不同溫度對無水保活條件下的中華鱉肌肉營養成分及血液生化指標影響
2014-02-27 12:03:51黃碩琳
食品科學 2014年6期
關鍵詞:質量
何 蓉,謝 晶*,蘇 輝,黎 柳,黃碩琳
(上海海洋大學食品學院,上海水產品加工及貯藏工程技術研究中心,上海 201306)
不同溫度對無水保活條件下的中華鱉肌肉營養成分及血液生化指標影響
何 蓉,謝 晶*,蘇 輝,黎 柳,黃碩琳
(上海海洋大學食品學院,上海水產品加工及貯藏工程技術研究中心,上海 201306)
設定4、10 ℃和18 ℃組研究中華鱉(Pelodiscus sinensis)在無水保活21 d中質量損失率、肌肉水分、脂肪、蛋白質以及血液葡萄糖(Glu)、總蛋白(TP)、白蛋白(ALB)、尿素、尿酸(UA)和肌酐(CREA)含量與乳酸脫氫酶(LDH)、堿性磷酸酶(ALP)和谷草轉氨酶(AST)活性的變化。結果表明:在保活過程中,4 ℃組中華鱉僅能存活11~16 d,其質量損失率、肌肉營養物質消耗量顯著低于10 ℃和18 ℃組(P<0.05),而血液Glu、LDH、AST、UA、CREA水平均顯著高于10 ℃和18 ℃組(P<0.05);18 ℃組質量損失率、肌肉營養物質消耗量、血液尿素濃度顯著高于4 ℃和10 ℃組(P<0.05);10 ℃組肌肉營養物質消耗量及血液生理指標水平間于4 ℃和18 ℃組之間;隨著保活時間的推移,3 個溫度組肌肉營養物質和血液TP、ALB、ALP水平均呈現下降趨勢,而質量損失率和血液尿素、UA水平均呈現上升趨勢;4 ℃組肌肉ATP和血液LDH、AST、CREA水平均呈現先升高后下降的趨勢;10 ℃和18 ℃組肌肉ATP含量呈現先下降后上升的趨勢,而血液AST活性呈現下降趨勢。該實驗結果表明,4 ℃組中華鱉的營養物質損害量低,但低溫應激對該組中華鱉新陳代謝水平和生命活力產生負面影響,18 ℃組中華鱉機體能量代謝水平旺盛,營養物質消耗量和質量損失率均較大,不利于實現保活價值。建議將中華鱉保活溫度設定為10 ℃左右。
中華鱉;溫度;營養物質;生化指標
中華鱉(P e l o d i s c u s s i n e n s i s)屬于爬行綱(Repitlia)、龜鱉目(Testudinata)、鱉科(Tironychidae),俗稱甲魚、團魚、王八等,我國現已成為世界鱉養殖第一大國。我國95%以上的水產品以鮮活形式上市銷售,保活運輸是水產品由養殖基地到銷售市場的關鍵環節。目前,水產品保活技術主要有充氧保活、麻醉保活、有水保活、無水保活等[1]。中華鱉抗逆性較強,對氧氣和水分的要求量不高,易于保活,因此國內外對中華鱉無水保活的研究尚未見報道。然而隨著物流小包裝水產品禮盒的興起,禮盒裝的中華鱉夏季低溫保活的需求也被提了出來。
中華鱉具有冬眠特點,屬于變溫動物,對溫度變化較敏感,曾有研究表明溫度是影響水產品正常生理功能的重要因素之一,它不但影響其標準代謝及內源氮的代謝,而且還影響機體免疫功能及健康等[2]。溫度可能是影響中華鱉 夏季無水保活運輸的關鍵因素,鑒于此,本實驗主要研究不同溫度對中華鱉體質量損失、肌肉營養成分及血液生化指標在無水保活過程中的影響,旨在揭示中華鱉在不同保活溫度下的新陳代謝特點,為其保活運輸提供理論參考。
1 材料與方法
1.1 材料與設備
中華鱉體質量(500±50)g,于2012年11月購自上海市銅川路水產交易市場,選取體格健康、無病害的中華鱉共90只。
BPS-250CL恒溫恒濕箱 上海一恒儀器有限公司;KDN-04C自動凱氏定氮儀 上海天呈科技有限公司;DHG-9073BS-III電熱恒溫干燥器 濟南博鑫生物技術有限公司;P1000N脂肪測定儀 鄭州中谷機械設備有限公司;BS-200全自動生化分析儀 深圳邁瑞生物醫療電子股份有限公司;1515Breeze2GPC高效液相色譜儀 徳祥科技有限公司;5810R高速冷凍離心機 上海艾測電子科技有限公司。
