[21—22]。最后使用decorana( )函數對底棲動物群落分布進行除趨勢對應分析 (DCA),顯示單峰模型 (gradient length > 4 standard units) 更為適合群落組成與環境因子之間關系的分析,因此采用cca( )函數進行了典范對應分析(CCA)[23,24]。
2 結果
2.1 青海湖湖區環境因子特征
在4類水體中, 多數環境因子差異顯著(表1)。其中, SAL、EC、MIN、ALK表現為咸水子湖>主體湖>淡水子湖和河流; 而 SiO2、TP、NO3-N則為入湖河流>淡水子湖>主體湖區和咸水子湖; TN 和CODMn在淡水子湖出現最高值, 河流和咸水子湖居中, 而在主體湖區最低。總體而言, 主體湖區的營養鹽濃度處于貧營養水平[25]; 而 3個子湖和入湖河流的營養鹽濃度相對較高。
2.2 青海湖湖區底棲動物群落結構
本文共記錄28個分類單元, 隸屬于3門5綱15科, 其中主體湖區9種, 淡水子湖9種, 咸水子湖5種, 入湖河流 15種(表 2)。主體湖、咸水子湖、淡水子湖和入湖河流的平均密度分別為455.8、1562.7、741.3和 331.2 ind./m2; 平均生物量分別為 1.18、3.70、2.75和4.53 g/m2(圖2)。
密度、生物量和物種數在4類水體間差異不顯著 (P > 0.05)。相對而言, 在密度方面咸水子湖最高,淡水子湖次之, 河流和主體湖最低; 生物量方面河流和咸水子湖較高, 主體湖最低, 淡水子湖居中;物種數河流和淡水子湖較高, 主體湖與咸水子湖較低(圖 2)。

表1 2011—2012年青海湖湖區主要水質參數Tab. 1 The water quality parameters in Qinghai Lake area from 2011 to 2012 (mean ± SD)
群落組成方面 4類水體差異較大, 主體湖區以喜鹽搖蚊(Chironomus salinarius)、前突搖蚊(Procladius sp.)和鉤蝦(Gammarus sp.)為優勢種, 三者相對密度分別為83.32%、10.58%和5.11%, 占總生物量的百分比分別為 81.49%、7.91%、10.6%。3個子湖主要以水生昆蟲為主, 其中優勢種類均為蠓(Ceratopogonidae spp.)及水甲(Hygrobiidae spp.), 二者在咸水和淡水子湖相對密度分別為 80.55%、18.77%和79.82%、19.47%。而入湖河流的優勢種為
鉤蝦、扁蜉(Heptageniidae spp.)、特氏直突搖蚊(Orthocladious thienemanni)和大蚊 (Tipulidae spp.),相對密度分別為62.80%、9.18%、8.21%、7.25%。

表2 青海湖湖區底棲動物物種名錄Tab. 2 List of the species of macroinvertebrates in Qinghai Lake area

圖2 四類水體平均密(A)、生物量(B)和物種數(C)的比較(Mean±SD)Fig. 2 Comparison of density, biomass and the number of the species between 4 types of water bodies in Qinghai Lake area Abbreviation: MB: main body; SAL: saline adjacent lake; FAL: freshwater adjacent lake; Irs: inflowing rivers
2.3 青海湖湖區底棲動物生物多樣性
4類水體的多樣性較低(圖 3)。Shannon-Wiener指數在4類水體之間差異不明顯(P>0.05); Margalef指數 4類水體之間相比, 主體湖區、咸水子湖與河流間差異顯著(P<0.05)。相對而言河流和淡水子湖多樣性較高, 咸水子湖和主體湖較低。
2.4 主體湖區大型底棲動物空間分布
