溫度和光徑對(duì)微綠球藻生長(zhǎng)及營(yíng)養(yǎng)成分含量的影響
2014-03-07 07:12:58杜曉鳳孫東紅
海洋科學(xué) 2014年4期
杜曉鳳, 鄒 寧, 孫東紅, 常 林, 趙 萍
(魯東大學(xué) 生命科學(xué)學(xué)院, 山東 煙臺(tái) 264025)
溫度和光徑對(duì)微綠球藻生長(zhǎng)及營(yíng)養(yǎng)成分含量的影響
杜曉鳳, 鄒 寧, 孫東紅, 常 林, 趙 萍
(魯東大學(xué) 生命科學(xué)學(xué)院, 山東 煙臺(tái) 264025)
研究了溫度和光生物反應(yīng)器的光徑對(duì)微綠球藻(Nannochloropsisoculata Hibberd)生長(zhǎng)及細(xì)胞內(nèi)幾種營(yíng)養(yǎng)成分含量的影響。結(jié)果表明, 溫度對(duì)微綠球藻的生長(zhǎng)有顯著影響(P<0.05), 對(duì)幾種營(yíng)養(yǎng)成分的含量有極顯著影響(P<0.01), 在 28℃的培養(yǎng)條件下, 細(xì)胞生長(zhǎng)速度最快, 此時(shí)蛋白質(zhì)含量最高, 為5.66%, 多糖含量最低, 僅為1.47%, 但最大生物產(chǎn)量卻是在30℃的條件下獲得, 總脂與葉綠素a的含量隨溫度的升高而降低, 類(lèi)胡蘿卜素含量隨溫度升高而增大。光生物反應(yīng)器的光徑對(duì)微綠球藻的生長(zhǎng)和總脂、可溶性蛋白、色素的含量有極顯著影響(P<0.01), 對(duì)多糖含量有顯著影響(P<0.05), 比生長(zhǎng)速率、最高細(xì)胞密度和單位體積產(chǎn)量均隨光徑的增大而減小, 單位面積產(chǎn)量隨光徑的增大而增大; 總脂、可溶性蛋白和多糖的含量隨光徑的增大而降低, 葉綠素和類(lèi)胡蘿卜素含量隨光徑增大而增大, 類(lèi)胡蘿卜素含量的增大趨勢(shì)比葉綠素更明顯。
微綠球藻(Nannochloropsisoculata Hibberd); 溫度; 光徑; 蛋白質(zhì); 多糖; 總脂; 色素
微綠球藻是一種海洋單胞藻, 呈球形, 其細(xì)胞壁極薄, 易于消化吸收, 加上其易培養(yǎng)、繁殖迅速、營(yíng)養(yǎng)豐富等特點(diǎn), 是泥蚶、蟹、蝦等幼苗及輪蟲(chóng)的優(yōu)良餌料, 在水產(chǎn)養(yǎng)殖中應(yīng)用較為廣泛。微綠球藻細(xì)胞含有極豐富的二十碳五烯酸(EPA)、蛋白質(zhì)、多糖等物質(zhì), 是健康食品和藥品的重要資源之一[1]。
目前國(guó)內(nèi)外對(duì)微綠球藻的研究多集中在藻的培養(yǎng)工藝[2-3]、分子系統(tǒng)學(xué)[4]及環(huán)境因子對(duì)脂肪酸組成及含量[5-7]的影響等方面, 對(duì)其培養(yǎng)條件的研究多集中在光照[8]、溫度[6,9-10]、氮源及氮濃度[11-14]、pH[15]等方面。本研究所用反應(yīng)器為自行研制的氣升式平板光生物反應(yīng)器, 與傳統(tǒng)反應(yīng)器相比, 能有效防止污染, 實(shí)現(xiàn)高密度純種培養(yǎng), 還可以增加光照面積, 提高光能利用率。作者旨在研究溫度和反應(yīng)器的光徑對(duì)微綠球藻生長(zhǎng)狀況的影響, 并分析了各營(yíng)養(yǎng)成分的含量, 為今后平板光生物反應(yīng)器的應(yīng)用及微綠球藻的培養(yǎng)研究提供參考。
