水楊酸處理對冷藏杏果實細胞超微結構的影響
2014-03-08 11:02:09侯媛媛趙亞婷
食品科學 2014年14期
關鍵詞:結構
朱 璇,侯媛媛,賈 燕,趙亞婷,馬 玄
(1.新疆農業大學食品科學與藥學學院,新疆 烏魯木齊 830052;2.新疆果品加工與保鮮工程技術研究中心,新疆 烏魯木齊 830052;3.天津科技大學食品工程與生物技術學院,天津 300457)
水楊酸處理對冷藏杏果實細胞超微結構的影響
朱 璇1,2,侯媛媛1,賈 燕3,趙亞婷1,馬 玄1
(1.新疆農業大學食品科學與藥學學院,新疆 烏魯木齊 830052;2.新疆果品加工與保鮮工程技術研究中心,新疆 烏魯木齊 830052;3.天津科技大學食品工程與生物技術學院,天津 300457)
以新疆塞買提杏為試驗材料,研究了0 ℃冷藏條件下0.01 g/L的水楊酸(salicylic acid,SA)處理對杏果實不同貯藏時期超微結構的影響,以蒸餾水處理的杏果實作為對照。結果表明,在冷藏初期,SA處理杏果實和對照杏果實的細胞排列整齊,細胞壁及各細胞器結構完整,二者細胞超微結構無明顯差異。從冷藏第28天開始,杏果實細胞超微結構發生明顯變化,出現細胞器變形或降解,細胞間隙大,細胞內含物減少,細胞空腔化等現象。與對照杏果實相比,SA處理能在一定程度上減輕低溫對杏果實細胞壁、葉綠體、線粒體等的破壞,維持細胞器和膜系統的完整性,從而提高杏果實冷藏期間的抗冷性。
水楊酸;杏;冷藏;超微結構
杏(Prunus armeniacal)屬薔薇科,李屬,呼吸躍變型果實。新疆是我國杏的最大產地,據統計,2012年杏果實產量為160.7萬 t,是疆內水果總量的26.72%[1]。賽買提杏是新疆南疆地區杏果中栽種,后熟期短,采后常溫下幾天內果實變軟、品質、風味迅速下降并出現腐爛[2]。低溫貯藏可以降低杏果實的呼吸代謝,延緩后熟衰老,抑制微生物生長,從而達到延長貯藏期的目的[3]。但杏果實對低溫環境敏感,在低溫條件下貯藏較長的時間易引起冷害。冷害會導致果蔬抗病性和耐藏性下降,降低產品質量,造成食用品質劣變和腐爛,極大地限制了低溫技術的應用。因此,如何增強杏果采后對低溫的耐受性,控制冷害的發生,已成為杏貯運產業中亟待解決的問題。
近年來,已有大量研究表明甜柿[4]、油桃[5]、梅果[6]、樹莓[7]、黃瓜[8]、蟠桃[9]等果實的后熟衰老及耐貯性與其超微結構的變化有著密切的關系,但有關超微結構與果實冷害關系的報道甚少。目前,有關低溫貯藏對果蔬超微結構影響的研究主要集中在枇杷[10]、桃[11]等果實上,而針對杏果實冷藏期間細胞超微結構變化的研究還未見相關報道。
