三葉爬山虎葉片解剖結構和光合生理特性對3種生境的響應
2014-02-25 03:33:28吳濤耿云芬柴勇郝佳波袁春明
生態環境學報 2014年10期
吳濤,耿云芬,柴勇,郝佳波,袁春明*
1. 云南省森林植物培育與開發利用重點實驗室,國家林業局云南珍稀瀕特森林植物保護和繁育實驗室,云南 昆明 650201;2. 云南省林業科學院,云南 昆明 650201
三葉爬山虎葉片解剖結構和光合生理特性對3種生境的響應
吳濤1,2,耿云芬1,2,柴勇2,郝佳波1,2,袁春明1,2*
1. 云南省森林植物培育與開發利用重點實驗室,國家林業局云南珍稀瀕特森林植物保護和繁育實驗室,云南 昆明 650201;2. 云南省林業科學院,云南 昆明 650201
藤本植物生活環境的時空變化較為劇烈,為適應異質性生境常表現出較大的可塑性,其形態解剖結構及光合生理特征被認為能很好地體現對異質生境的適應。為了明確藤本植物葉片結構和光合作用對不同生境光強的響應策略,以木質藤本三葉爬山虎(Parthenocissus himalayana)為對象,采用光合儀測定和解剖顯微觀察的方法研究了哀牢山亞熱帶濕性常綠闊葉林的林外(全光照)、林緣(遮蔭)和林內(蔭生)3種自然生境中三葉爬山虎的葉片解剖結構和光合生理特征的變化,以期闡述三葉爬山虎對不同光環境的生態適應能力及策略,為森林生態系統的管理和物種多樣性的保護及群落優化配置提供理論依據。結果表明:從林內到林外隨著生境光強增加,葉片厚度(157.77~299.17 μm)、上表皮厚度(21.30~28.40 μm)、柵欄組織厚度(30.83~124.65 μm)、柵欄組織細胞面積(430.95~652.97 μm2)顯著增大(P<0.01),柵欄組織細胞長度(29.23~49.54 μm)和周長(86.58~155.17 μm)、下表皮厚度(16.14~19.01 μm)、氣孔長度(24.13~27.10 μm)和氣孔密度(86.20~129.41個·mm-2)呈顯著上升趨勢(P<0.05)。柵欄組織細胞寬度(19.67~22.81 μm)在3種生境中無顯著差異。葉片解剖結構性狀的平均可塑性值為0.37,其中最大值是柵欄組織細胞長度(0.67),最小值是氣孔長度(0.11)。光飽和點(201.27~1299.17 μmol·m-2·s-1)、光補償點(3.86~29.88 μmol·m-2·s-1)、飽和光強最大光合速率(2.20~12.03 μmol·m-2·s-1)、暗呼吸速率(0.17~2.19 μmol·m-2·s-1)、CO2補償點(83.01~237.26 μmol·m-2·s-1)、飽和CO2最大凈光合速率(2.07~25.49 μmol·m-2·s-1)、光呼吸速率(0.36~7.57 μmol·m-2·s-1)、初始羧化效率(0.006~0.035 μmol·μmol-1)隨著生境光強的增高呈上升趨勢,而表觀量子效率(0.067~0.031 μmol·μmol-1)、CO2飽和點(2062.56~1385.31 μmol·m-2·s-1)呈下降趨勢。與光合生理參數相關性顯著的解剖性狀主要是葉片厚度、上表皮厚度及其細胞周長/面積、柵欄組織細胞長度、柵欄組織厚度及維管束占葉脈面積的比例等。研究說明,三葉爬山虎在林外生境中具有明顯的陽生葉特征,而在林內生境中具有明顯的陰生葉特征,表現出對異質生境很強的適應性,從葉片解剖結構和光合生理特性方面解釋了其在林外、林緣和林內均有分布的原因。
三葉爬山虎;葉片;解剖結構;光合特性;生境