1 材料與方法
1.1 實(shí)驗(yàn)材料
微綠球藻(Nannochloropsis oculataHibberd)藻種由魯東大學(xué)生命科學(xué)學(xué)院藻類(lèi)研究所提供。
1.2 培養(yǎng)方法及培養(yǎng)條件
采用氣升式平板光生物反應(yīng)器, 反應(yīng)器底部安裝由玻璃條固定的多孔通氣管, 向反應(yīng)器內(nèi)通入空氣。將反應(yīng)器和通氣管洗凈, 加滿(mǎn)自來(lái)水, 然后按 1 mL/L的量加入次氯酸, 滅菌24 h后, 將自來(lái)水倒出, 加入滅菌的培養(yǎng)基和海水, 培養(yǎng)所用海水取自自然海水,過(guò)濾滅菌后使用, 培養(yǎng)基配方參見(jiàn)鄒寧等[16]。同時(shí)接入相同的藻種, 控制接入后的藻種密度相同或接近,同時(shí)在自然光、室溫下進(jìn)行培養(yǎng), 培養(yǎng)基的初始 pH為8.0。
1.3 實(shí)驗(yàn)方法
1.3.1 實(shí)驗(yàn)梯度設(shè)置
溫度對(duì)微綠球藻影響: 選用光徑為 5cm的反應(yīng)器, 用碘鎢燈做光源, 控制光照強(qiáng)度約為 5000 lx,光周期L: D為10:14, 在碘鎢燈與反應(yīng)器之間放置加滿(mǎn)水的空白反應(yīng)器降溫, 控制溫度梯度為: 20、25、28、30、32℃, 每組設(shè)置3個(gè)平行。
光徑對(duì)微綠球藻的影響: 選取光徑分別為 5、10、15、20cm且長(zhǎng)度和高度均相同的4組反應(yīng)器, 每組3個(gè)平行, 培養(yǎng)過(guò)程測(cè)得光照強(qiáng)度為: 1300~31000 lx,白天溫度為23~30℃。
1.3.2 生長(zhǎng)測(cè)定方法
每天上午9: 00~10: 00取樣, 用紫外分光光度計(jì)測(cè)定藻液在波長(zhǎng)660 nm處的吸光值, 用血球計(jì)數(shù)板計(jì)數(shù)細(xì)胞密度。生長(zhǎng)計(jì)算公式為: 比生長(zhǎng)速率μ= (lnN2?lnN1)/(t2–t1)[17], 其中其中N2為試驗(yàn)剛進(jìn)入穩(wěn)定期的細(xì)胞數(shù)目,N1為試驗(yàn)剛進(jìn)入對(duì)數(shù)期的細(xì)胞數(shù)目,t2為試驗(yàn)剛進(jìn)入穩(wěn)定期的時(shí)間,t1為試驗(yàn)剛進(jìn)入對(duì)數(shù)期的時(shí)間。
1.3.3 微藻的采收
采收處于穩(wěn)定期的藻液, 用大型離心機(jī)離心, 3 000 r/min離心30 min, 藻泥用去離子水洗2~3次,一部分制成藻泥于冰箱中冷藏保存, 用于測(cè)定色素含量; 一部分于電熱鼓風(fēng)干燥箱中 65℃烘干至恒重,將烘干的藻粉置于干燥皿中待用。