水楊酸(salicylic acid,SA)是一種廣泛存在的植物內源小分子酚類化合物,其參與植物體內多種重要的生理生化過程,它的生理作用廣泛表現在對植物生長、發育、成熟和衰老等生理過程的調控以及抗病、抗熱、抗冷、抗鹽、抗旱等抗逆反應的誘導過程中[12]。SA已被廣泛應用于增強采后果蔬貯藏的抗冷性[13-15],但已有的研究主要集中于SA處理對果實冷害的控制效果和果實品質的影響,而從細胞超微結構角度進行研究的報道較少。本實驗研究了SA處理對采后杏果實冷藏期間超微結構的影響,探討了杏果實細胞超微結構與果實冷害的關系,一方面為杏果實貯藏期間冷害的防治提供新思路,另一方面可以為SA增強采后果實低溫耐受性提供理論依據。
1 材料與方法
1.1 材料與試劑
“塞買提”杏于2013年6月采自庫車縣烏恰鎮杏果園;檸檬酸鉛 北京達濟科儀科技有限公司;丙酮(分析純) 天津永晟精細化工有限公司。
1.2 儀器與設備
DHG-9123A型電熱恒溫鼓風干燥箱 上海一恒科技有限公司;UC6超薄切片機 德國徠卡(Leica)儀器有限公司;JEOLM-1230型透射電鏡 日本電子株式會社;醋酸雙氧鈾 上海譜振生物科技有限公司。
1.3 方法
1.3.1 原料處理
“塞買提”杏果實采收后12 h內運回新疆農業大學果蔬采后生理研究室,剔除傷、病果,選擇大小、果色均勻、成熟度相近的杏果實進行處理。在預實驗的基礎上,篩選出SA對杏果實冷害控制最佳的作用質量濃度為0.01 g/L。然后將杏果實用0.01 g/L的SA以減壓方式處理(將杏果實浸入到溶液中抽氣,至壓力降到0.05 MPa時保持2 min,然后放入空氣,讓果實在常壓下繼續浸泡8 min,取出自然晾干),處理后的杏果實置于溫度為0 ℃、90%~95%相對濕度的冷庫貯藏,以蒸餾水處理的杏果實作為對照,每處理用果10 kg,重復3 次。
1.3.2 電鏡樣品制備
從采收當天開始取樣作為原始樣,入庫第7天取一次樣,第28天取一次樣,冷藏第49天取一次樣,共取樣4 次。每次取樣的部位保持一致。用雙面刀片將果皮下2 mm左右果肉切成1 mm×1 mm×2 mm大小的塊,用體積分數為2.5%戊二醛(0.1 mol/L磷酸緩沖液配制,pH 7.2)在4 ℃條件下前固定14 h。磷酸緩沖液(pH 7.2)沖洗后,用體積分數為1%餓酸(0.1 mol/L磷酸緩沖液配制,pH 7.2)4 ℃條件下后固定2 h,用磷酸緩沖再次沖洗。用體積分數30%、50%、70%、80%、90%、95%乙醇和純丙酮梯度脫水,丙酮過度。環氧樹脂Epon-812滲透包埋,30 ℃溫箱聚合12 h 后,60 ℃聚合48 h。用UC6超薄切片機切成60 nm的薄片,再經醋酸雙氧鈾及檸檬酸鉛染色,JEOLM-1230型透射電鏡觀察拍照。
