基于mtCOII基因對山東省越冬代亞洲玉米螟不同種群的遺傳結構分析
2013-12-25 11:15:43李麗莉陶云荔
生態學報 2013年14期
李麗莉,于 毅,國 棟,陶云荔,褚 棟,*
(1.山東省農業科學院植保所,山東省植物病毒學重點實驗室,濟南 250100;2.青島農業大學農學與植物保護學院,山東省植物病蟲害綜合防控重點實驗室,青島 266109)
亞洲玉米螟Ostriniafurnacalis(Guenée)是我國玉米上的重要害蟲,它可在玉米的不同生育期危害,嚴重影響玉米的產量和質量[1]。該害蟲具有較廣的地理分布[2]和寄主范圍[3]。近年來,隨著全球氣候變暖以及玉米種植面積的擴大,其發生危害有不斷加重的趨勢[4]。
亞洲玉米螟不同種群遺傳結構分析對于該害蟲的演化、遷飛以及綜合治理等具有重要的意義[2]。目前,分子標記廣泛應用于物種的遺傳分化研究中[5]。線粒體COII基因(mtCOII)在昆蟲種群遺傳學研究中常作為分析地理種群系統發育關系的分子工具[6]。前人利用mtCOII基因分別對日本與中國的亞洲玉米螟不同種群進行遺傳分化分析的結果表明,日本與中國的亞洲玉米螟種群分別具有2個遺傳支系[2,7]。其中,李菁等[2]對1個山東省種群進行了遺傳分化分析,結果表明屬于其中1個支系。山東省是我國玉米的重要產區之一,又是黃淮海地區的主要糧食生產地,所處的地理位置特殊,種植結構復雜多樣,全省各地亞洲玉米螟的發生危害程度也有很大的不同[8]。因此,有必要進一步擴大采集范圍,全面揭示山東省越冬代亞洲玉米螟種群的遺傳分化。此外,中國、日本的亞洲玉米螟種群分化支系[2,7]之間的關系尚未清楚,山東省亞洲玉米螟種群與上述結果中不同分化種群[2,7]之間的關系尚需進一步解析。不同的分化種群往往對環境具有不同的適應能力[9-10],因此,明確山東省不同地理種群的亞洲玉米螟遺傳分化地位對于該害蟲的綜合防控具有重要的理論意義與實踐價值,并將促進我國亞洲玉米螟分子系統學的深入研究。
為了全面揭示亞洲玉米螟尤其是山東省亞洲玉米螟不同種群的遺傳結構,本研究對山東省9個地區17個亞洲玉米螟種群的mtCOII序列進行了測序分析,獲得126條mtCOII基因序列;結合GenBank上所有國內外亞洲玉米螟已有mtCOII序列,進一步解析了國內外亞洲玉米螟不同地理種群的系統進化關系;分析了山東省不同地理種群的遺傳多樣性;構建了利用mtCOII基因PCR-RFLP標記來快速鑒別亞洲玉米螟遺傳支系的方法。