木質藤本是森林植被中一類豐富多樣但具有共同生長策略即攀援生長的植物類群,它們在森林的多樣性、結構、功能和動態等方面具有重要的貢獻和作用(王海艷和喻理飛,2010)。木質藤本植物由于受攀援能力和支柱木資源的限制,以及時空變化十分劇烈的生境特點,其形態和生理特征常發生明顯的變化,表現出較大的可塑性。其中,葉片作為藤本植物對環境變化最為敏感的光合器官之一,其形態解剖結構及光合生理特征被認為最能體現環境因子的影響及植物對環境的適應(Mott等,1982;烏玉娜等,2010;龐杰等,2013)。亞熱帶常綠闊葉林林外、林緣、林內的生境具有較大的異質性,其中光照是影響藤本植物多度和分布的主要生境因子。木質藤本若要在這樣較大異質性的生境中生長,較高的可塑性無疑將使其對環境具有較高的潛在適應能力,這對木質藤本在異質光環境中的生存至關重要(Strauss-Debenedetti和Bazzaz,1991)。
木質藤本以其特有的植物學特性和生態功能,在水土保持、防風固沙、減輕污染、垂直綠化和美化環境等方面異軍突起,已經并將繼續發揮其他植物不可替代的作用(李益鋒等,2011)。然而,其有關研究工作在過去20年來才開始不斷出現(Gentry,1991;蔡永立和宋永昌,2000)。目前有關木質藤本植物生態適應研究的一些結論主要來自于對熱帶藤本的研究,對其它地區藤本的研究則相對不足(蔡永立和宋永昌,2001);木質藤本植物間及其與木本植物間的比較生態研究較多,而對同種藤本植物不同生境下的光合生理和形態可塑性研究較少(夏江寶,2007;陳亞軍等,2008);研究結果大多是在人工控制條件下獲得的,缺少對木質藤本植物在野外原生環境中葉片形態結構及光合生理特征的觀察和測定(江浩等,2011)。因此,本項研究以亞熱帶常綠闊葉林常見木質藤本三葉爬山虎(Parthenocissus himalayana)為材料,通過對其在不同生長光強的生境下葉片的光合作用和葉片解剖特征的研究,揭示其在不同光環境下的生態適應能力及策略,可為森林生態系統的管理和物種多樣性的保護及群落優化配置提供科學依據。
1 材料與方法
1.1 研究地區自然概況
研究地點位于云南中南部景東縣徐家壩的哀牢山國家自然保護區內的中國科學院哀牢山生態站附近的亞熱帶濕性常綠闊葉林,地理位置為24°32′ N,101°01′ E,海拔2450 m。本區屬西南季風性氣候,終年溫涼潮濕。年平均溫度為11.3 ℃,最冷月(1月)平均氣溫5.0 ℃,最熱月(7月)平均氣溫15.3 ℃,極端最高氣溫25.0 ℃,極端最低氣溫-8.3 ℃。該區年降水量為1931.1 mm,雨季(5—10月)降水量占年降水總量的85%左右。年平均相對濕度為83%。≥10 ℃的的積溫達3420 ℃,全年無霜期約180 d,日照時數為1239 h。林地土壤為山地黃棕壤。實驗在8月上旬開展,最高溫度為32.1 ℃,最低溫度為16.0 ℃,正午最大光強可達2200 μmol·m-2·s-1;坡向為西坡,坡度10°~23°。土壤肥沃,偏酸性(pH為4.4~4.9),土壤表面由凋落物層(厚度3~7 cm)覆蓋,土壤有機碳、全氮和全磷含量分別為12.91%、0.52%和0.06%,土壤表面的C/N比為14.4,30~50 cm深度C/N比為15.3(Liu等,2002.)。
研究地點所在研究區內有直徑不小于1 cm、高度不小于2 m的木質藤本23種,隸屬于16科、21屬(袁春明等,2010)。在研究地點選取一個沒有新近林窗干擾的森林邊緣,并且其林緣、林內(林緣向內50 m外)和林外(林緣向外50 m外)均有自然生長的三葉爬山虎(直徑1~2 cm)。林外、林緣、林內3種生境上午10:00時的光照強度和溫度分別為1800 μmol·m-2·s-1和28 ℃、600 μmol·m-2·s-1和 25 ℃、100 μmol·m-2·s-1和22 ℃。