2 結果與分析
2.1 杏果實采收當天透射電鏡觀察
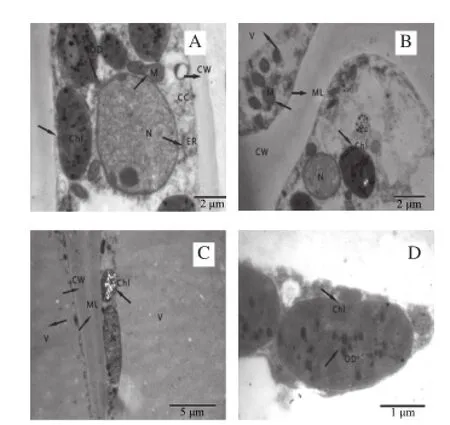
CW.細胞壁;M.線粒體;ML.中膠層;Chl.葉綠體;V.液泡;OD.嗜鋨顆粒;N.細胞核;CC.細胞內含物;ER.內質網。A.細胞整體結構,細胞壁、細胞核、葉綠體、線粒體、內質網等細胞器清晰可見,結構完整(×12 000);B.細胞壁、中膠層、液泡等結構完整,線粒體豐富(×12 000);C.細胞大液泡化,細胞質及其內含物緊貼細胞壁(×6 000);D.葉綠體基質片層和基粒片層結構完整,嗜鋨顆粒明顯(×30 000)。
通過透射電鏡觀察采收時杏果實細胞的超微結構,如圖1所示。采收當天杏果實細胞結構正常,排列整齊,各細胞器完好,質膜清晰;細胞壁結構完整,中膠層電子密度較高,初生壁緊貼中膠層兩邊;細胞明顯液泡化,液泡占細胞體積的90%以上,液泡的膜結構完整;細胞質被擠壓成一薄層緊貼細胞壁,內含大量結構完整、嵴發達的線粒體;葉綠體結構完整,基質片層與基粒片層緊密相連,整個葉綠體內部呈現一個完整的膜系統結構,嗜鋨顆粒分散其中,雙層膜結構完整;個別細胞內容物豐富,細胞核清晰可見,核仁明顯,粗面內質網及核糖體游離在細胞質中。
2.2 杏果實冷藏第7天透射電鏡觀察
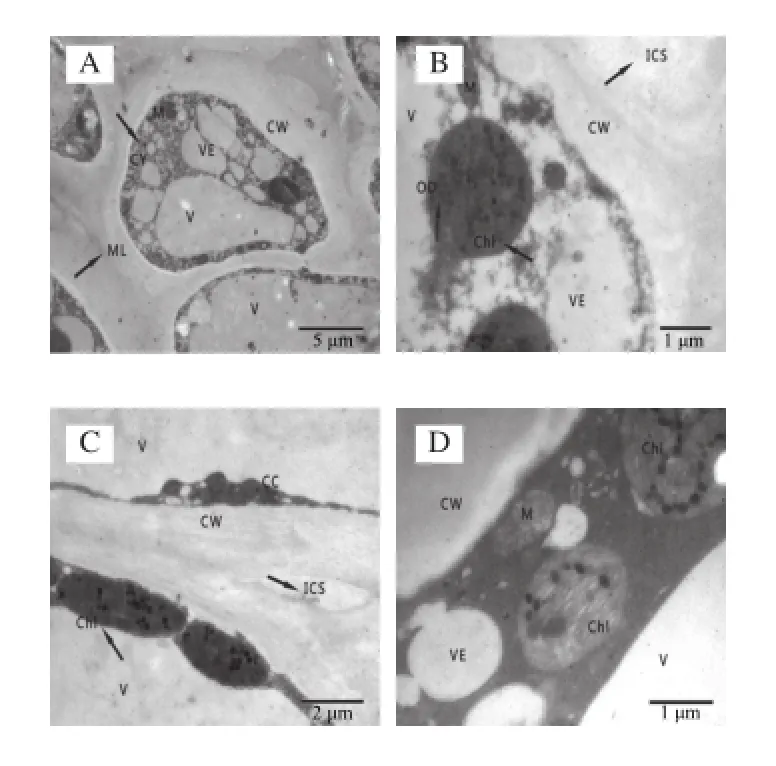
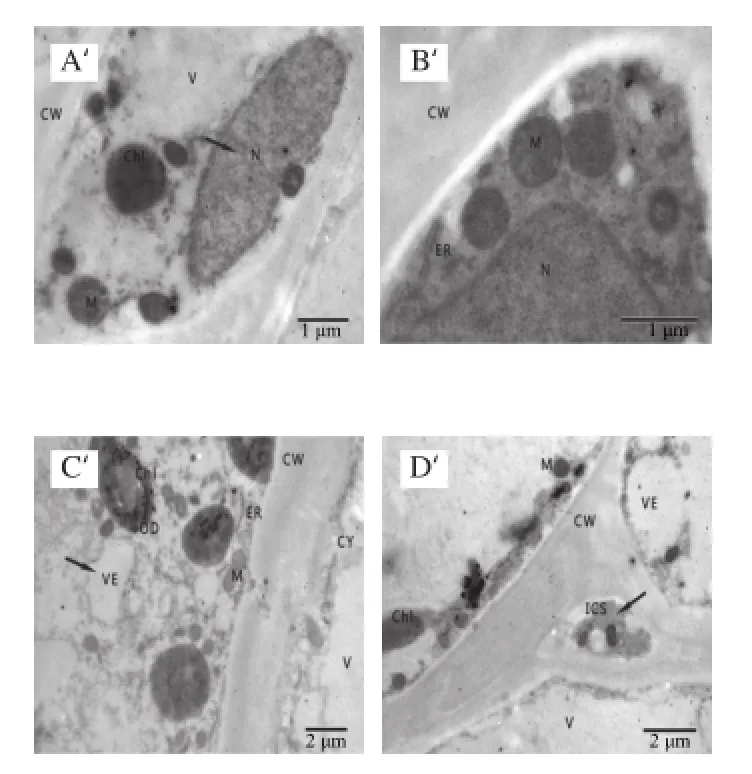
圖2 冷藏第7天杏果實超微結構Fig.2 Ultrastructure of apricot fruits after 7 days of cold storage
透射電鏡觀察表明,杏果實在冷藏的第7天時細胞間隙略有增大,大多數細胞器基本無損,排列有序,與采收時細胞結構無明顯差異。杏果實細胞葉綠體完整,部分細胞內少數葉綠體膜開始產生小泡,線粒體數量多,液泡完整,細胞質內含物緊貼細胞壁,細胞壁結構整齊,未發現降解;細胞質中有豐富的大小囊泡,液泡單層膜和細胞質膜還未受損。由圖2可知,杏果實冷藏的第7天,SA處理組和對照組果實細胞超微結構均未出現異常,且處理組與對照組之間沒有明顯差別。
2.3 杏果實冷藏第28天透射電鏡觀察
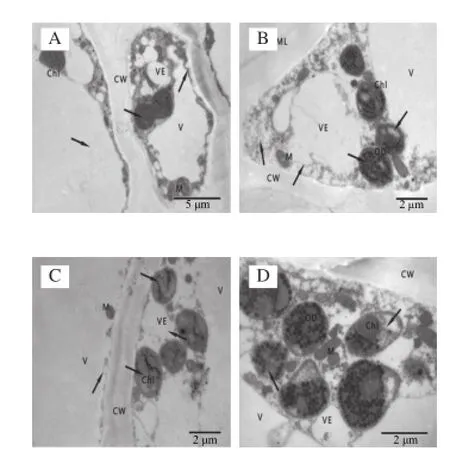
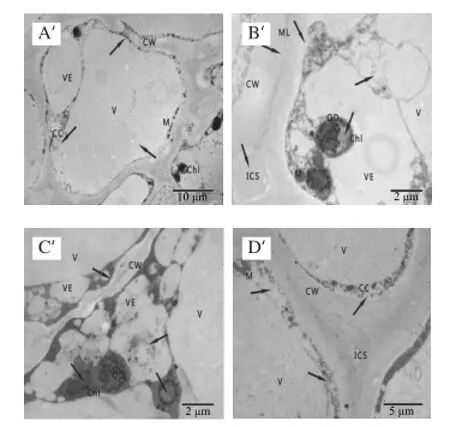