1.3.4 總脂含量的測(cè)定
準(zhǔn)確稱(chēng)取 0.15g±0.01g藻粉于具塞試管中, 加入 1 mL蒸餾水, 在混勻器上混勻后, 置于–20℃冰箱中反復(fù)凍融3次。加入5mL工業(yè)酒精, 混勻后在室溫下提取 3 h, 將上清液移入離心管中, 加入適量高效活性白土, 于搖床上充分搖勻脫色, 6000 r/min離心5 min, 收集上清于恒重的稱(chēng)量瓶中, 殘?jiān)祷卦嚬軆?nèi)重復(fù)提取2次, 合并上清, 將上清于旋轉(zhuǎn)蒸發(fā)儀中去除提取劑, 稱(chēng)質(zhì)量, 計(jì)算油脂含量。
油脂含量=(油脂質(zhì)量/藻粉質(zhì)量)×100%
1.3.5 可溶性蛋白含量的測(cè)定
準(zhǔn)確稱(chēng)取 0.15g±0.01g藻粉于 5mL離心管中,加入 2 mL蒸餾水, 在混勻器上混勻后, 置于?20℃冰箱中反復(fù)凍融3次, 再加入2 mL蒸餾水, 離心收集上清, 殘?jiān)舆m量蒸餾水重復(fù)離心, 合并上清液,定容到50 mL, 每組采用3個(gè)平行。采用考馬斯亮藍(lán)法測(cè)定蛋白含量[18]。
1.3.6 可溶性多糖含量的測(cè)定
準(zhǔn)確稱(chēng)取0.15 g±0.01 g藻粉于5 mL離心管中,加入 2 mL蒸餾水, 在快速混勻器上混勻后, 置于–20℃冰箱中反復(fù)凍融3次, 再加入2 mL蒸餾水, 離心收集上清; 殘?jiān)舆m量蒸餾水, 于沸水浴中加熱3 h, 離心收集上清。合并上清液并定容到50mL, 每組采用3個(gè)平行。采用蒽酮法測(cè)定多糖的含量[19]。
1.3.7 色素含量的測(cè)定
準(zhǔn)確稱(chēng)取0.15 g±0.01 g藻粉于具塞試管中, 加入 1 mL蒸餾水, 在混勻器上混勻后, 置于?20℃冰箱中反復(fù)凍融3次。加入5 mL80%的丙酮, 混勻后置于4℃冰箱內(nèi)提取3 h以上, 上清倒入5 mL離心管中離心, 取上清; 殘?jiān)谷朐嚬苤? 再加 5 mL 80%丙酮提取3h以上, 離心合并上清, 于25 mL容量瓶中定容。用Arnon法測(cè)定色素含量[20]。
1.3.8 數(shù)據(jù)處理
實(shí)驗(yàn)數(shù)據(jù)采用 SPSS16.0軟件進(jìn)行分析, 以P<0.05為顯著性差異, 以P<0.01為極顯著差異。
2 結(jié)果與分析
2.1 溫度對(duì)微綠球藻生長(zhǎng)及營(yíng)養(yǎng)成分含量的影響
2.1.1 溫度對(duì)微綠球藻生長(zhǎng)的影響
溫度對(duì)微綠球藻的生長(zhǎng)有顯著影響(P<0.05),如表1所示, 在適宜的溫度條件下, 微綠球藻生長(zhǎng)迅速, 30℃實(shí)驗(yàn)組的比生長(zhǎng)速率最高, 其次為28℃和32℃實(shí)驗(yàn)組, 3組間差異不顯著; 最高細(xì)胞密度隨溫度的升高先升高后降低, 30℃實(shí)驗(yàn)組最高, 32℃實(shí)驗(yàn)組次之, 2組間差異不顯著。單位面積產(chǎn)量和單位體積產(chǎn)量都隨溫度的升高先升高后降低,在 30℃時(shí)達(dá)到最高, 最高值分別為 1.988 g/(m2·d)和39.598 mg/(m2·d), 這與比生長(zhǎng)速率μ和最高細(xì)胞密度的變化趨勢(shì)一致。可粗略得出微綠球藻的最適生長(zhǎng)溫度為28~32℃。