1 材料與方法
1.1 供試昆蟲
亞洲玉米螟種群樣品于2004年10—11月采集自山東省9個地區的17個地點,采集帶蟲的玉米秸稈、穗軸放置室外自然條件下越冬,待第2年化蛹后取單頭蛹置于離心管中,-20 ℃保存至DNA提取。詳細情況見表1。
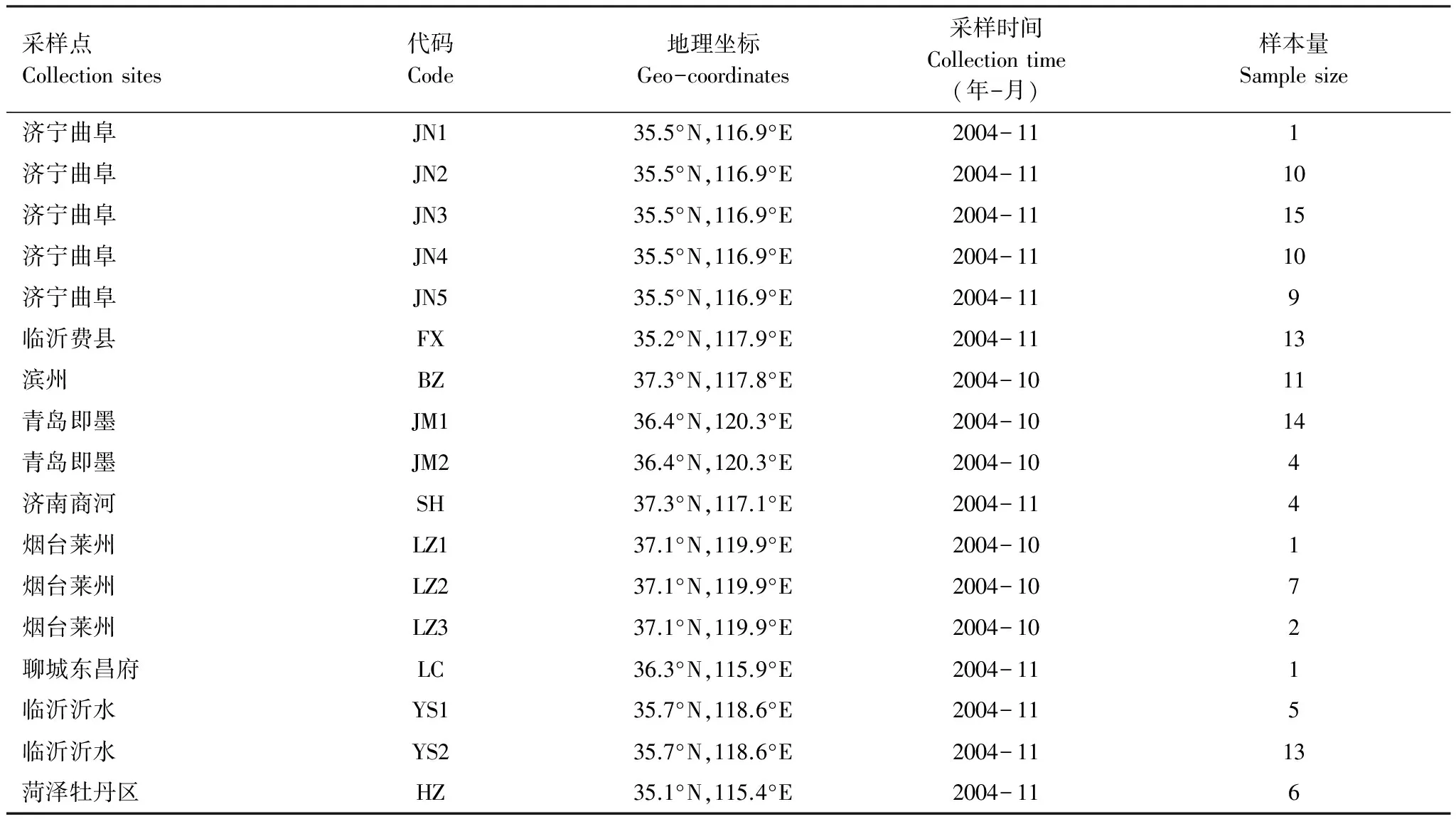
表1 山東省亞洲玉米螟不同種群樣品的采集情況
1.2 單頭玉米螟DNA提取、PCR反應條件及電泳檢測
單頭玉米螟DNA提取采用DNA提取試劑盒(天根生化科技),具體步驟按說明書進行。mtCOIIPCR反應體系及反應程序參照李菁等[2]。取5 μL PCR產物經1.0%瓊脂糖凝膠(含EB)電泳后,紫外燈下成像檢測。選取有條帶的個體進行5′端單向測序。
1.3 數據處理與統計分析
利用MEGA5.05[11]軟件將測序得到的序列進行多序列同源比對。采用軟件DnaSP 5.0[12]分析了單倍體型的數量,計算了山東省亞洲玉米螟不同采集種群(個體數量不少于3個)的多樣性指數:多態位點的個數(S);突變位點(η);單倍體型數量(H);核苷酸多態性(π);平均核苷酸差異(K);Jukes和Cantor校正π值(π(JC))。利用MEGA5.05[11]軟件,采用鄰接法(NJ)以家蠶和桃小食心蟲的mtCOII基因(GenBank登錄號分別為AB083339、HM156482)為外群,分別以最大似然法(ML)與鄰位法(NJ)構建所有亞洲玉米螟mtCOII基因單倍體型的系統樹。利用Arlequin3.11[13]軟件進行分子變異(AMOVA)分析。
1.4 亞洲玉米螟2個支系mtCOII酶切位點分析以及PCR-RFLP鑒別方法構建
利用酶切位點在線查詢(http://watcut.uwaterloo.ca/watcut/watcut/template.php?act= restriction _ new)對2個支系所有單倍體型進行了酶切位點分析。取5 μL的PCR產物加入1 μL內切酶PspFI(酶切位點為CCCAGC)以及2 μL的10×Buffer,用雙蒸水補足至20 μL。37 ℃消化5 min,80 ℃反應5 min終止反應,取5 μL酶切產物經1.5%瓊脂糖凝膠(含EB),100 V電壓電泳20 min后紫外燈下成像拍照。