圖3 冷藏第28天杏果實超微結構Fig.3 Ultrastructure of apricot fruits after 28 days of cold storage
杏果實在冷藏第28天細胞超微結構發生明顯變化。SA處理的杏果實細胞排列整齊,細胞壁厚度均勻,細胞壁中膠層略有降解,細胞內囊泡增多并膨脹;大多數葉綠體開始趨于圓形,基粒片層和基質片層結構模糊,出現空泡現象,葉綠體雙層膜結構模糊或消失,嗜鋨顆粒數目增多;線粒體數量較多,個別線粒體內出現小泡,內脊結構模糊,數量減少,液泡結構完整。而對照杏果實細胞間隙較大,細胞壁彎曲變形,細胞質膜皺縮,細胞質明顯減少,細胞器排列混亂,囊泡化較嚴重;葉綠體雙層膜結構破壞,有明顯的空泡化現象,基質片層和基粒片層結構消失;線粒體結構模糊且數量減少,內部呈小泡化;液泡膜折皺,胞間層開始分解,形成絮狀空隙。由此可知,采后冷藏28 d對照杏果實細胞已經出現冷害跡象,各細胞器結構明顯破壞,而SA處理的杏果實仍能較好維持細胞結構的完整。
2.4 杏果實冷藏第49天透射電鏡觀察
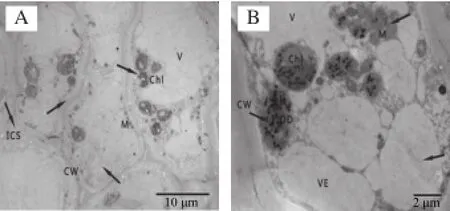
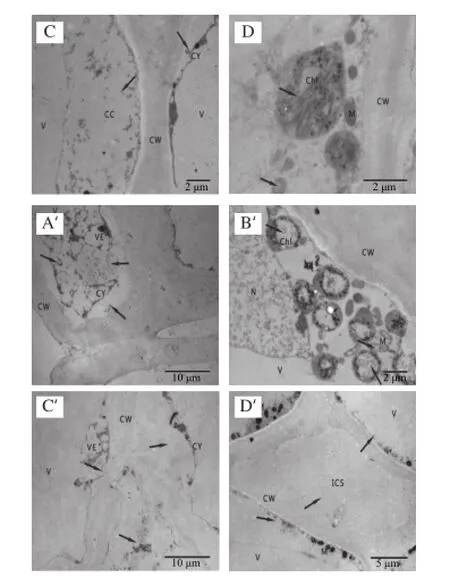
圖4 冷藏第49天杏果實超微結構Fig.4 Ultrastructure of apricot fruits after 49 days of cold storage
到冷藏后期,SA處理組和對照組杏果實細胞超微結構均發生不同程度的變化。對照組杏果實細胞已嚴重破壞,細胞壁嚴重彎曲變形并呈絮狀,有的已經解體;細胞內容物減少,質壁分離嚴重,囊泡明顯增多,聚集在細胞壁一側,中膠層降解消失;細胞間隙進一步增大,大量細胞器降解,液泡及細胞器內含物與原生質互溶,葉綠體和線粒體出現嚴重空腔現象,雙層膜結構消失。SA處理的杏果實細胞也已經變形,細胞壁彎曲,細胞質明顯減少,且出現質壁分離現象,細胞質膜折皺,細胞內區域化破壞,渾濁不清,細胞器排列紊亂,胞內出現絮狀沉淀物質堆積。細胞內囊泡膨脹,葉綠體、線粒體等細胞器仍清晰可見,個別已經變形、解體,細胞器雙層膜模糊或消失。但與對照杏果實相比細胞損傷程度較輕。
3 結 論
細胞是生物體最基本的結構和功能單位。超微結構的變化是果實發育成熟及衰老過程的重要特征[16-17],也可以反映果實的生理狀態。在低溫貯藏條件下,能引起植物生理代謝變化的同時,組織細胞的超微結構也會發生一系列相應變化,這些變化會導致組織細胞功能衰弱甚至喪失,進一步加速了細胞衰老與解體[18-19]。簡令成等[20-21]指出冷害會引起植物組織各類細胞器結構的變化,其破壞程度取決于植物品種的抗寒性。