2.1.2 溫度對(duì)微綠球藻營(yíng)養(yǎng)成分含量的影響

表1 溫度對(duì)微綠球藻生長(zhǎng)的影響Tab.1 Effect of temperature on the growth rate of N. oculata
溫度是影響微藻脂肪含量的重要因子之一, 溫度降低時(shí)藻細(xì)胞內(nèi)不飽和脂肪酸含量增加, 飽和脂肪酸含量減小[10,21]。溫度對(duì)總脂含量有極顯著影響(P<0.01), 由表 2可以看出, 在 20~32℃的溫度范圍內(nèi), 隨著溫度的升高, 微綠球藻細(xì)胞內(nèi)的總脂含量呈下降趨勢(shì)。James等[22]報(bào)道微綠球藻在35℃下生長(zhǎng)時(shí), EPA的含量比在25℃和30℃時(shí)減少4%; 溫度對(duì)可溶性蛋白和多糖含量有極顯著影響(P<0.01), 隨著溫度的升高, 可溶性蛋白含量先升高后降低, 在 28℃條件下達(dá)到最高值, 與此相反, 多糖含量先下降后升高, 在 28℃條件下達(dá)到最低。可能是由于在適宜的溫度條件下, 細(xì)胞生長(zhǎng)迅速, 合成了大量的蛋白, 此時(shí)消耗大量的能量, 造成多糖含量減小; 溫度對(duì)微綠球藻葉綠素和類(lèi)胡蘿卜素的含量有極顯著影響(P<0.01), 葉綠素含量隨著溫度的升高而減小, 類(lèi)胡蘿卜素的含量隨著溫度的升高而增大。
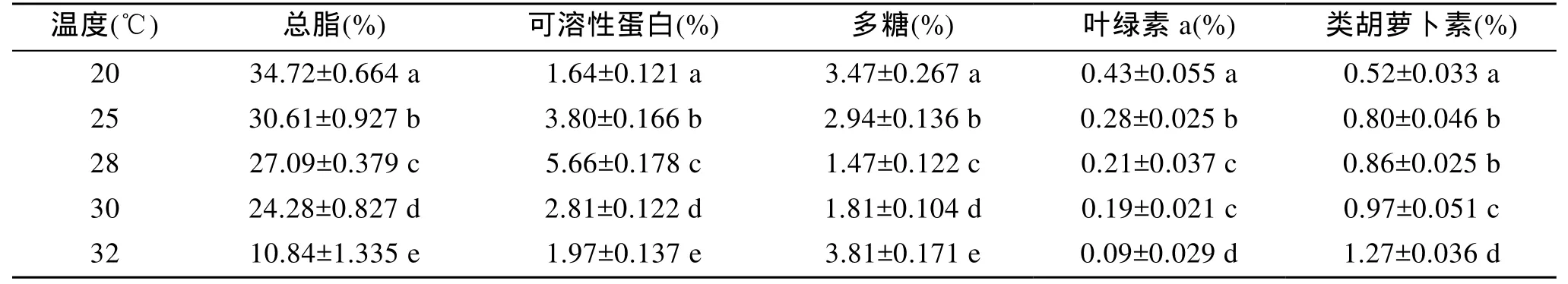
表2 溫度對(duì)微綠球藻可溶性蛋白、多糖、色素含量的影響Tab.2 Effect of temperature on the lipid, soluble protein, polysaccharide and pigments contents of N. oculata
2.2 反應(yīng)器的光徑對(duì)微綠球藻生長(zhǎng)及營(yíng)養(yǎng)成分含量的影響
2.2.1 光徑對(duì)微綠球藻生長(zhǎng)的影響
采用光生物反應(yīng)器對(duì)微綠球藻進(jìn)行高密度培養(yǎng),通過(guò)實(shí)驗(yàn)得出, 反應(yīng)器的光徑對(duì)微綠球藻生長(zhǎng)的影響如表3所示。
通過(guò)方差分析和多重比較可知, 反應(yīng)器的光徑對(duì)微綠球藻生長(zhǎng)的影響極顯著(P<0.01)。微綠球藻在5~20 cm的光徑范圍內(nèi)均能正常生長(zhǎng), 細(xì)胞密度與光徑成反比。光徑越大, 反應(yīng)器內(nèi)部的細(xì)胞接受到的有效光源越少, 不能充分的進(jìn)行光合作用, 光合產(chǎn)物少, 細(xì)胞生長(zhǎng)慢。