1.2 樣品的處理
將中華鱉置于曝過氣的自來水中暫養3 d,以排盡消化道內的排泄產物,暫養環境溫度為18 ℃左右。暫養后的中華鱉經臭氧水(質量濃度為1 mg/L)浸泡1 min以殺死體表細菌,再用濕潤紗布墊裹中華鱉后分裝于專用塑料盒中,每盒一只。
1.3 實驗設計
為防止溫度驟變對中華鱉造成的不良反應,實驗前采用降溫方式(降溫速率為3~5 ℃/h)將中華鱉過度至實驗設定溫度。將分裝好的中華鱉隨機分成3 組,每組30只,分別轉移至恒定溫度為4、10 ℃和18 ℃的恒溫恒濕箱內進行無水保活實驗,濕度恒定為100%,由于中華鱉對氧氣要求不高,本實驗的恒溫恒濕箱中流動氣體為空氣。分別在中華鱉無水保活的1、6、11、16、21 d采樣,每次采樣時,每個溫度組取6 只中華鱉,4 ℃組中華鱉在保活后期出現死亡,則取活樣進行實驗。為避免糞便等污染物對中華鱉保活實驗造成的影響,每天定時觀察并及時清除糞便物。
1.4 測定指標及方法
1.4.1 質量損失率
采用質量法:質量損失率/%=(M始p-Mnp)/M始p×100
式中:M始p為實驗前編號為p的中華鱉的質量/g(p=1,2,3…90);Mnp為第n天該鱉的質量/g(n=1、6、11、16、21)。
1.4.2 肌肉的指標測定
1.4.2.1 肌肉取樣
中華鱉去頭放血后,用解剖刀去除內臟組織及四肢根部脂肪塊,取四肢及頸部的肌肉組織,用高速組織搗碎機搗碎并混勻,將肉樣分裝在若干獨立包裝袋內并置于-40℃冰箱內保存,待測指標。
1.4.2.2 一般營養成分的測定
水分含量:采用105 ℃烘箱干燥法測定,參照GB 50093—2010《食品中水分的測定》;脂肪含量:采用自動脂肪測定儀法測定,參照GB/T 14772—2008《食品中粗脂肪的測定》;蛋白質含量:采用微量凱式定氮法測定,參照GB 50095—2010《食品中蛋白質的測定》。以上3種指標的含量均以占鮮質量的質量分數表示。
1.4.2.3 腺嘌呤核苷三磷酸(adenosine triphosphate,ATP)含量的測定
參考邱偉強等[3]方法。以每克肌肉中含有ATP含量表示,單位為μg/g。
1.4.3 血液指標的測定
1.4.3.1 取血方法
中華鱉快速斷頭取血,每毫升血液用20 U肝素鈉抗凝,4 ℃、3 000 r/min離心15 min[4],取上層血漿于-20 ℃冷凍保存,待測指標。
1.4.3.2 血液指標測定方法
葡萄糖(glucose,Glu)測定采用葡萄糖氧化酶法;總蛋白(total protein,TP)測定采用雙縮脲法;白蛋白(albnmin ,ALB)測定采用溴甲酚綠法;堿性磷酸酶(alkaline phosphatase,ALP)測定采用腺嘌呤核苷酸(adenosine monophosphate,AMP)緩沖液法;乳酸脫氫酶(lactate dehydrogenase,LDH)測定采用乳酸脫氫酶測定試劑盒(國際臨床化學聯合會的推薦方法);天冬氨酸氨基轉移酶(aspartate transaminase,AST)測定采用天冬氨酸氨基轉移酶測定試劑盒(國際臨床化學聯合會的推薦方法);尿酸(urine acid,UA)采用尿酸酶-過氧化物酶法;尿素采用紫外-谷氨酸脫氫酶法;肌酐(creatinine,CREA)測定采用肌酐測定試劑盒。上述指標利用生化分析儀進行測定,所需試劑盒均購自邁瑞試劑公司。
1.5 數據處理
采用Excel進行數據處理,并采用SPSS 19.0統計軟件中的One-Way-ANOVA程序對實驗所得數據進行方差分析,差異顯著則進行Duncan氏法多重比較分析。