底棲動物的密度和生物量呈現大體一致的空間分布格局, 表現為西南部大于東北部 (圖4a、b);兩者從湖心區域依次向沿岸區域逐漸增加, 最高點位于西南角菜擠河河口附近, 密度和生物量分別達2000 ind./m2、4 g/m2以上, 次高點位于泉吉河河口(800 ind./m2和 2.5 g/m2); 優勢種喜鹽搖蚊與底棲動物總體的密度和生物量空間分布大體一致(圖4c、d)。2.5 青海湖湖區底棲動物與水環境關系

圖3 青海湖區4類水體Shannon (A)和Margalef (B)指數的比較(Mean±SD)Fig. 3 Comparison of Shannon and Margalef Indice between the 4 types waterbodies in Qinghai Lake area Abbreviation: MB: main body; SAL: saline adjacent lake; FAL: freshwater adjacent lake; Irs: inflowing rivers

圖4 青海湖大型底棲動物總體的密度(a)和生物量(b)及優勢種喜鹽搖蚊的密度(c)和生物量(d)的空間分布Fig. 4 Density (a) and biomass (b) spatial distribution of total macrozoobenthos and density (c) and biomass (d) space distribution of Chiroindmus salinarius in Qinghai Lake
利用 envift()函數對 15個環境因子進行篩選,最后有7個關鍵環境因子進入CCA模型(表3)。主要環境因子的解釋率為65.2%, CCA 第一和第二排序軸的解釋率分別為 24.5%和 16.6%, 前兩軸累積解釋率為 41.1%。其中 pH (r2=0.8683)、水深(r2= 0.8527)、總氮(r2=0.7762)和硝氮(r2=0.7542)與底棲動物群落分布顯著相關, 其次是鹽度、礦化度和溫度(表3)。

表3 青海湖湖區底棲動物與關鍵環境因子Monte Carlo檢驗結果Tab. 3 Results of Monte Carlo test between the key environmental factors and the distribution of the zoobenthos in Qinghai Lake area
水深、pH和鹽度與CCA第一排序軸呈顯著負相關, 礦化度、總氮和硝氮與第一軸呈顯著正相關; CCA第二排序軸則與溫度和鹽度呈顯著正相關。物種與環境因子相互關系表明, 水深、pH和鹽度顯著影響了喜鹽搖蚊、前突搖蚊(Procladius sp.)等物種的空間分布(圖 5)。水甲(Hygrobiidae spp.)、蠓(Ceratopogonidae spp.)和梯形多足搖蚊(P. scalaenum)喜溫的昆蟲與溫度和總氮正相關。大蚊 (Tipulidae spp.)和帶襀(Taenionema sp.)等物種與鹽度呈負相關,與硝氮呈正相關, 這與該兩個物種偏好在淡水、營養鹽類相對較為豐富的溪流中生活相吻合(圖5)。

圖5 青海湖底棲動物群落結構與水環境關系的CCA分析Fig. 5 The CCA analysis between macrozoobenthos and the key environmental factors in Qinghai lake
3 討論
3.1 青海湖湖區大型底棲動物歷史動態
青海湖主體湖區的底棲動物物種組成單一、群落結構也較為簡單, 主要以搖蚊為主, 這與我國北方許多鹽堿湖泊的底棲動物群落組成較為相似[26,34]。就主體湖區而言, 與20世紀60年代相比, 本研究底棲動物的總分類單元減少了10個, 科級水平減少了6個科, 屬級水平減少了 10個屬, 其中軟體動物下降了3種、寡毛類下降了2種、其他水生昆蟲下降了4種、搖蚊類下降了1種; 平均生物量和密度略有上升[27]; 而對搖蚊類而言, 本此調查與20世紀80年代減少了1個屬5個分類單元[28](表4)。就整個青海湖區而言, 底棲動物的物種組成雖然比20世紀90年代在科級水平略有提升, 但其減少了 5屬 16個分類單元[29]; 搖蚊類與20世紀80年代相比減少了9屬[28](表4)。