2 結果與分析
2.1 亞洲玉米螟mtCOII基因單倍體型分析
對GenBank檢索以及本研究測序獲得的mtCOII進行比對處理后,片段長度為656 bp。通過GenBank檢索獲得亞洲玉米螟的mtCOII基因214條(截止2011年10月1日數據)。對上述214條序列通過DnaSP 5.0[12]分析得到56條單倍體型(編號為Hap1—Hap56)(表2)。本實驗中獲得126條mtCOII基因的部分序列,共有20個單倍體型,其中14個單倍體型能夠在GenBank中發現相應序列;有6種單倍體型為山東種群所特有(表3)。
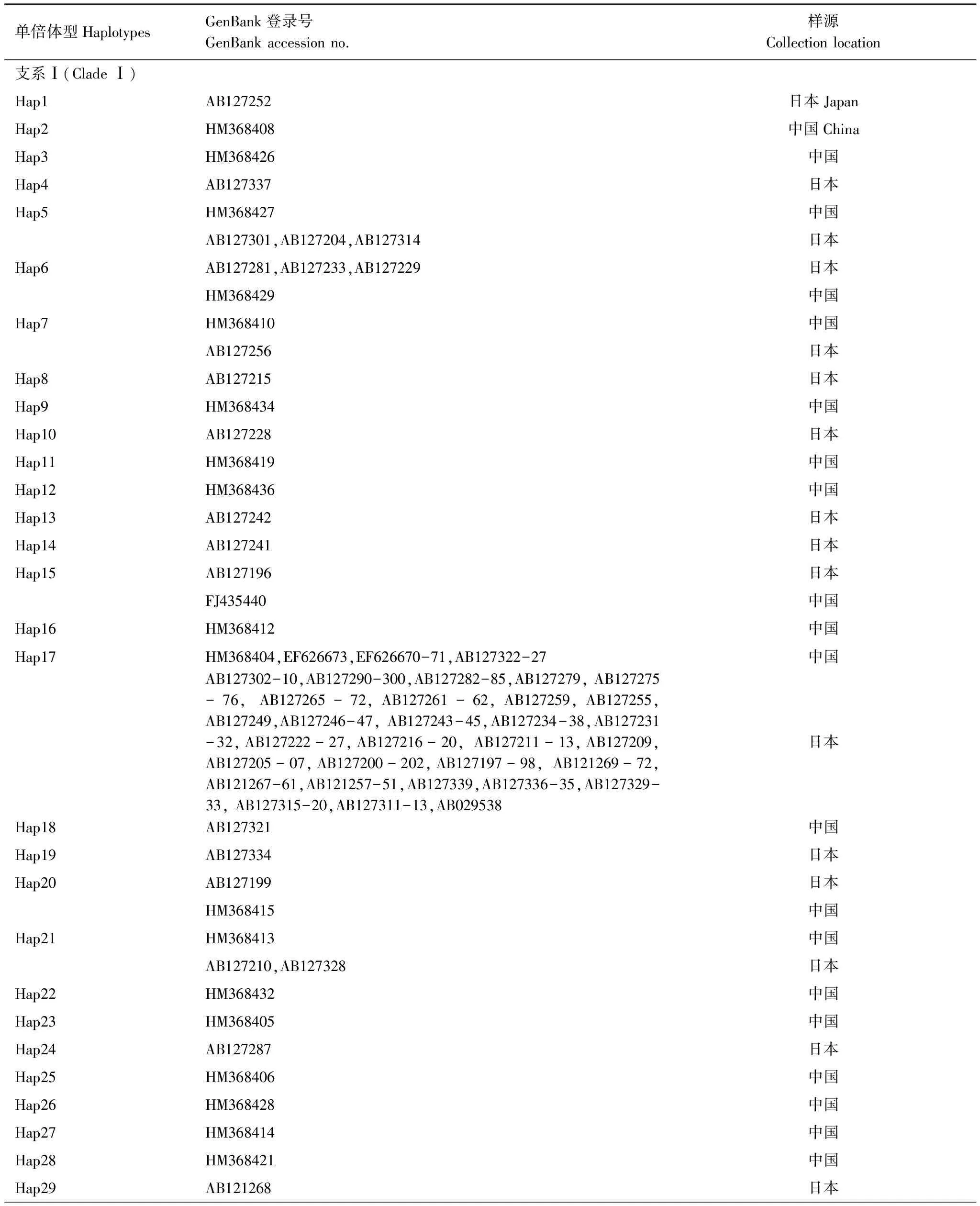
表2 GenBank中亞洲玉米螟mtCOII單倍體型分布情況
續表
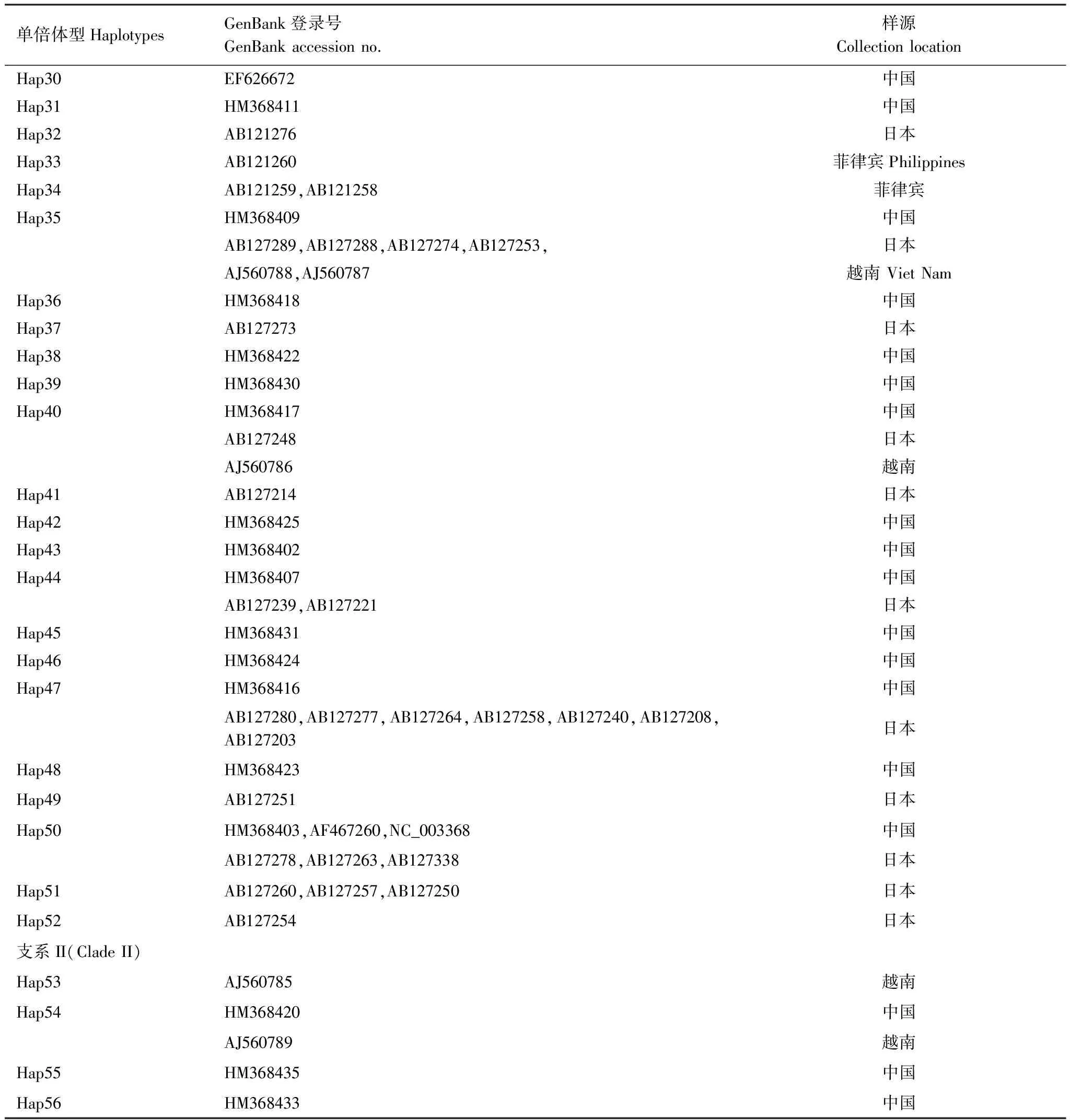
單倍體型HaplotypesGenBank登錄號GenBankaccessionno.樣源CollectionlocationHap30EF626672中國Hap31HM368411中國Hap32AB121276日本Hap33AB121260菲律賓PhilippinesHap34AB121259,AB121258菲律賓Hap35HM368409中國AB127289,AB127288,AB127274,AB127253,日本AJ560788,AJ560787越南VietNamHap36HM368418中國Hap37AB127273日本Hap38HM368422中國Hap39HM368430中國Hap40HM368417中國AB127248日本AJ560786越南Hap41AB127214日本Hap42HM368425中國Hap43HM368402中國Hap44HM368407中國AB127239,AB127221日本Hap45HM368431中國Hap46HM368424中國Hap47HM368416中國AB127280,AB127277,AB127264,AB127258,AB127240,AB127208,AB127203日本Hap48HM368423中國Hap49AB127251日本Hap50HM368403,AF467260,NC_003368中國AB127278,AB127263,AB127338日本Hap51AB127260,AB127257,AB127250日本Hap52AB127254日本支系II(CladeII)Hap53AJ560785越南Hap54HM368420中國AJ560789越南Hap55HM368435中國Hap56HM368433中國
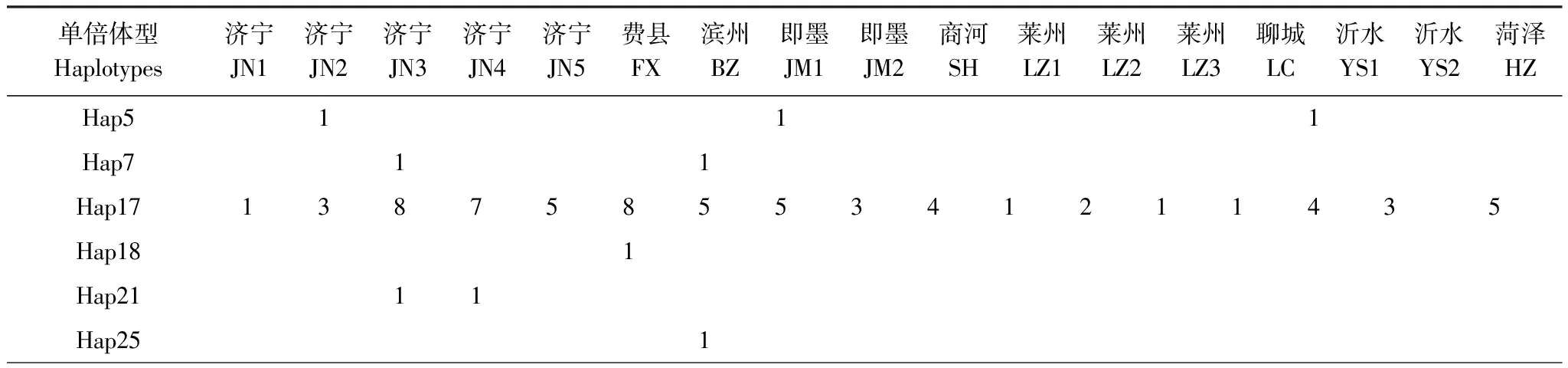
表3 本研究中山東省亞洲玉米螟mtCOII單倍體型分布情況
續表
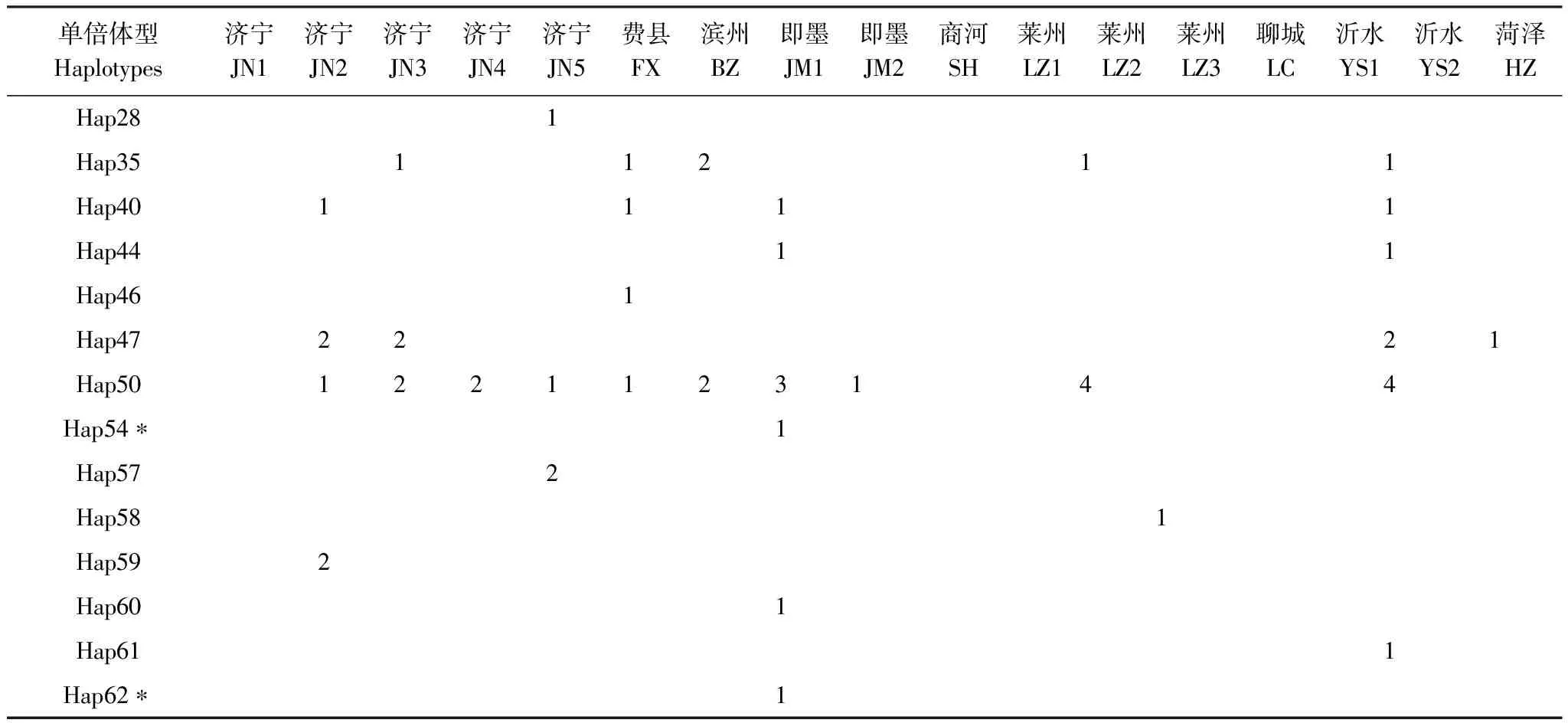
單倍體型Haplotypes濟寧JN1濟寧JN2濟寧JN3濟寧JN4濟寧JN5費縣FX濱州BZ即墨JM1即墨JM2商河SH萊州LZ1萊州LZ2萊州LZ3聊城LC沂水YS1沂水YS2菏澤HZHap281Hap3511211Hap401111Hap4411Hap461Hap472221Hap501221123144Hap54?1Hap572Hap581Hap592Hap601Hap611Hap62?1
*屬于支系Ⅱ;Grouped into Clade Ⅱ
2.2 mtCOII基因單倍體型系統進化關系分析
對上述亞洲玉米螟mtCOII基因62種單倍體型以最大似然法(ML)構建的系統樹(圖1;僅顯示置信值(Bootstrap)大于50%的分支)表明,所有單倍體型共分為2個支系。其中,57個單倍體型單獨聚為一支(置信值為58%);5個單倍體型單獨聚為一支(置信值為95%)。鄰位法(NJ)(結果未顯示)構建的系統樹與最大似然法(ML)構建的系統樹類似,也分為2個支系。
本實驗中只有山東省的JM1種群(即墨)中的2個個體(分別編號為Hap54、Hap62單倍體型)位于第2支系(Clade Ⅱ),山東其他種群均屬于第1支系(Clade Ⅰ)。
2.3 山東省亞洲玉米螟不同地理種群遺傳多樣性分析