2 結果與分析
2.1 不同溫度下中華鱉質量損失率

圖1 不同溫度條件下鱉質量損失率的變化Fig.1 Changes in body weight loss of Pelodiscus sinensis at different temperatures
如圖1所示,隨著保活時間的延長,3個溫度組體質量損失率逐漸增大,且18 ℃組中華鱉質量損失率顯著高于4 ℃組和10 ℃組(P<0.05),在21 d時損失率近11%,這表明環境溫度越高,質量損失率越大。
2.2 不同溫度下中華鱉肌肉成分的變化
2.2.1 一般營養成分

圖2 不同溫度條件下鱉肌肉水分(A)、脂肪(B)和蛋白質(C)含量變化Fig.2 Changes in muscle water, lipid and protein content of Pelodiscus sinensis at different temperatures
如圖2所示,3 個溫度組的肌肉水分、脂肪、蛋白質含量在無水保活過程中均呈現下降趨勢,且溫度越高此3種物質消耗量越大。這主要是由于中華鱉在保活過程中維持新陳代謝需要消耗水分,加之環境蒸發耗水,肌肉水分含量隨著保活時間的推移自然減少(圖2A);4 ℃和10 ℃組脂肪、蛋白質含量在6 d時顯著降低(P<0.05)(圖2B、2C),可能是由于保活前中華鱉從常溫(18 ℃)轉移到低溫環境時經歷了溫度驟變,使中華鱉產生強烈的應激反應,為適應新的溫度環境,其新陳代謝明顯加快,作為提供能量的脂肪及蛋白質含量也就明顯減少,隨后中華鱉逐漸適應了低溫環境,其能量消耗逐漸減緩;由于18 ℃組環境溫度較高,故中華鱉營養代謝旺盛,其脂肪及蛋白質含量在保活后期顯著低于其他兩組(P<0.05)。該實驗結果與上述測出的不同溫度下中華鱉質量損失相符。
2.2.2 肌肉ATP
由圖3可知,4 ℃組ATP含量在保活過程中呈現先顯著升高后顯著降低的趨勢(P<0.05),這是由于4℃組中華鱉在保活初期因低溫應激需求,機體產生的ATP含量增多,而后因生命活力削弱,ATP含量降低,這也說明在此溫度下中華鱉僅能夠保活11~16 d;本研究中,10 ℃和18 ℃組的ATP含量在保活前11 d呈明顯下降趨勢(P<0.05),這可能是由于10 ℃和18 ℃組中華鱉起初直接動用體內存儲的ATP提供能量,隨著饑餓程度加深,機體可通過糖異生途徑生成葡萄糖,直接分解轉化為ATP。此外,高溫度組ATP含量在保活過程中大體上低于低溫組,這是由于中華鱉在高溫環境下新陳代謝旺盛,其體內ATP消耗較快所致。

圖3 不同溫度條件下中華鱉肌肉ATP含量變化Fig.3 Changes in muscle ATP content of Pelodiscus sinensis at different temperatures
2.3 不同溫度下血液生化指標的變化
2.3.1 不同溫度對中華鱉血液葡萄糖(Glu)濃度的影響

圖4 不同溫度條件下血液Glu濃度的變化Fig.4 Changes in blood Glu concentration of Pelodiscus sinensis at different temperatures