1.2 實驗材料
選取林外、林緣、林內3種生境的三葉爬山虎植株距離地面60~100 cm處枝條的頂端向下第2~3片成熟葉為光合測定對象,每株1片,每種生境3~5個重復。光合測定結束后,將該葉片于中央位置處沿中脈剪取約1 cm2的樣品放入FAA固定液,用于葉片解剖結構觀察。
1.3 研究方法
1.3.1 光合生理特征測定
2013年8月10—16日,采用Li-6400便攜式光合測定儀(LI-COR,USA),配合Li-6400-02B紅藍光源和Li-6400-01液化CO2鋼瓶注入系統,對3種生境內相同地面高度的三葉爬山虎葉片進行光合-光響應曲線(Pn-PAR)和光合-CO2響應曲線(Pn-Ci)的測定。Pn-PAR曲線測定時,設置光合有效輻射梯度為0、25、50、75、100、150、200、300、400、600、800、1200、1600、2000 μmol·m-2·s-1,調整CO2濃度為400 μmol·mol-1,運用自動測量程序測定;Pn-Ci曲線測定時,光合有效輻射設定為1200 μmol·m-2·s-1,設置CO2濃度梯度為400、300、200、150、100、50、0、400、600、800、1200、1600 μmol·mol-1,運用自動測量程序測定。選取無病害植株的相同葉位的葉片分別掛牌標記,測定3個重復,測定時保持葉片自然生長角度不變。測定時間為9:00—12:00。根據Ye等(2012)的非直角雙曲線修正模型擬合Pn-PAR曲線,計算飽和光強最大光合速率(Pmax)、光飽和點(LSP)、光補償點(LCP)、表觀量子效率(AQY)和暗呼吸速率(Rd)。根據葉子飄和于強(2009)的非直角雙曲線修正模型擬合Pn-Ci曲線,計算飽和CO2最大光合速率(Amax)、CO2飽和點(Csp)、CO2補償點(Ccp)、表觀羧化效率(CE)和光呼吸速率(Rp)。
1.3.2 葉片解剖結構觀察
光合生理特征測定后,剪取該葉片主脈及其兩側1.0 cm×1.0 cm的小塊,用FAA固定,3個重復。采用常規石蠟法制片,在Leica顯微鏡下觀察和拍照,測量并記錄葉片的各結構,所得數據為10個視野的平均值。所觀測的葉片解剖結構特征包括葉片厚度、上表皮厚度、上表皮細胞面積、上表皮細胞周長、柵欄組織厚度、柵欄組織細胞寬度、柵欄組織細胞長度、柵欄組織細胞周長、柵欄組織細胞面積、海綿組織厚度、下表皮厚度、下表皮細胞周長、下表皮細胞面積、維管束面積占葉脈面積的比例、氣孔長度、氣孔密度,共16項。可塑性指數=(某一指標或參數在3種生境下的最大值 - 最小值)/最大值。
1.3.3 數據處理和分析
運用SPSS 19.0進行數據處理和模式擬合及相關參數的計算,對3種生境下各指標或參數進行方差分析時用LSD方法。
2 結果與分析
2.1 葉片解剖結構特征在不同生境下的差異
三葉爬山虎在林外、林緣和林內3種生境下的葉片解剖結構見圖1,觀測結果見表1。可以看出,其葉片類型屬于異面葉,葉肉組織分化為明顯的柵欄組織和海綿組織,但各特征在3種生境條件下存在明顯的差異。三葉爬山虎葉脈背面的腺毛在林外生境下較林緣和林內更明顯且常見。林外生境下,柵欄組織細胞緊鄰上、下表皮均有分布,上表皮處排列緊密,2~3層,下表皮處排列稀疏,1~2層;林緣生境下,柵欄組織細胞緊鄰上表皮單層分布且排列較為疏松;林內生境下,柵欄組織細胞單層分布且排列十分疏松。