如表 1所示, 比生長(zhǎng)速率與最高細(xì)胞密度均與光徑成反比。反應(yīng)器的光徑越小, 細(xì)胞的生長(zhǎng)速率越高[23]。隨著光徑的增大, 單位面積產(chǎn)量幾乎呈直線上升, 從5 cm的1.931 g/(m2·d)上升到20 cm的3.354 g/ (m2·d)。與此相反, 單位體積產(chǎn)量隨著光徑的增大而降低, 從 5 cm的 38.611 mg/(L·d)下降到 20 cm的17.272 mg/(L·d), 單位體積產(chǎn)量降了2倍多。
2.2.2 光徑對(duì)微綠球藻細(xì)胞中營(yíng)養(yǎng)成分含量的影響
光徑對(duì)油脂積累有極顯著影響(P< 0.01), 由表4可以看出, 微綠球藻細(xì)胞內(nèi)總脂含量與光徑成反比, 5 cm光徑實(shí)驗(yàn)組的總脂含量顯著高于其他組。在短光徑平板反應(yīng)器中培養(yǎng)微綠球藻, 可以大幅提高EPA 的含量[24]。光徑對(duì)可溶性蛋白含量有極顯著影響(P<0.01), 對(duì)多糖含量有顯著影響(P< 0.05), 可溶性蛋白和多糖含量均隨著光徑的增大而減小, 可知短光徑的光生物反應(yīng)器, 不僅有利于增大細(xì)胞密度,還有利于有機(jī)物物質(zhì)的積累。光徑對(duì)微綠球藻葉綠素和類(lèi)胡蘿卜素的含量有極顯著影響(P<0.01), 葉綠素和類(lèi)胡蘿卜素的含量均隨著光徑的增大而增大。在5 cm和10 cm光徑條件下, 類(lèi)胡蘿卜素的含量高于葉綠素的含量, 且光徑越小, 高出的比例越大, 這與Zou[25]的研究結(jié)果一致; 在15 cm和20 cm的光徑條件下, 葉綠素含量高于類(lèi)胡蘿卜素的含量, 且光徑越大, 高出的比例越大。
3 結(jié)論

表3 光徑對(duì)微綠球藻生長(zhǎng)的影響Tab.3 Effect of optical paths on the growth rate of N. oculata
溫度對(duì)微綠球藻的生長(zhǎng)有顯著影響, 反應(yīng)器的光徑有極顯著影響。微綠球藻只有在適宜的溫度條件下才能生長(zhǎng)旺盛和積累有機(jī)物。而光徑影響細(xì)胞接受到的有效光源, 靠近光生物反應(yīng)器表面區(qū)域內(nèi)的藻細(xì)胞才能接受到有效的光源, 光徑小的反應(yīng)器,表面積體積比大, 光暗循環(huán)周期短, 光合作用效率高, 積累的蛋白質(zhì)、多糖和總脂的含量高, 但較強(qiáng)光照對(duì)色素有一定破壞作用, 所以色素含量較低; 在相同的表面積下, 藻細(xì)胞數(shù)目與反應(yīng)器的光徑成正比, 所以單位面積產(chǎn)量隨著光徑的增大而增高[26-27]。
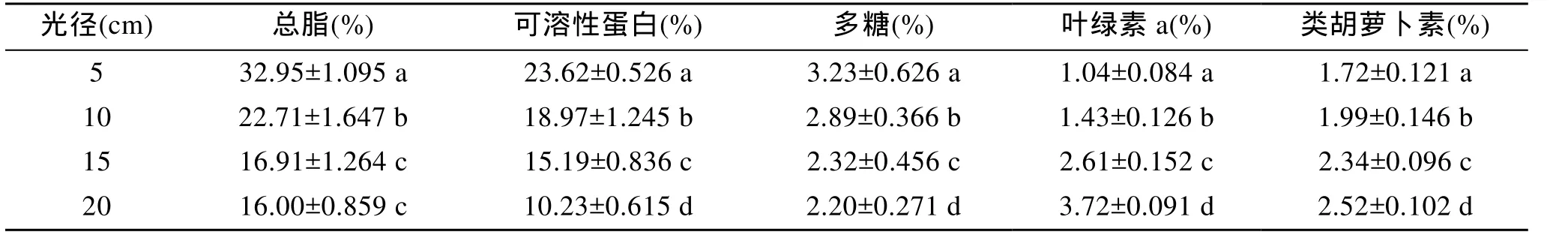
表4 子光徑對(duì)微綠球藻總脂、可溶性蛋白、多糖、色素含量的影響Tab.4 Effect of optical paths on the lipid, soluble protein, polysaccharide and pigments contents of N. oculata
[1] 趙大顯, 周志剛. 微綠球藻DNA質(zhì)粒文庫(kù)的構(gòu)建[J].海洋科學(xué), 2006, 30(5): 87-91.