Glu是生物體內重要能源物質,其含量不僅受胰島素和腎上腺激素的控制,還受機體的各種調節,常態下機體內的Glu濃度比較恒定,當環境和機體活動發生變化,Glu濃度也會發生改變[5-6]。由圖4可知,中華鱉Glu濃度對溫度脅迫較為敏感,4 ℃和10 ℃組的Glu濃度在保活前6 d呈顯著上升趨勢(P<0.05),這與低溫脅迫對東北蛙血糖影響的結果一致[7]。在本研究中,4 ℃和10 ℃組的中華鱉在保活開始經歷了溫度驟變的過程,交感-腎上腺髓質系統則作用于中樞神經系統,以利于重要器官的血液供應,蛋白質和脂肪的消耗使糖異生途徑加強,血糖含量不斷上升,以維持溫度應激時的能量需求,而4 ℃組Glu濃度在6 d后仍然維持在較高水平,這主要是由于該溫度下的中華鱉處于冬眠狀態,其葡萄糖消耗量減少所致。曾有研究表明葡萄糖是冬眠動物抵抗寒冷環境的一種低溫保護劑[8],低溫條件下葡萄糖與膜磷脂上的磷酸基團連接形成氫鍵,從而阻止和限制細胞膜因脫水而融合,并降低相變溫度,增加膜的流動性,使膜不易向凝膠相轉變,從而保持細胞膜液晶相[9]。10 ℃組Glu濃度在6 d后呈現下降趨勢,且長期維持在保活初始水平,這說明該組中華鱉經歷短期應激后,能夠長期適應10 ℃保活環境。
2.3.2 不同溫度對中華鱉血液TP、ALB質量濃度的影響

圖5 不同溫度條件下血液TP(A)和ALB(B)質量濃度的變化Fig.5 Changes in blood TP and ALB concentrations of Pelodiscus sinensisat different temperatures
血液中的總蛋白主要包括白蛋白和球蛋白,TP主要維持血漿膠體滲透壓與血液pH值平衡,具有運輸、免疫、凝血、供能等作用[5],ALB不僅可以作為內源性氨基酸營養源,也具有相當的酸堿緩沖能力、運輸代謝產物及調節滲透壓等生理功能[10],總之,血液TP和ALB可以用來診斷動物疾病和營養狀況,且體內良好營養狀況可使血液蛋白維持在較高的水平[11]。由圖5可知,本研究中TP、ALB及上述18 ℃組Glu濃度的變化不是呈線性,而是呈波動性下降趨勢,這說明保活過程中中華鱉對能量物質的需求并非是單一的。謝小軍等[12]認為,魚類在饑餓過程中對自身貯備的能量的利用有兩個方面的適應:一方面降低代謝水平以節約能量消耗;另一方面盡可能地將代謝保存在一定水平上,以保證在重新獲得食物供應時或其他環境脅迫時能產生適當的應激反應。因此中華鱉對于供能物質的動用出現波動性起伏的變化,應該是這兩個方面因素相互交替出現的結果。圖5B顯示,4 ℃和18 ℃組的ALB質量濃度在保活初期顯著下降(P<0.05),這可能是因為溫度過低或過高使中華鱉肝臟受損,白蛋白持續下降,引發機體特異性免疫器官產生更多球蛋白。
2.3.3 不同溫度對乳酸脫氫酶(LDH)、堿性磷酸酶(ALP)、谷草轉氨酶(AST)活性的影響
LDH作為重要的無氧代謝酶,存在于機體各組織器官細胞內,它可催化乳酸和氧化性輔酶Ⅰ轉化為丙酮酸和還原性輔酶Ⅰ,進而參加機體的能量代謝。調解理論認為,LDH活性主要取決于機體細胞內氧分壓的高低,當細胞內氧分壓降低時,LDH的合成加快,其活性增強;當細胞內供氧充足時,LDH的合成受到抑制,活性降低[13]。本研究中華鱉在保活過程中,低溫度組LDH活性高于高溫度組,這可能是因為溫度較低時,中華鱉體內無氧代謝較為活躍,以彌補有氧代謝的不足;也有可能是低溫應激使能量代謝增強所致。特別指出的是,4 ℃組LDH活性在保活前11 d呈現顯著上升,而后顯著下降的趨勢(圖6A),這說明4 ℃組中華鱉在保活前期處于強烈應激狀態,機體需要更多能量克服低溫應激的危害,而后代謝機能明顯下降。
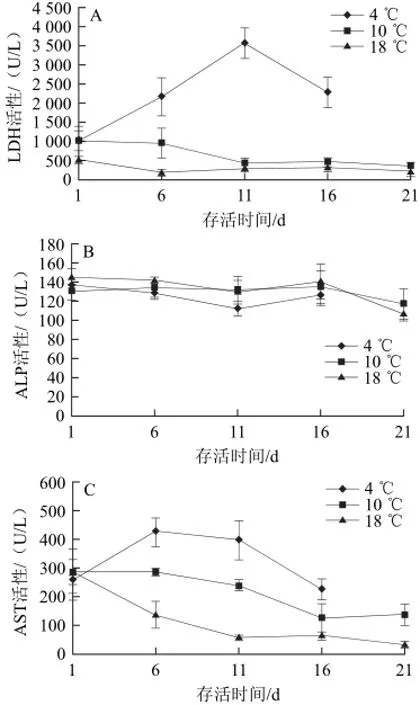
圖6 不同溫度條件下血液LDH(A)、ALP(B)和AST(C)活性變化Fig.6 Changes in blood LDH, ALP and AST activity Pelodiscus sinensis at different temperatures