電鏡觀察結果表明,剛采收及冷藏前期的杏果實細胞壁厚度均勻,中膠層明顯;葉綠體、線粒體、液泡、細胞核等細胞器結構完整;細胞質膜完好,沒有破裂現象的發生。冷藏的第28天,對照杏果實中膠層逐漸降解消失,細胞壁輕微變形,質壁分離,葉綠體趨于圓形并開始膨大,嗜鋨顆粒數目增多,體積變大。隨著貯藏時間的延長,杏果實細胞及其超微結構嚴重破壞,細胞發生質壁分離,細胞壁彎曲變形甚至解體,細胞膜結構遭到破壞,葉綠體、線粒體等細胞器發生變形或解體,液泡及細胞器內含物與原生質互溶,細胞空腔化,這與牛先前等[22]對枇杷和鄭麗梅等[23]對黃瓜幼苗的研究結果相似。與對照杏果實相比,SA處理杏果實能較好的維持細胞超微結構的完整性,顯著減輕杏果實細胞的損傷程度。與電鏡觀察的微觀結果對應,在宏觀表現上,本實驗結果表明,貯藏28 d后,對照杏果實果面呈現大面積水浸斑,硬度增加,出汁率下降,果肉褐變等明顯的冷害癥狀伴[24];而SA處理果實推遲7 d發生冷害,且冷害癥狀較對照果實輕,這與在冷藏的中期和后期的電鏡觀察結果相吻合。
植物冷害的自由基傷害學說指出,冷敏植物遭受冷脅迫后,活性氧自由基大量積累,導致活性氧代謝平衡失調,就會引起膜脂的不飽和鍵發生過氧化作用,細胞膜系統受到傷害是早期的冷害癥狀之一,最終加劇果實冷害的發生[25]。侯媛媛等[26]的研究表明,SA處理可減輕杏果實的冷害程度與SA能防止杏果實冷藏期間過高的氧化傷害,維持杏果實活性氧代謝平衡密切相關。本研究認為,SA處理能有效減輕杏果實冷藏期間質膜的傷害程度,與SA能防止杏果實冷藏期間過高的氧化傷害,維持杏果實活性氧代謝平衡是密切相關的。
綜上所述,杏果實冷藏期間細胞壁及各類細胞器的破壞可能是果實發生冷害的微觀表現。電鏡觀察表明,SA處理能較好的維持杏果實細胞壁、葉綠體、線粒體、液泡等細胞結構的完整性,這與SA能減輕杏果實冷藏期間冷害的發生、延長杏果實貯藏時間密切相關。
[1] 新疆維吾爾自治區統計局. 新疆統計年鑒[M]. 北京: 中國統計出版社, 2012: 338.
[2] 劉路, 張謙, 趙曉梅. 不同處理對賽買提杏貯藏保鮮效果的影響[J].食品工業, 2010(2): 32-34.
[3] 胡花麗, 李鵬霞, 梁麗松, 等. 不同貯藏溫度及成熟度對杏果實品質的影響[J]. 食品與發酵工業, 2009, 35(1): 182-185.
[4] 胡芳, 馬書尚, 張繼澍, 等. 1-甲基環丙烯對“富有”甜柿采后主要生理指標及細胞超微結構的影響[J]. 園藝學報, 2009, 36(4): 487-492.