本研究所觀測的16項葉片解剖結構特征,除柵欄組織細胞寬度在3種生境中無顯著差異外,其余15項指標均存在顯著或極顯著差異。隨著生境光照強度的降低,葉片厚度、上表皮厚度、柵欄組織厚度、柵欄組織細胞面積呈極顯著下降,柵欄組織細胞長度/周長、下表皮厚度、氣孔長度和氣孔密度整體上呈顯著下降趨勢。海綿組織厚度在林外和林內相近,二者之間差異不顯著,但較林緣的高且差異顯著。下表皮細胞周長及面積以林緣最高且與林外和林內差異顯著,林內和林外之間差異不顯著。主脈維管束面積占葉脈面積的比例為林外>林內>林緣,兩兩之間差異顯著。
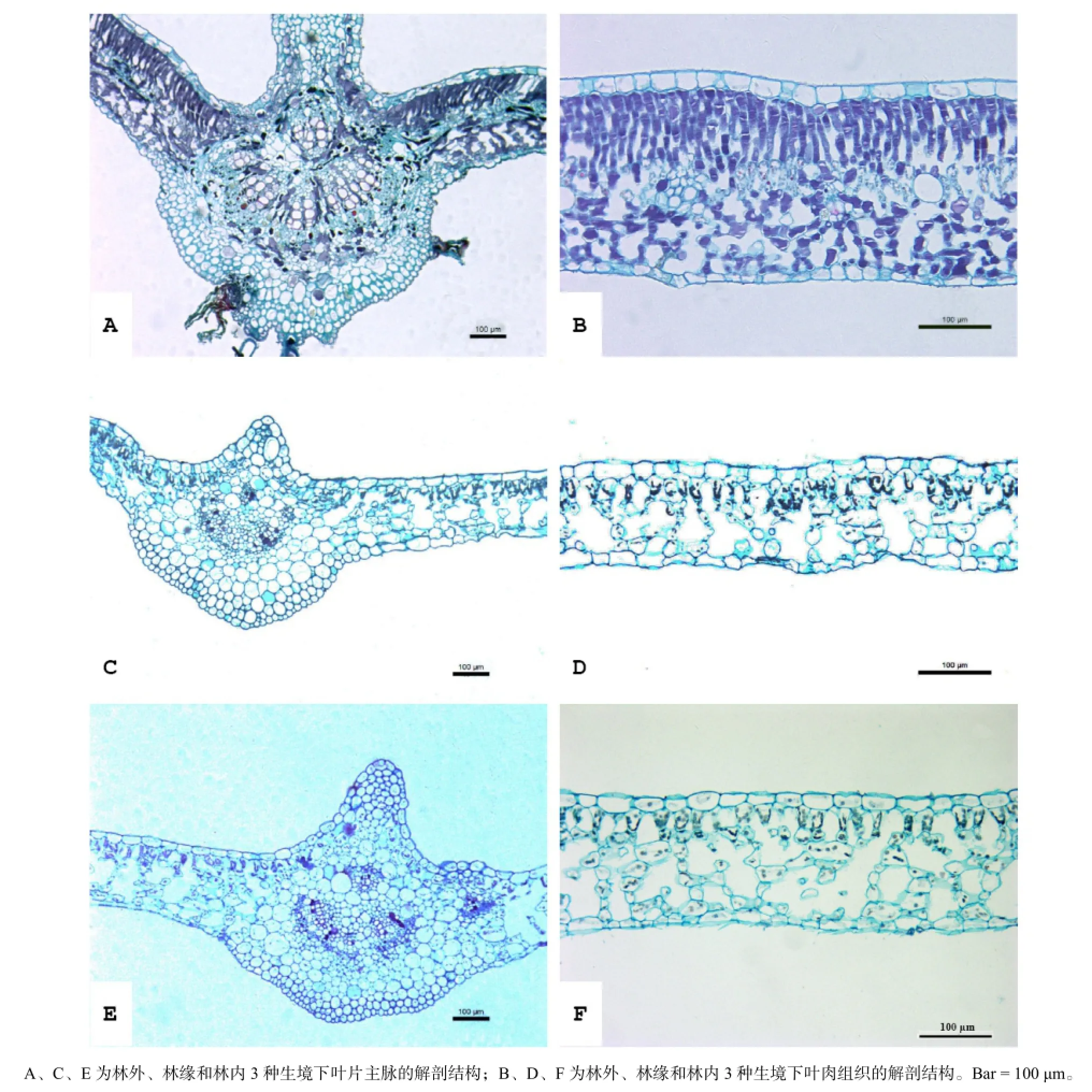
圖1 三葉爬山虎在林外、林緣和林內3種生境下的葉片解剖結構Fig.1 Leaf anatomical structure of Parthenocissus himalayana in three different habitats

表1 三葉爬山虎在3種生境下的葉片解剖特征比較Table 1 The comparatives of leaf anatomical structure of Parthenocissus himalayana in three different light habitats
各指標的表型可塑性從大到小,依次為柵欄組織細胞長度>柵欄組織厚度>柵欄組織細胞周長>維管束面積占葉脈面積的比例>柵欄組織細胞面積>葉片厚度>上表皮細胞面積>下表皮細胞面積>氣孔密度>上表皮細胞周長>上表皮厚度>柵欄組織細胞寬度>下表皮細胞周長>下表皮厚度>海綿組織厚度>氣孔長度。綜合方差分析和表型可塑性分析結果可以看出,柵欄組織細胞厚度/長度/周長/面積、葉片厚度、上表皮厚度、氣孔密度和維管束面積占葉脈面積的比例是最能反映三葉爬山虎對生境響應的指標。
2.2 光合生理特性在不同生境下的差異
光補償點越小,表明植物利用弱光的能力越強;而光飽和點越高則表明植物適應強光的能力越強。光合-光響應曲線參數(表2)表明,隨著生境光強的降低,三葉爬山虎的光飽和點和光補償點呈極顯著下降趨勢,而最大凈光合速率、暗呼吸速率和表觀量子效率呈顯著下降趨勢。三葉爬山虎林緣的最大凈光合速率下降至林外的39%,表觀量子效率降至其的56%,光飽和點降至其的59%,光補償點降至其的25%,暗呼吸速率降至其的21%,表明三葉爬山虎從林外到林緣,主要通過降低光補償點和暗呼吸速率來減緩其凈光合速率的快速下降。
光合-CO2濃度響應曲線參數(表3)表明,三葉爬山虎林外的飽和CO2飽和凈光合速率分別是林緣和林內的3.23倍和12.31倍,初始羧化效率分別是其4.38倍和5.83倍,光呼吸速率分別是其17.60倍和21.03倍,CO2補償點分別是其3.32倍和2.86倍,而CO2飽和點分別是其78%和67%。參數顯示,三葉爬山虎處于林緣和林內生境時,雖然飽和CO2凈光合速率較林外生境顯著降低,但其CO2補償點也顯著降低并且可利用的CO2濃度區間范圍更大,光呼吸速率顯著降低,即三葉爬山虎主要通過降低CO2補償點和光呼吸速率來適應其飽和CO2凈光合速率在林緣和林內生境時光強的減弱。

表2 三葉爬山虎在3種生境下的光合-光響應曲線特征參數Table 2 The parameters of light response curve (Pn-PAR) of Parthenocissus himalayana in three different light habitats

表3 三葉爬山虎在3種生境下的光合-CO2響應曲線特征參數Table 3 The parameters of CO2response curve (Pn-Ci) of Parthenocissus himalayana in three different light habitats