ALP在機體內直接參與磷酸基團的轉移和代謝過程,在魚體內的免疫防護和物質代謝中起重要作用[14]。其活性的高低可用來判斷動物對物質消化吸收和代謝調控的能力。有關魚類的研究發現饑餓會導致草魚血液ALP活性降低[15],這與本研究結果一致。由圖6B可知,本研究中3 組中華鱉血液ALP活性在保活過程中呈下降趨勢,且18 ℃組存在顯著性差異(P<0.05),這可能是由于中華鱉處于無食物的保活環境中,機體通過降低營養代謝水平而采取的自我保護措施。
AST活性可以反應動物心肌細胞受損和代謝程度,它是機體組織細胞膜完整性的一個重要標志[16-17],一般情況下血清中AST活性較低,心、肝等內臟器官受損傷會使組織中AST釋放到血液中,而使血液中的AST活性升高[18],由圖6C可知,低溫度組AST活性在保活1 d后顯著高于高溫度組(P<0.05),這可能是低溫刺激使中華鱉心肌收縮力加強,心肌細胞代謝加快,從而使心肌受損所致;3 個溫度組AST活性均隨著保活時間的延長呈現下降的趨勢,這可能是中華鱉長期受饑餓的影響所致,有關魚類的研究也發現,饑餓脅迫會使養殖鱸魚(Lateolabrax japonicus)和哲羅魚(Hucho taimen)血液中AST活性明顯降低[19-20]。
2.3.4 不同溫度下血液尿素、尿酸(UA)、肌酐(CREA)的變化

圖7 不同溫度條件下血液尿素(A)、UA(B)和CREA(C)濃度變化Fig.7 Changes in blood urea, UA and CREA concentration of Pelodiscus sinensis at different temperatures
尿素是蛋白質和氨基酸代謝的最終產物,在蛋白代謝過程中,蛋白質被分解為氨基酸,再經去氨基化生成的氨在肝臟中合成尿素。導致血液尿素濃度升高來自兩方面的因素,一是生理性因素,如蛋白質被消耗;二是病理性因素,如腎前性、腎性及腎后性疾病等所引起[21]。圖7A顯示,隨著時間的延長,3 個溫度組的尿素濃度呈上升趨勢,曾有研究表明,在饑餓狀態下不同動物尿素的排放規律不一致,呈現下降或上升趨勢[22],本研究中18 ℃組尿素濃度顯著高于4 ℃和10 ℃組(P<0.05),且在保活11 d后尿素濃度急劇上升,這可能是因為18 ℃組中華鱉耗能多,自身蛋白質代謝率高,該結果與該溫度下肌肉蛋白質消耗量最大的結果相一致;UA是嘌呤分解代謝產物,CREA主要是體內肌酸的代謝產物,這兩種物質亦由腎臟代謝排出;由圖7B可以看出,隨著保活時間的延長,4 ℃和10 ℃組的UA濃度積累越高,這與劉偉東等[23]研究的低溫無水保活條件下大菱鲆(Scophthalmus maximus)血液UA濃度的變化趨勢類似;由圖7B、7C所知,4 ℃組UA和CREA濃度在保活過程中明顯高于其他兩組(P<0.05),這說明低溫環境對中華鱉腎臟功能造成一定的損傷。
3 結 論
本研究主要研究了3 個溫度組中華鱉在保活21 d過程中質量損失率、肌肉營養物質及血液生理指標的變化特點,分析結果表明,4 ℃組中華鱉ATP、Glu、LDH、AST、CREA等含量在保活初期維持在較高水平,說明該溫度組中華鱉在保活初期代謝活躍從而應對低溫應激,由于低溫應激過于強烈,體內能量代謝能力和生命活力在保活后期明顯削弱,僅能存活11~16 d;18 ℃組中華鱉新陳代謝水平在保活過程中明顯高于其他兩組,最終出現消瘦現象,質量損失率近11%,但至實驗結束都能存活;10 ℃組中華鱉在保活初期表現出短暫的低溫應激,而后逐漸適應了該溫度環境,其新陳代謝水平在保活過程中較低,最終體質量損失率約為3.7%,在實驗期間存活良好。從本研究結果可以看出,中華鱉適宜保活溫度約為10 ℃。
[1] 何蓉, 謝晶. 水產品保活技術研究現狀和進展[J]. 食品與機械, 2012, 28(5): 243-246.