[5] 孫芳娟, 韓明玉, 趙彩萍, 等. 不同采收成熟度油桃貯藏效果及果肉細胞超微結構觀察[J]. 果樹學報, 2009, 26(4): 450-455.
[6] 陸勝民, 席玙芳. 采后處理對梅果細胞超微結構的影響[J]. 食品科學, 2001, 22(6): 62-65.
[7] 謝娟. 紅樹莓果實生長發育規律及超微結構研究[D]. 哈爾濱: 東北農業大學, 2011.
[8] 王志坤, 秦智偉, 李艷秋, 等. 黃瓜果實衰老過程中果皮超微結構的變化[J]. 園藝學報, 2007, 34(4): 889-894.
[9] 李銀, 張輝, 駱建敏, 等. 蟠桃果實發育成熟過程中果肉細胞超微結構的變化[J]. 新疆農業科學, 2011, 48(6): 1006-1010.
[10] 鄭國華, 張賀英. 不同低溫脅迫下早鐘6號枇杷幼果細胞超微結構的變化[J]. 福建農林大學學報: 自然科學版, 2008, 37(5): 473-476.
[11] 段玉權, 馮雙慶, 趙玉梅, 等. 1-甲基環丙烯處理對冷藏桃果肉細胞超微結構的影響[J]. 中國農業科學, 2004, 37(12): 2039-2042.
[12] MOHAMMADREZA A, MORTEZA S. Impact of salicylic acid on post harvest physiology of horticultural crops[J]. Trends in Food Science and Technology, 2010(21): 502-509.
[13] AGHDAM S M, ASGHARI R M, MORADBEyGI H, et al. Effect of postharvest salicylic acid treatment on reducing chilling injury in tomato fruit[J]. Romanian Biotechnological Letters, 2012, 17(4): 7466-7473.
[14] CAO Shifeng, HU Zhichao, ZHENG yonghua, et al. Synergistic effect of heat treatment and salicylic acid on alleviating internal browning in cold-stored peach fruit[J]. Postharvest Biology and Technology, 2010, 58(2): 93-97.