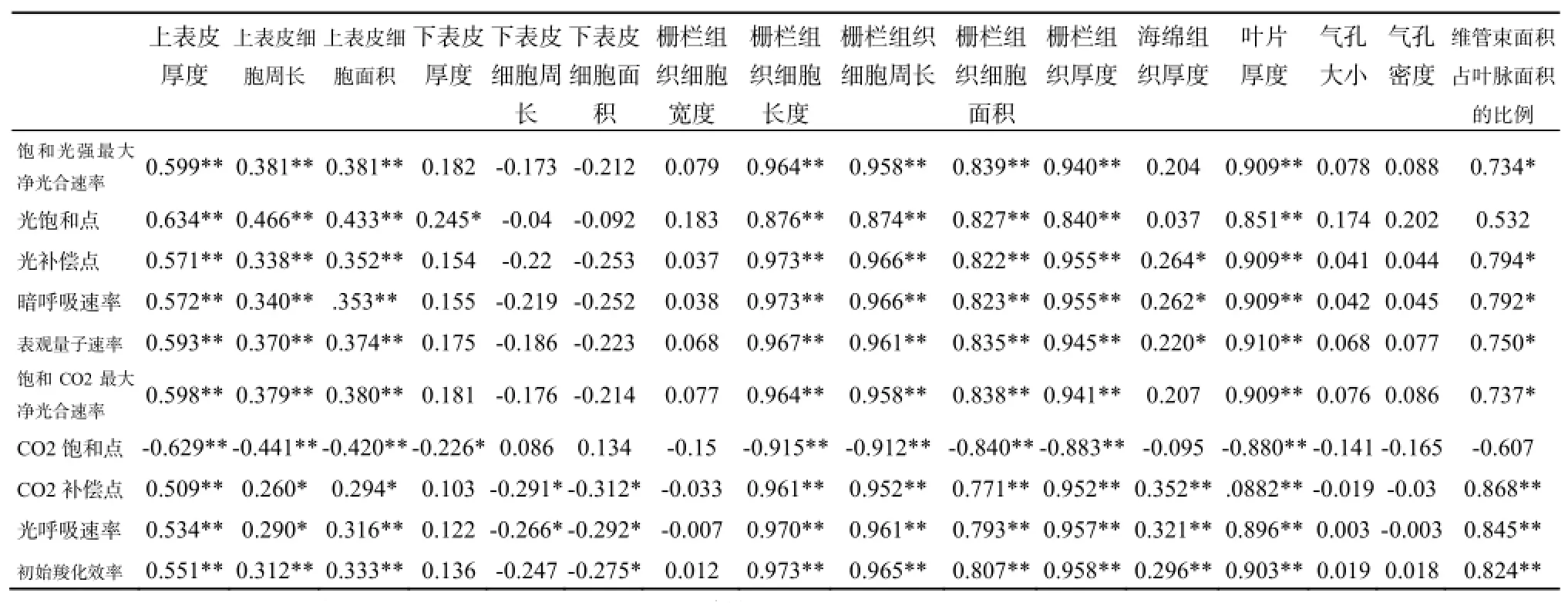
表4 不同生境下三葉爬山虎葉片解剖結構和光合生理參數之間的相關性分析Table 4 The correlation analysis between photosynthetic parameters and anatomical structures characteristics of Parthenocissus himalayana in three different light habitats
2.3 不同生境下三葉爬山虎葉片解剖結構和光合生理參數的關系
如表4所示,10個光合生理參數與16個葉片解剖性狀之間的160對相關性分析結果中,顯著性相關(P<0.05)18對,極顯著相關(P<0.01)83對。其中,與光合生理參數相關性顯著和極顯著的解剖性狀主要是上表皮厚度及其細胞周長/面積、柵欄組織細胞長度/周長/面積、葉片厚度、柵欄組織厚度及維管束占葉脈面積的比例;海綿組織厚度、氣孔大小及密度、下表皮厚度及其細胞周長/面積、柵欄組織細胞寬度與光合生理參數幾乎無顯著(P<0.05)相關性。說明海綿組織厚度和氣孔大小及其密度等解剖結構不影響三葉爬山虎在不同光強生境下的光合作用,而上表皮厚度及其細胞周長/面積、柵欄組織厚度及其細胞長度/周長/面積等解剖結構則有較明顯的影響。
3 討論
3.1 解剖結構對不同生境的響應
葉片解剖結構能夠反映植物對環境變化的響應(李芳蘭和包維楷,2005;季子敬等,2013)。厚葉結構、發達的柵欄組織、排列緊密平直的表皮細胞是陽生葉的典型特征,而發達的海綿組織、凸透形的表皮細胞則是葉片對于弱光、水分充足的一種適應特性(賀金生等,1994;王榮和郭志華,2007;陳模舜和柯世省,2013)。本項研究中,三葉爬山虎葉片厚度表現出隨生境光強增強而變厚的趨勢,且差異極顯著;林外葉片厚度為林內的1.90倍。造成其差異的主要原因是柵欄組織厚度和表皮(尤其是上表皮)厚度的增加;柵欄組織厚度的增加主要由柵欄組織細胞層數增加所致,柵欄組織細胞長度也起到一定作用。海綿組織厚度表現出隨生境光強減弱而增大的趨勢,差異顯著。此外,隨生境光強減弱柵欄組織細胞排列呈現出由緊密向疏松變化的趨勢;形狀由長棒狀趨于短錐狀;表皮細胞的細胞壁由平直趨于弧形彎曲(圖1)。這些結果表明,三葉爬山虎在林外生境中具有明顯的陽生葉特征,而在林內生境中表現出明顯的陰生葉特性,表現出對異質生境的良好適應性。
植物生長在高光強下氣孔密度一般都比弱光強下高,如楓香(Liquidambar formosana) (王榮和郭志華,2007)和白三葉(Trifolium repens)(史剛榮和蔡慶生,2006),與本研究結果一致,推測這樣有利于改善在較高光強下由于具有較大的葉肉表面積而造成CO2擴散受阻的情況(史剛榮和蔡慶生,2006)。但豚草(Ambrosia cordifolia)葉片在遮蔭條件下為氣孔下生,在高光強時則為氣孔雙生,且上表面的氣孔密度在高光強時降低,而下表面的則增加(Mott等,1982)。這說明光強對氣孔密度的影響因物種不同而不同。
3.2 光合特性對不同生境的響應