[2] MARTINEZ-PALACIOS C A, TOVAR B E, TAYLOR J F, et al. Effect of temperature on growth and survival of Chirostoma estor estor, Jordan 1879, monitored using a simple video technique for remote measurement of length and mass of larval and juvenile fishes[J]. Aquaculture, 2002, 209(1/4): 369-377.
[3] 邱偉強, 陳剛, 陳瞬勝, 等. 離子對反相高效液相色譜法同時檢測水產品中6 種ATP關聯化合物[J]. 水產學報, 2011, 35(11): 1745-1751.
[4] 廖曉霞, 葉均安. 中華鱉的消化道指數和血液生化指標[J]. 上海水產大學學報, 2006, 15(1): 105-108.
[5] 周玉, 郭文場, 楊振國, 等. 魚類血液學指標研究的進展[J]. 上海水產大學學報, 2001, 10(2): 163-165.
[6] 孫學亮, 邢克智, 陳成勛, 等. 急性溫度脅迫對半滑舌鰨血液指標的影響[J]. 水產科學, 2010, 29(7): 387-392.
[7] 李秀峰. 低溫脅迫下東北林蛙血糖和相關酶的探究[D]. 哈爾濱: 東北林業大學, 2012.
[8] 劉云, 楊小林, 王朝莉. 中華鱉冬眠期與活動期部分生理生化性質的比較研究[J]. 四川大學學報, 2002, 39(增刊1): 82-84.
[9] 李秀峰. 低溫脅迫下東北林蛙血糖和相關酶的研究[D]. 哈爾濱: 東北林業大學, 2012.
[10] LUK’YANENKO V I, KHABAROV M V, LUK’YANENKO V V. Heterogeneity and polymorphism of functionally specialized blood proteins in migratory fi sh: the case study of the north caspian population of the Russian sturgeon during sea and river periods of life. 1. Albumins[J]. Biology Bulletin, 2002, 29(3): 293-299.
[11] MOMMSEN T P, FRENCH C J, HOCHACHKA P W. Sites and patterns of protein and amino acid utilization during the spawning migration of salmon[J]. Canadian Journal of Zoology, 1980, 58(10): 1785-1799.
[12] 謝小軍, 鄧利, 張波. 饑餓對魚類生理生態學影響的研究進展[J]. 水生生物學報, 1988, 22(2): 181-187.
[13] 李永才, 黃溢明. 比較生理學[M]. 北京: 高等教育出版社, 1993: 123-134. [14] 蔣春琴, 冷向軍, 李小勤, 等. 醋酸棉酚對異育銀鯽生長性能血清生化指標和棉酚殘留的影響[J]. 動物營養學報, 2012, 24(1): 168-175.
[15] 華雪銘, 朱站英, 邢思華, 等. 饑餓對草魚非特異免疫水平的影響[J].動物學雜志, 2012, 47(2): 91-97.
[16] JYOTHI B, NARAYAN G. Effect of phorate on certain protein prof i les of serum in freshwater fi sh, Clarias batrachus (Linn.)[J]. Journal of Environmental Biology, 2001, 22(3): 233-235.
[17] 崔杰峰, 潘柏申. 急性心肌梗死血清酶標志物沿革[J]. 上海醫學檢驗雜志, 2000, 15(1): 13-14.
[18] 林仕梅, 羅莉, 葉元士. 喹乙醇對草魚幾項生理指標的影響[J]. 西南農業學報, 1998, 11(2): 119-123.
