牡蠣活性肽的分離及其免疫抑制作用的實(shí)驗(yàn)研究
2013-12-23 05:13:34李超柱陳艷華陳艷輝張艷秋方懷義
海洋科學(xué) 2013年4期
關(guān)鍵詞:小鼠
李超柱, 陳艷華, 陳艷輝, 張艷秋, 方懷義
(1. 欽州學(xué)院, 廣西 欽州 535000; 2. 廣西醫(yī)科大學(xué) 附屬腫瘤醫(yī)院, 廣西 南寧 530021)
牡蠣(Oyster )俗稱(chēng)大蠔、蠣黃、蠣子、海蠣等, 不僅是一種肉嫩、味鮮營(yíng)養(yǎng)價(jià)值極高的海產(chǎn)食品, 也是一種很有藥用價(jià)值的海洋生物, 近些年來(lái)受到了國(guó)內(nèi)外學(xué)者的重視。不少學(xué)者報(bào)道了牡蠣提取物具有抗腫瘤作用、抗氧化、對(duì)腫瘤細(xì)胞的放射增敏、清除氧自由基以及對(duì)動(dòng)物機(jī)體的免疫調(diào)節(jié)作用等[1-7], 但對(duì)于牡蠣活性肽是否具有免疫調(diào)節(jié)作用, 尤其是免疫抑制作用仍鮮見(jiàn)報(bào)道。
蛋白質(zhì)是一類(lèi)最重要的大分子物質(zhì), 具有很重要的生理功能, 天然蛋白質(zhì)不完全水解產(chǎn)生的長(zhǎng)短不一的一些肽段稱(chēng)為小分子多肽, 小分子多肽中具有生物活性的肽稱(chēng)為活性肽。生物來(lái)源于海洋的活性物質(zhì)由于特殊生長(zhǎng)環(huán)境, 因而具有許多獨(dú)特的性質(zhì), 如結(jié)構(gòu)特異、活性強(qiáng)、含量低微、毒副作用小等特點(diǎn)[8-9]。本研究利用動(dòng)物蛋白水解酶水解牡蠣肉, 并對(duì)最佳水解產(chǎn)物進(jìn)行柱層析和超濾分離獲得一種低分子質(zhì)量(約為3 kD)活性肽, 并以小鼠脾淋巴細(xì)胞為實(shí)驗(yàn)?zāi)P? 通過(guò)淋巴細(xì)胞增殖實(shí)驗(yàn)和混合淋巴細(xì)胞實(shí)驗(yàn), 從細(xì)胞、分子水平研究牡蠣活性肽對(duì)小鼠脾淋巴細(xì)胞的免疫抑制活性及作用機(jī)制。
1 材料、試劑與儀器
1.1 材料、試劑
新鮮牡蠣, 采自廣西北部灣茅尾海海域, 去殼、清洗并勻漿, 置-20℃儲(chǔ)存?zhèn)溆? 動(dòng)物蛋白酶購(gòu)自廣西龐博生物有限公司(≥200 u/mg )。
SPF級(jí)BALB/c小鼠和昆明小鼠由廣西醫(yī)科大學(xué)實(shí)驗(yàn)動(dòng)物中心提供, 雄性, 體重22~24 g, 周齡6~8周。