青海湖底棲動物方面的研究資料極度匱乏, 就本文數據, 這50年來青海湖底棲動物生物多樣性呈現下降的態勢。這可能一方面與鹽度、水域面積變化等有關, 另一方面可能與本研究對河流及子湖調查力度較為薄弱有關[30,31]。
3.2 青海湖底棲動物與水環境之間的關系
許多研究表明, 水深是影響靜水生態系統底棲動物群落結構的關鍵因子, 水深的改變會導致一些重要理化因子(如溫度和溶氧等)發生變化, 從而間接影響底棲動物的分布[32—34]。主體湖Surfer模擬結果顯示, 底棲動物密度和生物量的空間分布呈現,從西北部沿岸帶向湖心深水區逐漸遞減的趨勢, 且水深較淺的西南部底棲動物的密度和生物量遠高于水深較深的東北部。此外, 湖心帶相比湖泊沿岸帶通常具有較高的底棲動物多樣性和生產力, 因為沿岸帶豐富的有機質來源為底棲動物提供了充沛的食物, 更加有利于底棲動物生存[35]。
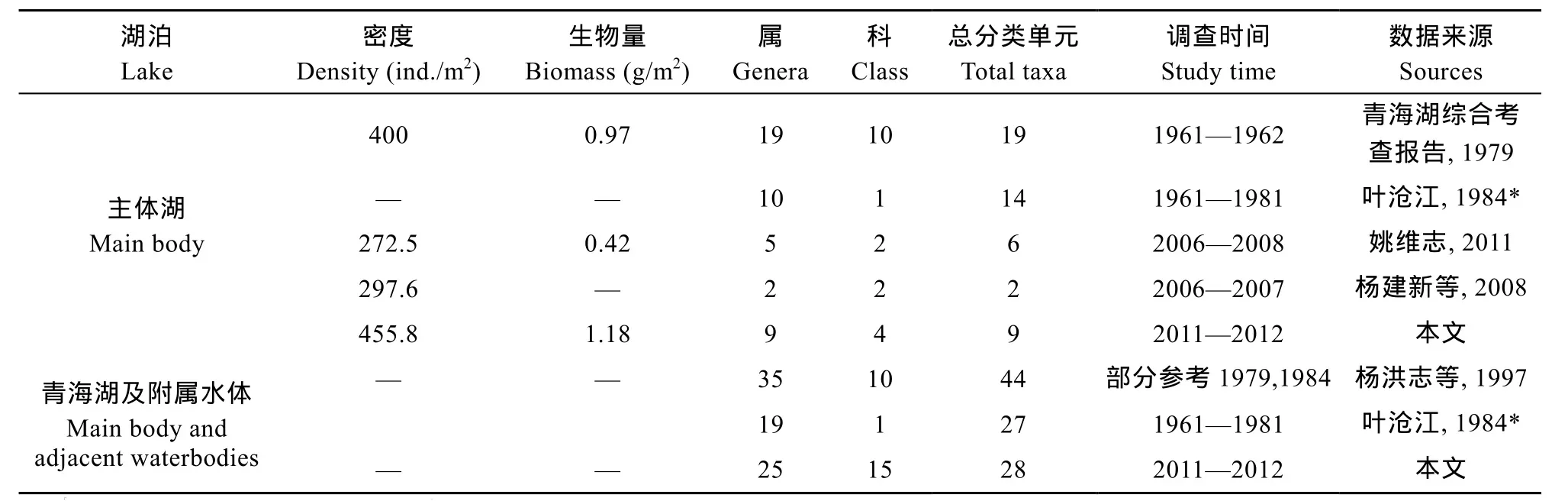
表4 不同階段青海湖調查數據Tab. 4 The data of Qinghai Lake in different investigation periods
pH是另一個顯著影響底棲動物分布格局的環境因子, 謝祚渾等[36]研究我國北方鹽堿水域中的底棲動物指出, 不同的底棲動物對酸堿度的耐受不同,如羽搖蚊(C. plumosus)、前突搖蚊和蠓科幼蟲能夠在pH 9—10內的水體中生存, 然而其他種類大多生活于pH 7—9內的水體內。這與本文CCA分析結果一致, 扁蜉、特氏直突搖蚊、大蚊等物種與pH呈負相關, 表明這些物種對pH的耐受范圍較窄, 因而它們大多分布在pH相對較低的入湖河流中; 相反, 喜鹽搖蚊、前突搖蚊及蠓等物種大多分布于主體湖和咸水子湖中。
通常在咸水及半咸水的水體中, 鹽度往往成為影響底棲動物群落組成與分布的關鍵因素, 隨著鹽度的增加, 底棲動物的物種多樣性將逐漸降低, 而在水體鹽度值界于 5‰—8‰時, 種類組成呈現單一化的趨勢[37,38]。另外, 不同種類的底棲動物對于鹽度的耐受范圍也差異很大: 如喜鹽搖蚊作為咸水水體的廣布物種, 其可在 5‰—50‰鹽度范圍內生存,花紋前突搖蚊(P. Choreus)和蠓科的一些種類均可耐受10‰以上的鹽度; 而橢圓蘿卜螺(Radix swinhoei)和扁旋螺(Gyraulus compressus) 等螺類大多只能在低于6‰ 鹽度的水體中生存[36,39,40]。本文調查發現,金沙灣、尕海及主體湖鹽度與河流和子湖洱海差異明顯, 因而洱海和河流的物種數目、多樣性指數相比主體湖及2個咸水子湖較高。青海湖是典型的高原深水型咸水湖泊, 水體中大量鹽份的存在, 限制了其他物種的入侵與生存, 致使河流及附近淡水湖泊中較為豐富的底棲動物種類無法擴散到主體湖,卻為喜鹽的喜鹽搖蚊和前突搖蚊等物種提供了良好的生存環境。因此, 鹽度可能是導致青海湖底棲動物物種數目單一的重要原因。
致謝:
感謝中國科學院水生生物研究所王丑明在采樣和標本鑒定工作中給予的幫助。
[1] Yao W Z. Study on the resource of feed organisms in Qinghai Lake during summer [D]. Southwest University PH.D thesis. 2011 [姚維志. 青海湖夏季餌料生物資源研究.西南大學博士學位論文. 2011]
[2] Chen G C, Peng M, Zhou L H, et al. Relationship between ecological environment change and human activity in Qinghai Lake region: A preliminary study [J]. Chinese Journal of Ecology, 1994, 13(2): 44—49 [陳桂琛, 彭敏, 周立華, 等. 青海湖地區生態環境演變與人類關系的初步研究. 生態學雜志, 1994, 13(2): 44—49]