[19] 錢云霞, 陳惠群, 孫江飛. 饑餓對養殖鱸魚血液生理生化指標的影響[J]. 中國水產科學, 2002, 9(2): 133-136.
[20] 楊成輝, 蔡勛, 劉霞, 等. 饑餓和再投喂對哲羅魚幼魚血液生理生化指標的影響[J]. 淡水漁業, 2009, 39(1): 36-40.
[21] LI S Z, YUAN X J, YANG Y Y, et al. Effect of cold stress by immersing in water on some enzyme activities and digestive tract mucosal hyperemia in chicken[J]. Chinese Journal of Applied and Environmental Biology, 2001, 7(3): 267-27 0.
[22] RIOS F S A, MORAES G, OBA E T, et al. Mobilization and recovery of energy stores in traira, Hoplias malabaricus Bloch (Teleostei, Erythrinidae) during long-term starvation and after re-feeding[J]. Journal of Comparative Physiology, 2006, 176(7): 721-728.
[23] 劉偉東, 薛長湖, 殷邦忠, 等. 低溫下大菱鲆有水和無水保活過程中生理生化變化的研究[J]. 漁業科學進展, 2009, 30(5): 69-74.
Effect of Temperature on Muscle Nutritional Components and Blood Biochemical Parameters of Pelodiscus sinensis Alive without Water
HE Rong, XIE Jing*, SU Hui, LI Liu, HUANG Shuo-lin
(Shanghai Aquatic Products Processing and Storage Engineering Technology Research Center, College of Food Science and Technology, Shanghai Ocean University, Shanghai 201306, China)
Pelodiscus sinensis was kept alive without water by incubation in a thermostatic chamber set at different temperatures (4, 10 and 18 ℃) for 21 days. The changes in body weight loss, the contents of water, lipid and protein in muscle, and the concentrations of glucose (Glu), total protein (TP), albumin (ALB), urea, uric acid (UA) and creatinine (CREA) as well as the activities of lactate dehydrogenase (LDH), alkaline phosphatase (ALP) and aspartate aminotransferase (AST) in blood were examined during the incubation period. The results indicated that the survival time of Pelodiscus sinensis at 4 ℃ was 11–16 days. The body weight loss and nutrient consumption of Pelodiscus sinensis at this temperature were lower than those observed at 10 and 18 ℃ (P < 0.05) and the concentrations of Glu, UA and CREA, and the activities of LDH and AST were also higher (P < 0.05). The body weight loss, nutrient consumption and blood urea concentration of Pelodiscus sinensis at 18 ℃ were higher than those observed at 4 and 10 ℃ (P < 0.05). Additionally, the nutrient consumption and blood physiological indexes of Pelodiscus sinensis at 10 ℃ were between those at 4 and 18 ℃. The nutrition, blood TP and ALB levels and ALP activity of Pelodiscus sinensis at three different temperatures showed a decreasing trend during the incubation period, while the body weight loss, and urea and UA levels exhibited an increasing trend. The levels of ATP, LDH, AST and CREA in Pelodiscus sinensis at 4 ℃ increased initially and then decreased.Furthermore, ATP content of Pelodiscus sinensis at 10 and 18 ℃ exhibited an initial decrease followed by a increase, while AST activity always decreased. These results suggest that the nutrient consumption of Pelodiscus sinensis at 4 ℃ is lower, while low temperature stress has a harmful effect on the metabolism and vitality of Pelodiscus sinensis. The body metabolism rate of Pelodiscus sinensis at 18 ℃ is faster and the nutrient consumption and body weight rate are higher. Therefore, the most appropriate temperature for keeping Pelodiscus sinensis alive is about 10 ℃.
Pelodiscus sinensis; temperature; nutrition; biochemical parameters
TS254.4
A
1002-6630(2014)06-0194-06
10.7506/spkx1002-6630-201406042
2013-05-19
“十二五”國家科技支撐計劃項目(2012BAD38B04);上海市科委農業重點項目(11dz1960400);上海市科委工程中心建設項目(11DZ2280300);上海高校一流學科建設計劃項目
何蓉(1988—),女,碩士研究生,研究方向為中華鱉的保活。E-mail:herong2007@126.com
*通信作者:謝晶(1968—),女,教授,博士,研究方向為水產品保鮮保活。E-mail:jxie@shou.edu.cn
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