RPMI 1640培養(yǎng)基(賽默飛世爾生物化學(xué)制品(北京)有限公司); 小牛血清(杭州四季青生物材料研究所); 磷酸鹽緩沖液(PBS); 淋巴細(xì)胞分離液; 絲裂霉素(惠州鴻雨科技有限公司); MTT(sigma公司); 伴刀豆蛋白A(ConA)(Sigma公司, 用雙蒸水配成2mg/mL的溶液, 過(guò)濾除菌)。
1.2 主要儀器
低速離心機(jī)(北京醫(yī)用離心機(jī)廠); 低溫孔板離心機(jī)(sigma公司); CO2培養(yǎng)箱(Thermo 公司), 酶聯(lián)免疫檢測(cè)儀( Thermo(上海)儀器有限公司); 倒置光學(xué)顯微鏡(Nikon中國(guó)有限公司); 超凈工作臺(tái)( 蘇州安泰空氣技術(shù)有限公司)。
2 方法
2.1 牡蠣蛋白活性肽的制備
原料蛋白—預(yù)處理—選擇最佳酶解條件水解—高溫滅酶—活性炭脫苦味去色—柱層析—超濾膜分離—冷凍干燥—組成與結(jié)構(gòu)鑒定[10]。
將水解產(chǎn)物分子質(zhì)量小于3 kD的活性肽配置成250、125、62.5、31.6、15.6、7.8 mg/L的濃度梯度。
2.2 小鼠淋巴細(xì)胞增殖實(shí)驗(yàn)
小鼠脾淋巴細(xì)胞懸液的制備及原代培養(yǎng)方法, 頸椎脫臼處死BALB/c小鼠75%酒精消毒小鼠皮膚, 著重腹部消毒, 取出脾臟, 置有PBS緩沖液培養(yǎng)皿中撕脾, 經(jīng)200目濾網(wǎng)過(guò)濾, 制備脾淋巴細(xì)胞懸液(冰上進(jìn)行), 離心(2 000 r/min, 5 min), 棄上清液, 將脾細(xì)胞懸液體積按1 : 1的比例加入預(yù)裝有等量淋巴細(xì)胞分離液的離心管中, 離心30 min(2 000 r/min)。離心完畢吸取界面層單個(gè)核細(xì)胞(白細(xì)胞層), 再次用PBS緩沖液沖洗離心5 min(500 r/min), 棄上清液, 用含10%FCS的PRMI-1640培養(yǎng)基重懸細(xì)胞, 調(diào)整細(xì)胞濃度至1×106個(gè)/mL。
將細(xì)胞調(diào)好濃度后, 加入96孔板, 每孔100 μL, 用含10%胎牛血清的PRMI-1640培養(yǎng)基將ConA濃度配制為20 mg/L(終濃度為10 mg/L ), 加入干預(yù)活性肽的終濃度分別為250、125、62.5、31.6、15.6、7.8 mg/L, 每孔100 μL; 每組平行做3個(gè)復(fù)孔, 并設(shè)淋巴細(xì)胞+ConA空白對(duì)照3個(gè)復(fù)孔。
96孔板置于CO2培養(yǎng)箱培養(yǎng)48 h, 加入5 g/LMTT, 每孔20 μL。4 h后低溫孔板離心機(jī)(3 000 r/min,4℃)離心, 10 min; 棄上清液, 加0.4 mol/L的鹽酸化異丙醇, 每孔100 μL; 待結(jié)晶物溶解后, 用酶標(biāo)儀于492nm處測(cè)定吸光度。