光合-光響應和光合-CO2響應參數提供了植物對變化的環境條件適應能力的光合特性信息,反映了植物對光照強度和生境大氣CO2濃度的適應能力,在一定程度上決定了植物的分布模式和物種豐度。植物的光飽和點(LSP)表示對光照強度的需要量,其值越高表示對光強的需求越大。植物光補償點(LCP)和表觀量子效率(AQY)反映其對弱光的適應能力,LCP值越小、AQY越大,表明植物利用弱光的能力越強(柴勝豐等,2013;陳模舜和柯世省,2013)。本項研究中,三葉爬山虎的LSP在3種光照生境下差異顯著,林外的LSP是林緣的1.7倍、林內的6.5倍。林外的光補償點LCP顯著高于林緣,但林緣和林內差異不顯著。三葉爬山虎在林外的LSP高而林內的LCP低,說明其對光強范圍的適應能力廣;其在林內的LSP和LCP均顯著低于林外,而林緣和林內的AQY顯著高于林外,說明其對弱光的利用能力強,既能適應林外的全光照光強又能適應林內遮蔭條件。這與夏江寶(2007)的研究結果一致,其認為三葉爬山虎耐蔭性較強,維持較高光合速率的適宜光強范圍為600~1600 μmol·m-2·s-1。
3.3 三葉爬山虎對生境光強的適應策略
植物表型可塑性與其環境適應能力關系緊密,是衡量植物對環境響應的重要指標,可塑性值大在一定程度上表明植物適應異質生境的能力強(薛靜,2010)。楓香的葉片解剖特征在異質光生境間的總體可塑性值為0.30(王榮和郭志華,2007),白三葉的為0.26(史剛榮和蔡慶生,2006)。三葉爬山虎的總體可塑性值為0.37,大于楓香和白三葉,一定程度上表明藤本植物對異質光生境的適應能力強于木本植物和草本植物。另外,三葉爬山虎的可塑性值最大的是柵欄組織細胞長度,可塑性值較小的是氣孔長度、海綿組織厚度和下表皮厚度,均與楓香和白三葉相同,表明植物在葉片解剖結構上響應異質光生境的策略相同,主要是通過增加柵欄組織細胞長度和柵欄組織厚度增加葉片厚度,從而適應更強的光照環境。柵欄組織細胞長度和層數在植物適應異質光環境方面起到了極其重要的作用。
三葉爬山虎在哀牢山常綠闊葉林的分布模式為在林窗邊緣(等同于本研究中的林緣)和林內分布較多而在林窗(等同于本研究中的林外)較少(袁春明等,2008)。本項研究中,在10:00~14:00光照較強時,林外生境的光強有時會超過光飽和點,林緣生境的光強接近但小于光飽和點,林內生境的光強則僅約為光飽和點的一半,三葉爬山虎在林外比林緣和林內更整齊平直的上表皮細胞、更長和更多層數的柵欄組織細胞、更大的光飽和點,這些均可視為其對林外高光強和林內弱光強的適應,因此可以認為三葉爬山虎能夠適應林外、林緣和林內的光照條件,而其在林外分布較少可能是攀附生長所需的支持物缺少的關系(袁春明等,2010)。
4 結論
1)三葉爬山虎表現出對異質光強生境的良好適應性,在林外全光照生境下呈現厚葉結構、發達的柵欄組織、排列緊密平直的表皮細胞等陽生葉特征;在林內蔭蔽生境下呈現葉片變薄、海綿組織發達、柵欄組織細胞排列疏松、表皮細胞壁弧形彎曲等陰生葉特征。造成三葉爬山虎葉片厚度隨生境光強的增強而顯著變厚的主要原因是柵欄組織厚度和表皮(尤其是上表皮)厚度的增加;同時,柵欄組織厚度的增加主要由柵欄組織細胞層數增加所致,柵欄組織細胞長度也起到一定作用。
2)從林外到林內,三葉爬山虎主要通過降低光補償點、暗呼吸速率、CO2補償點和光呼吸速率,增加CO2濃度使用區間來適應逐漸減弱的光強生境。
3)相關性分析表明,與光合生理參數相關性極顯著/顯著的葉片解剖性狀主要是上表皮厚度及其細胞周長/面積、柵欄組織細胞長度/周長/面積、葉片厚度、柵欄組織厚度及維管束占葉脈面積的比例;柵欄組織細胞寬度、氣孔大小及密度與光合生理參數的相關性不顯著。
4)可塑性值分析表明,三葉爬山虎從葉片解剖結構上響應異質光生境的策略主要是通過增加柵欄組織細胞長度和柵欄組織厚度以增加葉片厚度,從而適應更強的光照環境。柵欄組織細胞長度和層數在植物適應異質光環境方面起到了極其重要的作用。
致謝:試驗樣地選擇和光合生理參數的測定得到中國科學院哀牢山森林生態系統研究站的大力支持,特表感謝!