[15] SAyyARI M, BABALAR M, KALANTARI S, et al. Effect of salicylic acid treatment on reducing chilling injury in stored pomegranates[J]. Postharvest Biology and Technology, 2009, 53(3): 152-154.
[16] 任亞梅, 劉興華, 徐春雅, 等. 不同處理對獼猴桃采后生理和細胞超微結構的影響[J]. 農業工程學報, 2008, 24(7): 217-221.
[17] 張大鵬, 李氓, 王毅. 葡萄果實發育過程中果肉細胞超微結構的觀察[J]. 植物學報, 1997, 39(5): 389-396.
[18] 杜艷, 喻方圓, 甘習華, 等. 低溫脅迫下兩種七葉樹苗木超微結構的比較[J]. 南京林業大學學報, 2007, 31(3): 111-114.
[19] POMEROy M K, ANDREWS C J. Ultrastructural changes in shoot apex cells of winter wheat seedlings during ice encasement[J]. Canadian Journal of Botany, 1978, 56: 786-794.
[20] 簡令成. 植物抗寒機理研究的新進展[J]. 植物學通報, 1992, 9(3): 17-22.
[21] 簡令成, 李積宏, 李本湘, 等. 楊樹頂芽細胞內質網與其他膜系統的結構聯系及其在休眠過程中的變化[J]. 植物學報, 2000, 42(8): 803-810.
[22] 牛先前, 鄭國華, 林秀香, 等. 冰核細菌對低溫脅迫下枇杷幼果中果肉超微結構的影響[J]. 中國生態農業學報, 2011, 19(2): 388-393.
[23] 鄭麗梅, 司龍亭, 韓貴超. 低溫處理對不同耐寒性黃瓜幼苗葉片超微結構的影響[J]. 西北農業學報, 2009, 18(4): 276-279.
[24] 袁潔, 朱璇, 逄煥明, 等. 外源水楊酸處理對采后杏果實抗冷性的影響[J]. 食品工業科技, 2013, 34(22): 349-351.
[25] MITTLER R. Oxidative stress, antioxidants and stress tolerance[J]. Trends in Plant Science, 2002, 7(9): 405-410.
[26] 侯媛媛, 朱璇, 王英, 等. 水楊酸處理對杏果實冷害及活性氧代謝的影響[J]. 食品科學, 2014, 35(4): 1-5.
Effect of Salicylic Acid Treatment on Cell Ultrastructure of Apricot Fruit during Cold Storage
ZHU Xuan1,2, HOU Yuan-yuan1, JIA Yan3, ZHAO Ya-ting1, MA Xuan1
(1. College of Food Science and Pharmacy, Xinjiang Agricultural University, ürümqi 830052, China; 2. Xinjiang Engineering Technology Research Center of Fruit Processing and Preservation, ürümqi 830052, China; 3. College of Food Engineering and Biotechnology, Tianjin University of Science and Technology, Tianjin 300457, China)
Saimaiti apricot fruits from Xinjiang, China were treated with 0.01 g/L salicylic acid (SA) and then stored at 0 ℃to explore the change of cell ultrastructure during different cold storage periods, and distilled water was used as control treatment. The results showed that the cell ultrastructure of apricot fruits from the salicylic acid treatment and control groups had no significant difference in neat rows or the integrity of cell wall structure and cell organelles based on cell arrangement. The ultrastructure of apricot fruits changed obviously after the 28thday, which suggested that cell organelles was deformed or dissolved. Meanwhile, enlarged intercellular gap junctions, decreased cell contents, and empty cell cavity were observed. Compared with the control, the improvement of cold resistance in apricot fruits by SA treatment was closely related to the alleviated damage of cell wall, chloroplast and mitochondria, and the integrality maintenance of cell organelles and membrane system.
salicylic acid; apricot fruit; cold storage; ultrastructure
TS255.3
A
1002-6630(2014)14-0193-05
10.7506/spkx1002-6630-201414038
2014-01-03
國家自然科學基金地區科學基金項目(31260408);公益性行業(農業)科研專項(201303075)
朱璇(1971—),女,副教授,博士,研究方向為果蔬貯藏與保鮮。E-mail:13999877961@126.com
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