2.3 混合淋巴細(xì)胞實(shí)驗(yàn)
取BALB/c鼠脾淋巴細(xì)胞懸液稱(chēng)為反應(yīng)細(xì)胞(A)。取適量50 mg/L絲裂霉素加入昆明鼠脾淋巴細(xì)胞懸液內(nèi)至終濃度25 mg/L, 37℃保溫30 min, PBS液洗兩遍, 用含小牛血清10%的RPMI-1640懸液細(xì)胞, 計(jì)數(shù), 配成一定濃度細(xì)胞, 稱(chēng)為絲裂霉素處理的刺激細(xì)胞(B)。將上述制備好的兩種淋巴細(xì)胞按1 : 1的體積加入培養(yǎng)皿混合加96孔板, 對(duì)照組為100 μL 培養(yǎng)基+100 μL混合脾淋巴細(xì)胞, 加藥組為100 μL不同濃度的干預(yù)活性肽 + 100 μL混和脾淋巴細(xì)胞。在37℃、5% CO2培養(yǎng)箱中孵育48 h, 于培養(yǎng)結(jié)束前4 h加入5 g/L MTT, 每孔20 μL, 培養(yǎng)結(jié)束顯微鏡下觀察其MTT結(jié)晶情況。然后用低溫孔板離心機(jī)(3000 r/min,4℃)離心, 10 min; 棄上清, 加0.4 mol/L的鹽酸化異丙醇, 每孔100 μL; 待結(jié)晶物溶解后, 用酶標(biāo)儀于492 nm處測(cè)定吸光度。
2.4 ELISA 方法對(duì)體外培養(yǎng)的細(xì)胞進(jìn)行細(xì)胞因子產(chǎn)生能力檢測(cè)
刀豆蛋白(ConA)激活的小鼠脾臟淋巴細(xì)胞, 經(jīng)過(guò)活性肽作用后, 以 ELISA 方法檢測(cè)細(xì)胞培養(yǎng)上清液中細(xì)胞因子所分泌的細(xì)胞因子 IL-2和 IFN-γ的變化。首先, 原代培養(yǎng)小鼠脾淋巴細(xì)胞, 調(diào)節(jié)細(xì)胞懸液(2×106個(gè)/mL)。加入 24 孔培養(yǎng)板中, 1 mL/孔, 置 37℃、5%CO2培養(yǎng)箱培養(yǎng) 4 h, 加入 0.5 mL ConA(終濃度 5 mg/L)以及不同濃度的牡蠣活性肽, 牡蠣活性肽的濃度分別為低劑量 15.6 mg/L, 中劑量 62.5 mg/L, 高劑量 125 mg/L各 0.5 mL, 置37 , ℃5%CO2培養(yǎng) 48 h, 收集上清, -20℃保存待測(cè)。隨后進(jìn)行 IL-2 和 IFN-γ 的ELISA 檢測(cè)。
2.5 統(tǒng)計(jì)分析方法
采用SPSS17.0 統(tǒng)計(jì)軟件包中多個(gè)樣本率比較的方法進(jìn)行統(tǒng)計(jì)學(xué)分析, 計(jì)量數(shù)據(jù)用均數(shù)±標(biāo)準(zhǔn)差(x±s)表示, 以P<0.05為差異有統(tǒng)計(jì)學(xué)意義。