GENTRY A H. 1991. The distribution and evolution of climbing plants[A]//PUTZ F E and MOONEY H A. The Biology of Vines[M]. Cambridge: Cambridge University Press: 3-49.
LIU W Y, FOX J E D, XU Z F. 2002. Biomass and nutrient accumulation in montane evergreen broad-leaved forest (Lithocarpus xylocarpus type) in Ailao Mountains, SW China[J]. Forest Ecology and Management, 158: 223-235.
MOTT K A, Gibson A G, O' Leary J W. 1982. The adaptive significance of amphistomatic leaves [J]. Plant Cell Environ, 9: 455-460.
STRAUSS-DEBENEDETTI S, Bazzaz F A. 1991. Plasticity and acclimation to light in tropical Moraceae of different successional positions [J]. Oecologia, 87: 377-387.
YE Z P, YU Q, KANG H J. 2012. Evaluation of photosynthetic electron flow using simultaneous measurements of gas exchange and chlorophyll fluorescence under photorespiratory conditions[J].Photosynthetica, 50(3): 472-476.
YE Z P. 2012. Nonlinear optical absorption of photosynthetic pigment molecules in leaves[J]. Photosynthesis Research, 112(1): 31-37.
蔡永立, 宋永昌. 2000. 中國亞熱帶東部藤本植物的多樣性[J]. 武漢植物研究, 18(5):15-18.
蔡永立, 宋永昌. 2001. 浙江天童常綠闊葉林藤本植物的適應生態學 I.葉片解剖特征的比較[J]. 植物生態學報, 25(1): 90-98.
柴勝豐, 莊雪影, 韋霄, 等. 2013. 光照強度對瀕危植物毛瓣金花茶光合生理特性的影響[J] 西北植物學報, 33(3): 547-554.
陳模舜, 柯世省. 2013. 天臺鵝耳櫪葉片的解剖結構和光合特性對光照的適應[J]. 林業科學, 49(2): 46-53.
陳亞軍, 朱師丹, 曹坤芳. 2008. 兩種光照下木質藤本和樹木幼苗的生理生態學特征[J]. 生態學報, 28(12): 6034-6042.
賀金生, 陳偉烈, 王勛陵. 1994. 高山櫟葉的形態結構及其與生態環境的關系[J]. 植物生態學報, 18(3): 219-227.
季子敬, 全先奎, 王傳寬. 2013. 興安落葉松針葉解剖結構變化及其光合能力對氣候變化的適應性[J]. 生態學報, 33(21): 6967-6974.
江浩, 周國逸, 黃鈺輝, 等. 2011. 南亞熱帶常綠闊葉林林冠不同部位藤本植物的光合生理特征及其對環境因子的適應[J]. 植物生態學報, 35 (5): 567-576.
李芳蘭, 包維楷. 2005. 2011. 植物葉片形態解剖結構對環境變化的響應與適應[J]. 植物學通報, 22(Z): 118-127.
李益鋒, 陶抵輝, 王紹卿. 2011. 藤本植物異軍突起 藤本產業值得開發[J]. 江蘇農業科學, 39(4): 5-8.
龐杰, 張鳳蘭, 郝麗珍, 等. 2013. 沙芥幼苗葉片解剖結構和光合作用對干旱脅迫的響應[J]. 生態環境學報, 22(4): 575-581.
史剛榮, 蔡慶生. 2006. 白三葉葉片解剖可塑性及其對光強的響應[J].草地學報, 14(4): 301-305.