基于mtCOII基因對山東省不同種群遺傳多樣性分析如表4所示。由表4可以看出,所測的玉米螟種群的遺傳多樣性均較高,例如總體單倍體型多樣性指數Hd為0.695;種群內單倍體型多樣性指數為0.333—0.889,平均為0.611,其中單倍體型多樣度最高的種群為JN2(濟寧)種群,最低的為HZ(菏澤)種群。AMOVA分析結果顯示(表5),山東省亞洲玉米螟2個支系之間的方差組分為4.78994,占方差比率的79.42%;2個支系之間的方差組分為1.24116,占方差比率的20.58%;這說明山東亞洲玉米螟的遺傳變異主要來自2個支系之間,支系間的遺傳差異達到極顯著水平(P<0.0001)。
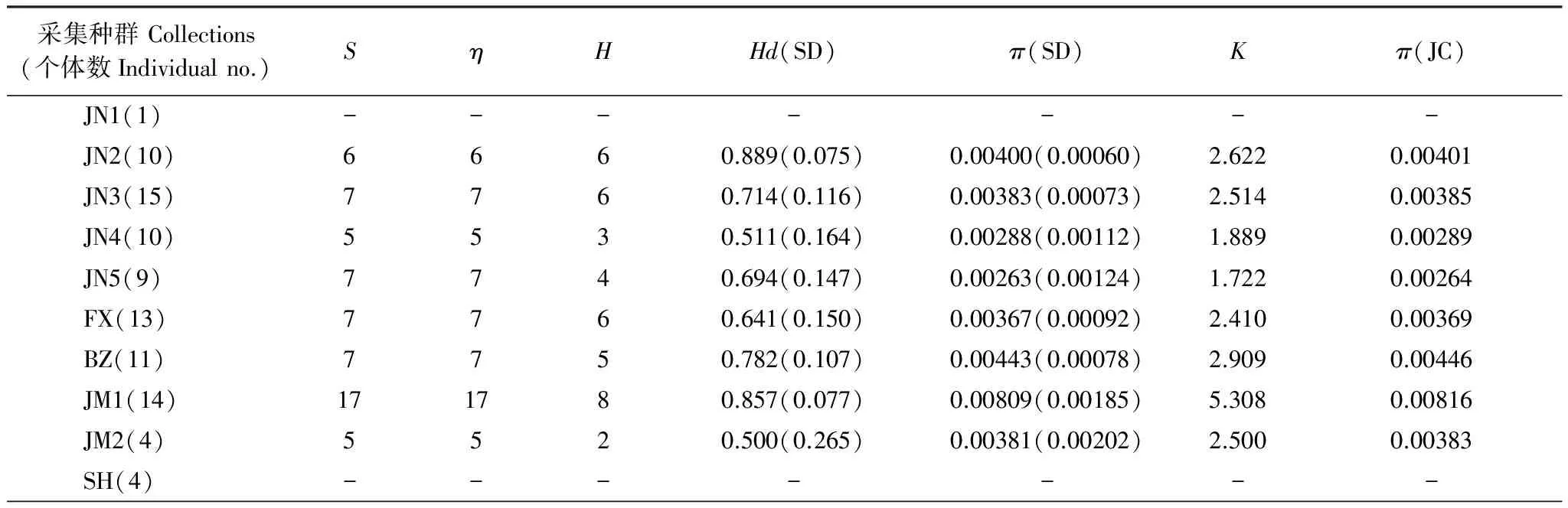
表4 基于mtCOII序列對山東省亞洲玉米螟不同種群的遺傳多樣性分析
續表
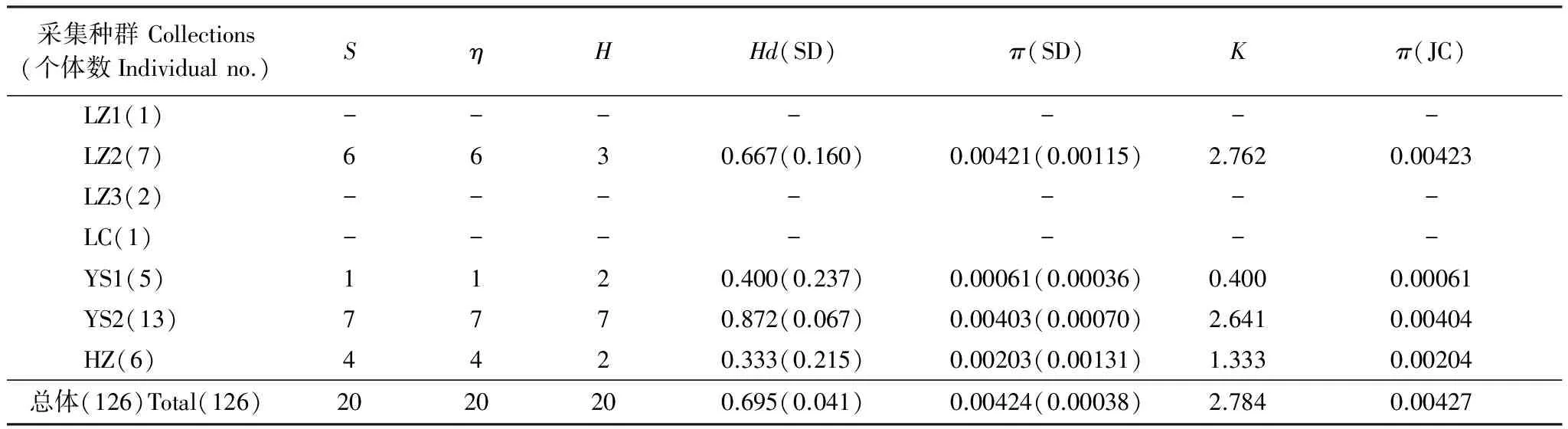
采集種群Collections(個體數Individualno.)SηHHd(SD)π(SD)Kπ(JC)LZ1(1)-------LZ2(7)6630.667(0.160)0.00421(0.00115)2.7620.00423LZ3(2)-------LC(1)-------YS1(5)1120.400(0.237)0.00061(0.00036)0.4000.00061YS2(13)7770.872(0.067)0.00403(0.00070)2.6410.00404HZ(6)4420.333(0.215)0.00203(0.00131)1.3330.00204總體(126)Total(126)2020200.695(0.041)0.00424(0.00038)2.7840.00427
S: 多態位點的個數number of polymorphic(segregating)sites;η: 總的突變位點total number of mutations;H: 單倍體型數量number of haplotypes;Hd: 單倍體型多態性Haplotype diversity;π: 核酸多樣性nucleotide diversity;K: 平均核苷酸差異average number of nucleotide differences;π(JC): Jukes 和 Cantor校正的π值;-:未計算