[2] Xu F, Hu H H, Cong W, et al. Growth characteristics and eicosapentaenoic acid production byNannochloropsissp. in mixotrophic conditions [J]. Biotechnology Letters, 2004, 26(1): 51-53.
[3] Liliana R, Graziella C Z, Laura B, et al. Growth medium recycling inNannochloropsissp. mass cultivation[J]. Biomolecular Engineering, 2003, 20(4-6): 243-248.
[4] Shoichiro S, Mika A, Hideaki M. Taxonomic characterization of a marine Nannochloropsis species,N. oceanicsp. nov. (Eustigmatophyceae) [J]. Phycologia, 2002, 41(3): 273-279.
[5] 蔣霞敏, 鄭亦周. 14種微藻總脂含量和脂肪酸組成研究[J]. 水生生物學(xué)報(bào), 2003, 27(3): 243-247.
[6] 周洪琪, 易翠平, 丁卓平, 等. 環(huán)境因子對(duì)青島大扁藻、亞心形扁藻、微綠球藻脂肪酸組成的影響[J]. 浙江海洋學(xué)院學(xué)報(bào)(自然科學(xué)版), 2001, 20(Supplement): 112-117.
[7] 魏東, 張學(xué)成, 鄒立紅, 等. 細(xì)胞生長(zhǎng)時(shí)期對(duì)兩種海洋微藻總脂含量和脂肪酸組成的影響[J]. 青島海洋大學(xué)學(xué)報(bào), 2000, 30(3): 503-509.
[8] 杜曉鳳, 鄒寧, 孫東紅, 等. 光照強(qiáng)度對(duì)微綠球藻生長(zhǎng)及有機(jī)質(zhì)積累的影響[J]. 生物過(guò)程, 2011, 1(2): 18-21.
[9] 張海琪, 蔣霞敏, 潘雙葉, 等. 光照、溫度、碳源及接種密度對(duì)微綠球藻生長(zhǎng)的影響[J]. 河北漁業(yè), 2001, (1): 9-13.
[10] Susan M R, Luong-Van T, George L, et al. Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures[J]. Aquaculture, 2002, 211(1-4): 195-214.
[11] 魏東, 張學(xué)成, 隋正紅, 等. 氮源和 N/P對(duì)眼點(diǎn)擬微球藻的生長(zhǎng)、總脂含量和脂肪酸組成的影響[J]. 海洋科學(xué), 2000, 24(7): 46-51.
[12] 潘庭雙, 胡賢江, 侯冠軍, 等. 氮對(duì)微綠球藻生長(zhǎng)的影響[J]. 安徽農(nóng)業(yè)科學(xué), 2001, 29(4): 548-555.
[13] 蔣霞敏. 溫度、光照、氮含量對(duì)微綠球藻生長(zhǎng)及脂肪酸組成的影響[J]. 海洋科學(xué), 2002, 26(8): 9-13.
[14] 黃旭雄, 周洪琪, 袁燦東, 等. 氮源及濃度對(duì)微綠球藻營(yíng)養(yǎng)價(jià)值的影響[J]. 上海水產(chǎn)大學(xué)學(xué)報(bào), 2003, 12(2): 113-116.
[15] 王秀良, 劉晨臨, 張學(xué)成. pH對(duì)眼點(diǎn)擬微綠球藻的生長(zhǎng)、總脂含量及脂肪酸組成的影響[J]. 海洋科學(xué), 2002, 26(5): 63-67.
[16] 鄒寧, 孫東紅, 韓亞香. CO2對(duì)牟氏角毛藻高密度培養(yǎng)的影響[J]. 生物工程學(xué)報(bào), 2005, 21(5): 844-847.
[17] S. John P. Principles of microbes and cell cultivation[M]. New York: Blackwell Scientific Publications, 1975: 260-268.
[18] 叢峰松. 生物化學(xué)實(shí)驗(yàn)[M]. 上海: 上海交通大學(xué)出版社, 2005.