[3] Feng S, Tang M C, Zhou L S. The water level of Qinghai Lake change in recent 600 years [J]. Journal of Lake Sciences, 2000, 12(3): 205—210 [馮松, 湯懋蒼, 周陸生.青海湖近 600年的水位變化. 湖泊科學, 2000, 12(3): 205—210]
[4] Zhen E L, Shen J, Wang S M, et al. Lake sediment records of climate and environment evolution of Qinghai Lake in recent 900 years [J]. Journal of Lake Sciences, 2002, 14(1):32—38 [張恩樓, 沈吉, 王蘇民, 等. 青海湖近900年來氣候環境演化的湖泊沉積記錄. 湖泊科學, 2002, 14(1): 32—38]
[5] Ma Y. Correlation analysis between the changing of water level and the climate elements of Qinghai Lake [J]. Journal of Lake Sciences, 1996, 8(2): 103—106 [馬鈺. 青海湖水位變化與湖區氣候要素的相關分析. 湖泊科學, 1996, 8(2): 103—106]
[6] Yang H Y, Jia S F. Water balance and water resources allocation of Qinghai Lake [J]. Journal of Lake Sciences, 2003, 15(1): 35—40 [燕華云, 賈紹風. 青海湖水量平衡分析與水資源優化配置研究. 湖泊科學, 2003, 15(1): 35—40]
[7] Hu A, Zhu S Q, Wang J L, et al. Fish Fauna of Qinghai Lake and the Biology of Gymnocypris przewalskii [M]. Beijing: Science Press. 1975, 94 [青海省生物研究所編. 青海湖地區的魚類區系和青海湖裸鯉的生物學. 北京: 科學出版社. 1975, 94]
[8] Chen Y D. Studies on the Potamogetonaceae in Qinghai Lake [J]. Acta Hydrobiologica Sinica, 1987, 11(3): 228—235 [陳耀東. 青海湖眼子菜科植物的研究. 水生生物學報, 1987, 11(3): 228—235]
[9] Yang J X, Qi H F, Si J Q, et al. The summer investigation of aquatic organisms in Qinghai Lake [J]. Qinghai Science and Technology, 2008, 6: 19—24 [楊建新, 祁洪芳, 史建全, 等.青海湖夏季水生生物調查. 青海科技, 2008, 6: 19—24]
[10] Covich A P, Palmer M A, Crowl T A. The role of benthic invertebrate species in freshwater ecosystems: zoobenthic species influence energy flows and nutrient cycling [J]. BioScience, 1999, 49(2): 119—127