圖1 利用最大似然法(ML)構建的亞洲玉米螟不同地理種群COII單倍體型間的系統樹
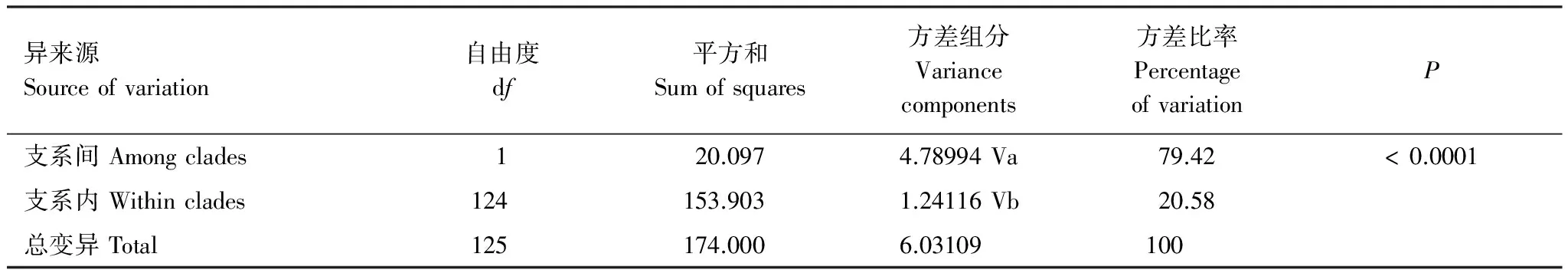
表5 亞洲玉米螟不同支系線粒體基因COII的分子變異分析
固定系數 Fixation index,Fst: 0.79421
2.4 基于PCR-RFLP方法鑒別亞洲玉米螟2個支系

圖2 亞洲玉米螟支系Ⅱ的mtCOII基因序列中PspFI酶切位點位置
利用酶切位點在線查詢的方法,發現PspFI內切酶(酶切位點為CCCAGC)僅存在于支系Ⅱ中,而支系Ⅰ中不存在該酶切位點(圖2)。由AF467260、NC_003368可知,PCR產物片段長度824 bp,酶切位點位于320 bp左右,因此可以推測酶切片段為500 bp與300 bp左右。本研究表明通過PspFI內切酶對山東省支系Ⅰ及支系Ⅱ亞洲玉米螟擴增mtCOII基因PCR產物酶切顯示,支系Ⅱ中JM1種群的2個個體酶切后有3條帶(分別約800 bp、500 bp、300 bp),其他支系I種群個體只有1條帶(約800 bp)(圖3)。
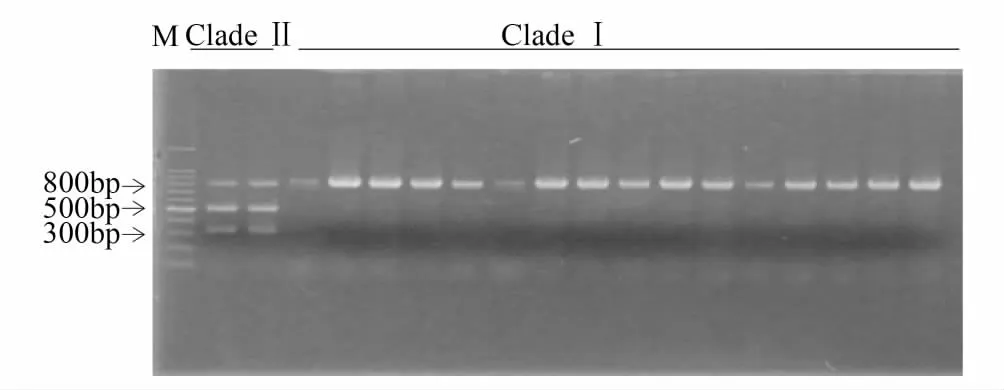
圖3 亞洲玉米螟支系Ⅰ與Ⅱ的mtCOII酶切結果
3 討論
Hoshizaki等[7]基于mtCOII基因發現日本的亞洲玉米螟分為2個支系(支系A和B),其中支系A在日本分布廣泛,占絕對優勢,在對日本16個采樣種群檢測中有15個點均含有支系A;在被檢測的中國樣品中僅發現了支系A;在菲律賓僅發現支系B個體。李菁等[2]利用mtCOII基因發現中國17個地理種群的亞洲玉米螟也存在2個較大的支系。本研究對目前全球亞洲玉米螟分析結果表明:全球的亞洲玉米螟具有2個遺傳支系(Clade Ⅰ和Clade Ⅱ),與李菁等[2]結果是一致的;而Hoshizaki等[7]中的支系A和B均在支系Ⅰ(Clade Ⅰ)中,沒有明顯的遺傳分化。近來研究表明,很多物種存在遺傳分化,這些分化種群往往對環境的適應能力不同[9-10]。例如Brunner等[14]研究發現,西花薊馬2個遺傳支系(G型和L型)對生存環境的適應性存在很大的差異,G型常生活于高溫干燥(hot-dry)地區,L型大都生活在低溫潮濕(cool-moist)地區。李菁等[2]與本文的研究表明,亞洲玉米螟支系Ⅱ的個體比例較低,這些數據提示不同支系內亞洲玉米螟的生物學、生態學及生理學方面可能存在差異,進而影響該害蟲種群的地理分布及其擴散。這些問題有待于進一步研究,相關研究結果對于該害蟲的可持續控制具有重要的指導價值。