王海艷, 喻理飛. 2010. 不同光環境下3種藤本植物幼苗的光合特性[J].浙江林學院學報, 27(6): 858-864.
王榮, 郭志華. 2007. 不同光環境下楓香幼苗的葉片解剖結構[J]. 生態學雜志, 26(11): 1719-1724.
烏玉娜, 陶建平, 趙科, 等. 2010. 海南霸王嶺天然次生林邊緣效應下木質藤本的變化[J]. 林業科學, 46(5): 1-6.
夏江寶. 2007. 北方主要木質藤本植物光合效率及其對水分與光照的響應[D]. 泰安: 山東農業大學: 1-286.
薛靜,王國驕,李建東,等. 2010. 不同水分條件下三裂葉豚草葉解剖結構的生態適應性[J]. 生態環境學報, 19(3): 686-691.
葉子飄, 于強. 2009. 植物氣孔導度的機理模型[J]. 植物生態學報, 33 (4): 772-782.
袁春明, 劉文耀, 楊國平, 等. 2010. 哀牢山濕性常綠闊葉林木質藤本植物的物種多樣性及其與支柱木的關系. 林業科學, 46(1): 15-22.
袁春明, 劉文耀, 楊國平. 2008. 哀牢山濕性常綠闊葉林林窗木質藤本植物的物種組成與多樣性[J]. 山地學報, 26(1): 29-35.
Response of Leaf Anatomical Structure and Photosynthesis Characteristics of Parthenocissus himalayana to Three Habitat Types
WU Tao1,2, GENG Yunfen1,2, CHAI Yong2, HAO Jiabo1,2, YUAN Chunming1,2*
1. Yunnan Laboratory for Conservation of Rare, Endangered & Endemic Forest Plants, Public Key Laboratory of the State Forestry Administration; Yunnan Provincial Key Laboratory of Cultivation and Exploitation of Forest Plants, Kunming 650201, China; 2. Yunnan Academy of Forestry, Kunming 650201, China
Temporal and spatial changes of habitats of vine plants are often intense, so plants should exhibit great plasticity in order to adapt to heterogeneous habitats. The anatomical structures and photosynthetic characteristics of leaves are considered to greatly reflect the adaptation of plants to different environmental regimes. The objective of the present study was to compare the anatomic and photosynthetic properties of the leaves of liana Parthenocissus himalayana grown in different locations (field, forest edge, and forest interior in the Ailao Mountains, Yunnan, China) with paraffin sectioning and Li-6400 photosynthetic apparatus. The reactions of P. himalayana to different light environments and its survival mechanisms were evaluated. A significant correlation was found between the anatomical structure characteristics and the photosynthesis physiological properties. The intensification of light, from the forest interior to the field, caused a considerable increase in the thickness of the whole leaf and the tissues within upper epidermis, palisade tissue, and palisade cells area, from 157.77 to 299.17 μm, 21.30 to 28.40 μm, 30.83 to 124.65 μm, 430.95 to 652.97 μm2, respectively. There was also a significant rise in the length (from 29.23 to 49.54 μm) and circumference of palisade cell (from 86.58 to 155.17 μm). The thickness of the lower epidermis and the stomatal size and density were substantially increased. There were no significant differences in width of palisade cells (19.67-22.81 μm) among different habitats. The average value of plasticity indexes of leaf anatomical characteristics was 0.37, among which the length of palisade cells had the maximum value (0.67) and the size of stoma had the minimum one (0.11). The light saturation point (LSP), light compensation point (LCP), and the maximum photosynthetic rate under saturation light intensity (Pmax) were markedly elevated. Higher were also the values of a number of indicators, including the dark respiration rate (Rd), CO2compensation point (Ccp), light respiration rate (Rp), and initial carboxylation efficiency (CE). These results indicated that Parthenocissus himalayana have a remarkable light adaptability in heterogeneous habitats in terms of leaf anatomical structure and photosynthetic characteristics.
Parthenocissus himalayana; leaf; anatomical structure; photosynthesis characteristics; habitat
Q945.79
A
1674-5906(2014)10-1586-07
吳濤,耿云芬,柴勇,郝佳波,袁春明. 三葉爬山虎葉片解剖結構和光合生理特性對3種生境的響應[J]. 生態環境學報, 2014, 23(10): 1586-1592.
WU Tao, GENG Yunfen, CHAI Yong, HAO Jiabo, YUAN Chunming. Response of leaf anatomical structure and photosynthesis characteristics of Parthenocissus himalayana to three habitat types [J]. Ecology and Environmental Sciences, 2014, 23(10): 1586-1592.
國家自然科學基金項目(31160136)
吳濤(1979年生),男,助理研究員,博士,主要從事林木遺傳育種和植物生理學研究。E-mail:ynafwt@126.com
*通信作者:袁春明(1964年生),男,副研究員,博士,主要從事植物生態、恢復生態和保護生物學等方面的研究。E-mail:yuanchunming1017@163.com
2014-09-09