[11] Lindegaard C. The role of zoobenthos in energy flow in deep, oligotrophic Lake Thingvallavatn, Iceland [J]. Hydrobiologia, 1992, 243(1): 185—195
[12] Lindegaard C. The role of zoobenthos in energy flow in two shallow lakes [J]. Hydrobiologia, 1994, 275(1): 313—322
[13] Bonsdorff E. Drifting algae and zoobenthos effects on settling and community structure [J]. Netherlands Journal of Sea Research, 1992, 30: 57—62
[14] Schilling E G, Loftin C S, Huryn A D. Effects of introduced fish on macroinvertebrate communities in historically fishless headwater and kettle lakes [J]. Biological Conservation, 2009, 142(12): 3030—3038
[15] Wang S M, Dou H S. Lakes in China [M]. Beijing: Science Press. 1998, 472—473 [王蘇民, 竇鴻身. 中國湖泊志. 北京: 科學出版社. 1998, 472—473]
[16] The Editor of Water and Wastewater Monitoring Analysis Method. Water and Wastewater Monitoring Analysis Method (4th edition) [M]. Beijing: China Environmental Science Press. 2002, 211—279 [國家環保局, 水和廢水監測分析方法編委會編. 水和廢水監測分析方法(第四版)北京: 中國環境科學出版社. 2002, 211—279]
[17] Bunn S E, Edward D H, Loneragan N R. Spatial and temporal variation in the macroinvertebrate fauna of streams of the northern jarrah forest, Western Australia: community structure [J]. Freshwater Biology, 1986, 16: 67—92
[18] Xiong J, Jiang X M, Wang C M, et al. Community variation of macrozoobenthos and bioassessment of Dongqian Lake, Ningbo [J]. Research of Environmental Sciences, 2012, 25(3): 282—289 [熊晶, 蔣小明, 王丑明, 等. 寧波東錢湖大型底棲動物群落動態及水質生物學評價. 環境科學研究, 2012, 25(3): 282—289]
[19] Beckmann C F, DeLuca M, Devlin J T, et al. Investigations into resting-state connectivity using independent component analysis [J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2005, 360: 1001—1013.