李菁等[2]研究中分析的山東省亞洲玉米螟種群(1個種群)屬于支系Ⅰ(Clade Ⅰ)。本研究中發現山東省亞洲玉米螟單倍體型分布于支系Ⅰ與支系Ⅱ。這表明,樣品的數量增加可以發現更多的具有遺傳分化的個體。盡管如此,本研究中發現的支系Ⅱ個體在山東省所有樣品中的比例很低(1.9%)。同時,這些單倍體型在山東省的分布缺乏明顯的地理分布格局,與全國分布格局類似[2]。本研究發現,山東省亞洲玉米螟mtCOII基因存在豐富的單倍體型(20個);在山東省,亞洲玉米螟種群Hd(0.333—0.889)與全國其他種群Hd(0.424—0.862)相似[2]。山東省總群體的單倍體型多樣性指數Hd為0.695,略低于全國水平(0.811)[2]。上述結果表明山東省亞洲玉米螟種群線粒體基因具有較高的多態性。其中,JM1(即墨)種群具有較高的遺傳多樣性,具有多種可能性,尚需進一步的分析。
PspFI內切酶(酶切位點為CCCAGC)對支系Ⅰ與Ⅱ的亞洲玉米螟mtCOII基因的PCR產物酶切結果表明,該酶能夠有效鑒別2個支系:支系Ⅱ(JM1種群的2個個體)有3條帶(分別約800 bp、500 bp、300 bp),支系Ⅰ個體只有1條帶(約800 bp)。然而根據序列分析,支系Ⅱ(JM1種群的2個個體)PCR產物應該被切成2條帶(分別約為500 bp、300 bp),而本研究中尚有PCR產物(約800 bp)不能完全切開,其原因有待于進一步分析。盡管如此,利用PspFI內切酶消化mtCOII基因的PCR-RFLP快速鑒別技術為亞洲玉米螟2支系的生物學及生態學進一步研究奠定了基礎。
致謝: 美國斯坦福大學Daniel Thomas Rogan幫助修改英文摘要,特此致謝。
[1]Zhou D Y, He K L. Asian Corn Borer and Its Integrated Management. Beijing: Golden Shield Press, 1995: 1-102.