[19] 張志良. 植物生理學(xué)實(shí)驗(yàn)指導(dǎo)(第二版)[M]. 北京:高等教育出版社, 2001: 160-162.
[20] 陳建勛, 王曉峰. 植物生理學(xué)實(shí)驗(yàn)指導(dǎo)(第二版)[M].廣州: 華南理工大學(xué)出版社, 2006: 81-84.
[21] Daniel V L, Guy A T. Low temperature-Induced alterations in the chloroplast and microsomal membranes of dunaliella salina[J]. Plant Physiology, 1982, 69(6): 13.
[22] James C M, Al-Hinty S, Salman A E. Growth and ω3 fatty acid and amino acid composition of microalgae under different temperature regimes[J]. Aquaculture, 1989, 77(4): 337-351.
[23] Amos R, Zou N. Efficient utilisation of high photon irradiance for mass production of photoautotrophic micro-organisms[J]. Journal of Applied Phycology, 1999, 11(1): 123-127.
[24] Zou N, Zhang C W, Zvi Cohen et al. Production of cell mass and eicosapentaenoic acid (EPA) in ultrahigh cell density cultures ofNannochloropsissp. (Eustigmatophyceae)[J]. European Journal of Phycology, 2000, 35(2): 127-133.
[25] Zou N, Amos R. Light-path length and population density in photoacclimation ofNannochloropsissp. (Eustigmatophyceae) [J]. Journal of Applied Phycology, 2000, 12(3-5): 349-354.
[26] Zou N, Amos R. Effect of light-path length in outdoor flat plate reactors on output rate of cell mass and of EPA inNannochloropsissp.[J]. Journal of Biotechnology, 1999, 70(1-3): 351-356.
[27] Qiang H, Yair Z, Amos R. Combined effects of light intensity, light-path and culture density on output rate ofSpirulina platensis(Cyanobacteria)[J]. European Journal of Phycology 1998, 33(2): 165-171.
(本文編輯: 梁德海)
Effect of temperature and optical path on growth rate and accumulation of nutrients ofNannochloropsissp.
DU Xiao-feng, ZOU Ning, SUN Dong-hong, CHANG Lin, ZHAO Ping
(College of Life Sciences, Ludong University, Yantai 264025, China)
Jun., 12, 2013
Nannochloropsissp.; temperature; optical path; protein; polysaccharide; lipid; pigments
The effect of temperature and optical path of flat plate glass reactors on growth rate and organics contents ofNannochloropsissp. was investigated, using a range of optical paths from 5 to 20 cm. The results showed that the temperature had a significant effect on the growth ofNannochloropsis(P<0.05), and a very significant effect on the contents of mutrients (P<0.01). The cell growth had a highest speed at 28℃, when the content of soluble protein reached the maximum and that of polysaccharides reached the minimum. The biomass yield reached the maximum at 30℃. The contents of lipid and chlorophyll decreased with the increase of temperature, while the content of carotenoid increased with the increase of temperature. The optical path of photobioreactors had a very significant effect on the growth ofNannochloropsis(P<0.01). Specific growth rate, maximal cell density (107cell/mL) and yield per unit volume (mg /(L·d) ) decreased with increase in optical path. Yield per unit area (g /(m2·d) ) increased with increased optical path. The contents of lipid, soluble protein, and polysaccharides decreased with increased optical path, but the contents of chlorophyll and carotenoid increased with increased optical path. The increase trend of carotenoids was more obvious than that of chlorophyll. In summary, the optimal optical path determines the optimal culture density, as well as yield per unit volume and the productivity of organics. Under our conditions the optimal optical path for culturingNannochloropsisto accumulate lipid, protein and polysaccharide in plate reactors was 5 cm, the optimal optical path for accumulating pigments was 20 cm.
Q949.2
A
1000-3096(2014)04-0050-05
10.11759/hykx20120419002
2013-06-12;
2014-02-18
杜曉鳳(1985-), 女, 山東臨沂人, 碩士研究生, 研究方向:藻類(lèi)細(xì)胞培養(yǎng)及活性物質(zhì)的提取, 電話: 15589528961, E-mail: efeng 000999@163.com; 鄒寧, 通信作者, E-mail: ningzou76@126.com
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