[20] Zeng Z X, Chen H X. How to use Sufer 8.0 to draw isoline map [J]. Guangdong Meteorology, 2006, 3: 64—65 [曾志雄,陳慧嫻. 如何使用Sufer 8.0畫等值線圖. 廣東氣象, 2006, 3: 64—65]
[21] Flores L N, Barone R. Phytoplankton dynamics in two reservoirs with different trophic state (Lake Rosamarina and Lake Arancio, Sicily, Italy) [J]. Hydrobiologia, 1998, 369/370: 163—178
[22] Muylaert K, Sabbe K, Vyverman W. Spatial and temporal dynamics of phytoplankton communities in a freshwater tidal estuary (Schelde, Belgium) [J]. Estuarine, Coasta and Shelf Science, 2000, 50: 673— 687
[23] Leps J, Smilauer P. Multivariate Analysis of Ecological Data Using CANOCO [M]. University of Cambridge Press. 2003, 51
[24] Lai J S, Mi X C. Ordination analysis of Ecological data using Vegan package in R [J]. Advances in Biodiversity Conservation and Research in China IX, 2005, 332—343 [賴江山, 米湘成. 基于Vegan軟件包的生態學數據排序分析.中國生物多樣性保護與研究進展IX, 2005, 332—343]
[25] Wang M C, Liu X Q, Zhang J H. Evaluate method and classification standard on lake eutrophication [J]. Environmental Monitoring in China, 2002, 18(5): 47—49 [王明翠, 劉雪芹, 張建輝. 湖泊富營養化評價方法及分級標準. 環境科學研究, 2002, 18(5): 47—49]
[26] Lan C J, Shen Y, Wang B X, et al. Investigation of plants and benthic macroinvertebrates of in inner Mongolia-Xinjiang Plateau [J]. Journal of Lake Science, 2010, 22(6): 888—893 [蘭策介, 沈元, 王備新, 等. 蒙新高原湖泊高等水生植物和大型底棲無脊椎動物調查. 湖泊科學, 2010, 22(6): 888—893]
[27] Lanzhou Institute of Geology CIS. The Report of Integrated Investigation on Qinghai Lake [M]. Beijing: Science Press. 1979, 50—54 [中國科學院蘭州地質研究所等編. 青海湖綜合考察報告. 北京: 科學出版社.1979, 50—54]
[28] Ye C J. The larvae of some Tendipedidae midge (Diptera) from Qinghai Lake and its’ river system [J]. Acta Biologica Plateau Sinica, 1984, 2: 67—74 [葉滄江. 青海湖及其附屬水系的搖蚊幼蟲. 高原生物學集刊, 1984, 2: 67—74]
[29] Yang H Z, Wang J L. Analysis on the benthos and their productivity in Qinghai Lake [J]. Qinghai Science and Technology, 1997, 4(3): 36—39 [楊洪志, 王基琳. 青海湖底棲生物及其生產力分析. 青海科技, 1997, 4(3): 36—39]
[30] Shen F, Kuang D B. Remote sensing investigation and survey of Qinghai Lake in the past 25 years [J]. Journal of Lake Sciences, 2003, 15(4): 289—296 [沈芳, 匡定波. 青海湖最近 25年變化的遙感調查與研究. 湖泊科學, 2003, 15(4): 289—296]
[31] Tian R W. The hydrologic condations of the Qinghai Lake [J]. Tansactions of Oceanology and Limnology, 1988, 4: 18—25 [田任午. 青海湖的水文概況. 海洋湖沼通報. 1988, 4: 18—25]
[32] Petridis D, Sinis A. Benthic maerofauna of Tavropos Reservoir (central Greece) [J]. Hydrobiologia, 1993, 262: 1—12
[33] Baudo R, Ochhipinti A, Nocentini A M.Benthos of Lake Orta in the year 1996 [J]. Journal of Limnology, 2001, 60(2) : 241—248
[34] Zhang C W, Zhang T L, Zhu T B, et al. Community structure of Macrozoobenthos and its relationship with environmental factors in lake Hongze [J]. Journal of Hydroecology, 2012, 33(3): 27—33 [張超文, 張堂林, 朱挺兵, 等. 洪澤湖大型底棲動物群落結構及其與環境因子關系. 水生態學雜志, 2012, 33(3): 27—33]
[35] Horne A J, Goldman C R (editors). Limnology (2nd edition) [M]. McGraw-Hill Inc, New York. 1994, 17—19
[36] Xie Z H, Zhou Y B. Zoobenthos in inland saline waters from autonomous region northern parts of China [J]. Journal of Dalian Fisheries University, 2002, 17(3): 176—186 [謝祚渾,周一兵. 中國北方鹽堿水域中的底棲動物. 大連水產學院學報, 2002, 17(3): 176—186]