[2]Li J, Zhang Y, Wang Z Y, He K L, Wang Q. Genetic differentiation and gene flow among different geographical populations of the Asian corn borerOstriniafurnacalis(Guenée) (Lepidoptera Crambidae) in China estimated by mitochondrialCOIIgene sequences. Acta Entomologica Sinica, 2010, 53(10): 1135-1143.
[3]Wang Z Y, Lu X, He K L, Zhou D R. Review of history, precent situation and prospect of the Asian maize borer research in China. Journal of Shenyang Agricultural University, 2000, 31(5): 402-412.
[4]The Ministry of Agriculture. Control scheme of corn borer in 2008. Rural Practical Technology, 2008, (5): 1-37.
[5]Chu D, Zhang Y J, Wan F H. Application of molecular marker techniques in invasion ecology. Chinese Journal of Applied Ecology, 2007, 18(6): 1383-1387.
[6]Bu Y, Zheng Z M. Roles and status of cytochrome oxidase II gene in the research of insect molecular systematics. Entomological Knowledge, 2005, 42(1): 18-22.
[7]Hoshizaki S, Washimori R, Kubota S, Ohno S, Huang Y, Tatsuki S, Ishikawa Y. Two mitochondrial lineages occur in the Asian corn borer,Ostriniafurnacalis(Lepidoptera: Crambidae), in Japan. Bulletin of Entomological Research, 2008, 98(5): 519-526.
[8]Li L L, Yu Y, Zhang A S, Zhang S C, Men X Y, Zhang J J. Investigation on the species of overwintering maize pests and parasitoids of over wintering Asian corn borer larvae in Shandong Province. Joumal of Shandong Agricultural Science, 2009, (2): 75-77.
[9]Scheffer S J, Lewis M L. Mitochondrial phylogeography of vegetable pestLiriomyzasativae(Diptera: Agromyzidae): divergent clades and invasive populations. Annals of the Entomological Society of America, 2005, 98(2): 181-186.
[10]Winkler G, Dodson J J, Lee C E. Heterogeneity within the native range: population genetic analyses of sympatric invasive and noninvasive populations of the freshwater invading copepodEurytemoraaffinis. Molecular Ecology, 2008, 17(1): 415-430.
[11]Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 2011. doi: 10.1093/molbev/msr121.
[12]Librado P, Rozas J. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics, 2009, 25(11): 1451-1452.
[13]Excoffier LG, Laval S, Schneider S. Arlequin ver 3.0: an integrated software package for population genetics data analysis. Evol Bioinformatics, 2005, 1:47-50.
[14]Brunner P C, Frey J E. Habitat-specific population structure in native western flower thripsFrankliniellaoccidentalis(Insecta, Thysanoptera). Journal of Evolutionary Biology, 2010, 23(4): 797-804.
參考文獻:
[1]周大榮, 何康來. 玉米螟綜合防治. 北京: 金盾出版社, 1995: 1-102.
[2]李菁, 張穎, 王振營, 何康來, 王強. 基于線粒體DNACOII基因的亞洲玉米螟中國不同地理種群遺傳分化及基因流研究. 昆蟲學報, 2010, 53(10): 1135-1143.
[3]王振營, 魯新, 何康來, 周大榮. 我國研究亞洲玉米螟歷史、現狀與展望. 沈陽農業大學學報, 2000, 31(5): 402-412.
[4]農業部. 2008年玉米螟防治方案. 農村實用技術, 2008, (5): 1-37.
[5]褚棟, 張友軍, 萬方浩. 分子標記技術在入侵生態學研究中的應用. 應用生態學報, 2007, 18(6): 1383-1387.
[6]卜云, 鄭哲民. COII基因在昆蟲分子系統學研究中的作用和地位. 昆蟲知識, 2005, 42(1): 18-22.
[8]李麗莉, 于毅, 張安盛, 張思聰. 山東省玉米越冬害蟲種類及亞洲玉米螟越冬幼蟲寄生性天敵調查. 山東農業科學, 2009, (2): 75-77.
猜你喜歡
山東交通科技(2022年3期)2022-08-05 03:06:24
中國集體經濟(2022年9期)2022-04-12 01:07:35
雜技與魔術(2022年1期)2022-03-16 10:22:48
——山東省濟寧市老年大學之歌
老年教育(老年大學)(2021年10期)2021-11-12 09:15:48
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
新生代(2018年23期)2018-11-13 22:41:10
中國學校體育(2016年11期)2016-12-01 10:42:48
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