[37] Cartier V, Claret C, Garnier R, et al. How salinity affects life cycle of a brackish water species, Chironomus salinarius KIEFFER (Diptera: Chironomidae) [J]. Journal of Experimental Marine Biology and Ecology, 2011, 405: 93—98
[38] Williams W D. Salinity as a determinant of salt lake fauna: a question of scale [J]. Hydrobiologia, 1990, 197: 257—266
[39] Gainey Jr. L F, Greenberg M J. Physiological basis of the species abundance-salinity relationship in Molluscs: a speculation [J]. Marine Biology, 1977, 40: 41—49
[40] Wang B Q, Xue J Z, Liu L, et al. Community structure of mcacrobenthos in different salt ponds of Altay prefecture, Xinjiang [J]. Arid Land Geography, 2012, 35(2): 265—273 [王寶強, 薛俊增, 劉婧, 等. 新疆阿勒泰地區不同鹽池大型底棲動物的群落結構. 干旱區地理, 2012, 35(2): 265—273]
STRUTURE AND SPATIAL DISTRIBUTION PATTERNS OF MACROZOOBENTHOS IN QINGHAI LAKE AREA
MENG Xing-Liang1,2, HE Yu-Bang3, SONG Zhuo-Yan1,2, AO Hong-Yi1, ZHANG Hu3and JIANG Xiao-Ming1
(1. Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Qinghai Lake National Nature Reserve Administration Bureau, Xining 810008, China)
To study macrozoobenthos community structures, distributions and key underlying drive forces of the Qinghai Lake (QL), one of typical saline lakes in Tibetan Plateau, the samples of benthic animals were taken via four different times during 2011—2012 from QL area, including main body (MB), 3 adjacent lakes (ALs) and 5 inflowing rivers (IRs). We identified 28 taxa: 9 in MB, 5 in 2 saline ALs, 9 in 1 freshwater AL, and 15 in IRs. The mean density of 4 types of water bodies were 455.8, 1562.7, 741.3, and 331.2 ind./m2, respectively. Biomass of these 4 different water bodies were 1.18, 3.70, 2.75, and 4.53 g/m2respectively. Chiroinomus salinarius was the major species (83.32% of the total abundance) in MB. The general spatial distribution pattern was supported by the Surfer Model, indicating that the highest value of macrozoobenthos abundance and biomass occurred in the southeast littoral zone of MB, especially in the sites near estuaries of IRs (e.g., Buha, Caiji and Quanji Rivers), and that the lowest value was in the profundal zone of MB. The CCA analysis showed that pH, water depth, total nitrogen, and salinity were the key environmental factors to affect macrozoobenthos community variance.
Macrozoobenthos; Spatial pattern; Qinghai Lake; Multivariate analysis
Q145+.2
A
1000-3207(2014)05-0819-09
10.7541/2014.123
2013-06-26;
2013-11-28
國家科技基礎條件平臺重點項目“青海湖綜合研究專題數據庫”(BSDN2009-18)資助
孟星亮(1982—), 男, 河南湯陰人; 博士研究生; 主要從事底棲動物多樣性及分類學研究。E-mail: xingliang01@163.com
蔣小明(1983—), 男, 助理研究員; 主要從事底棲動物多樣性及分類學研究。E-mail: jxm123.83@163.com
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
南大法學(2021年3期)2021-08-13 09:22:32
阿來研究(2021年1期)2021-07-31 07:39:04
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中國自行車(2018年9期)2018-10-13 06:17:10
金色年華(2016年13期)2016-02-28 01:43:27
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12

- 水生生物學報的其它文章
- GILL MEDIATES IMMUNE RESPONSES AFTER GRASS CARP REOVIRUS CHALLENGE IN GRASS CARP (CTENOPHARYNGODON IDELLA)
- MORPHOLOGICAL OBSERVATION AND RBCL SEQUENCE ANALYSIS OF A NEW SPECIES FROM CHINA, GRATELOUPIA BOAOENSIS WANG ET LUAN SP. NOV.(HALYMENIACEAE, RHODOPHYTA)
- GILL MEDIATES IMMUNE RESPONSES AFTER GRASS CARP REOVIRUS CHALLENGE IN GRASS CARP (CTENOPHARYNGODON IDELLA)
- 養殖密度對俄羅斯鱘幼魚的生長、生理和免疫指標的影響
- 一種快速構建集胞藻6803 petBD必需基因定點突變株的方法
- 大通湖環棱螺的次級生產力