3 結(jié)果與討論
3.1 對(duì)小鼠脾淋巴細(xì)胞增殖和混合淋巴細(xì)胞生長(zhǎng)的影響
從表1 可以看出, 牡蠣活性肽對(duì)小鼠脾淋巴細(xì)胞的生長(zhǎng)抑制率出現(xiàn)明顯的量效關(guān)系, 即在一定濃度范圍內(nèi), 隨著濃度的升高, 抑制率增大。其IC50為125 mg/L。
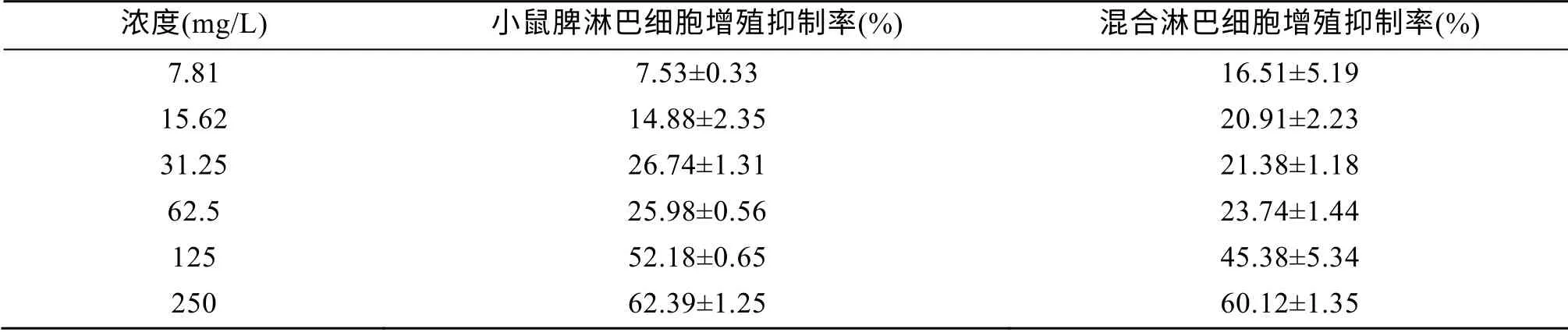
表1 牡蠣活性肽對(duì)淋巴細(xì)胞的生長(zhǎng)抑制率(n =3, x±s) Tab. 1 Inhibition of lymphocyte proliferation by oyster peptides (n = 3, x ± s)
免疫抑制劑通過(guò)抑制免疫細(xì)胞(T細(xì)胞和B細(xì)胞等巨噬細(xì)胞)的增殖和功能, 降低抗體免疫反應(yīng)。在特異性抗原刺激下可以使相應(yīng)的淋巴細(xì)胞克隆發(fā)生增殖。如刀豆蛋白ConA是T淋巴細(xì)胞有絲分裂原, 能在體外激活T細(xì)胞增殖, 并促進(jìn)T細(xì)胞和單核細(xì)胞產(chǎn)生細(xì)胞因子[11]; 而脂多糖(Lipopolysaccharide, LPS)則刺激B細(xì)胞發(fā)生增殖。因此, 本研究采用混合淋巴細(xì)胞的增殖模型研究牡蠣蛋白活性肽的免疫活性。
3.2 對(duì)小鼠脾淋巴細(xì)胞因子IFN-γ和IL-2分泌的影響
采用雙夾心ELISA方法分別檢測(cè)小鼠脾淋巴細(xì)胞培養(yǎng)液上清中的細(xì)胞因子, 實(shí)驗(yàn)結(jié)果表明, 牡蠣活性肽對(duì)ConA激活的小鼠脾臟淋巴細(xì)胞上清中分泌的細(xì)胞因子IL-2, IFN-γ有顯著的抑制作用, 呈現(xiàn)一定的劑量依賴(lài)性, 見(jiàn)圖2、圖3。
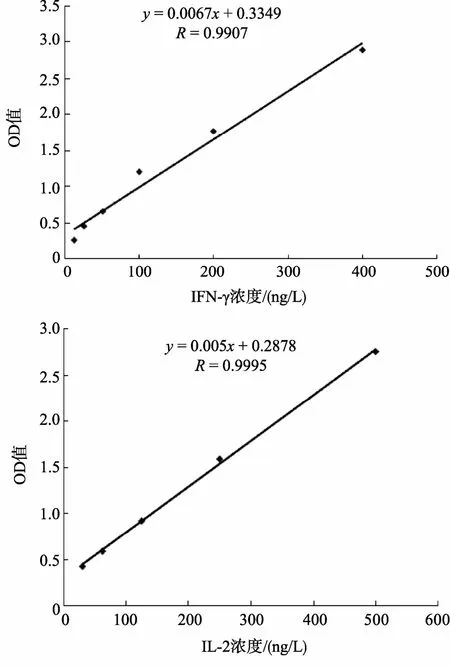
圖1 細(xì)胞因子IFN-γ和IL-2標(biāo)準(zhǔn)曲線 Fig. 1 The standard curves of cytokines IFN-γ and IL-2
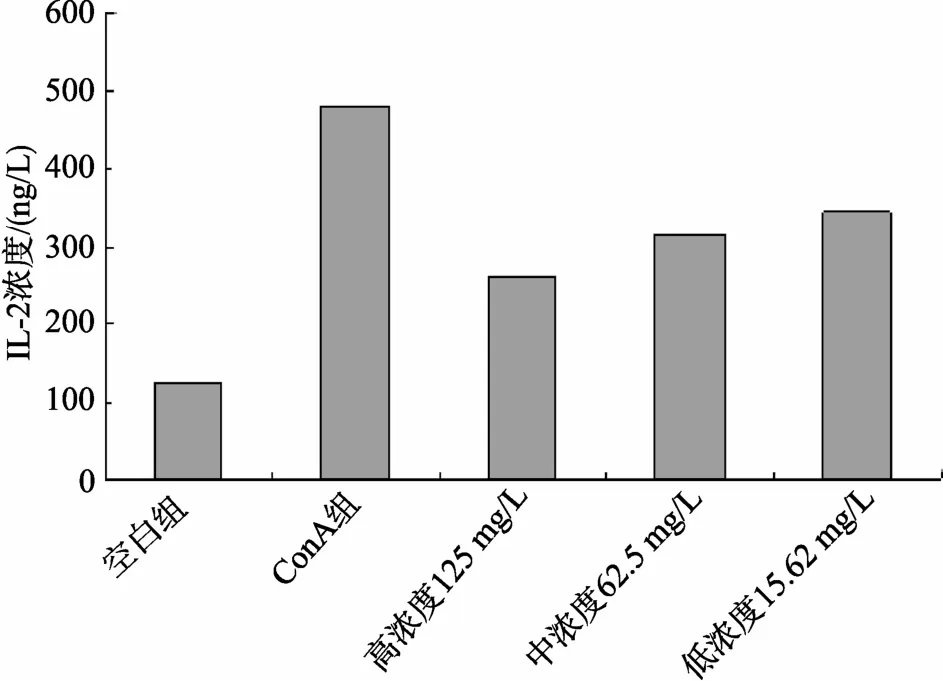
圖2 ELISA檢測(cè)牡蠣活性肽作用后對(duì)細(xì)胞上清中IL-2的影響 Fig. 2 The detection of IL-2 in supernatant influenced by oyster bioactive peptides using ELISA method
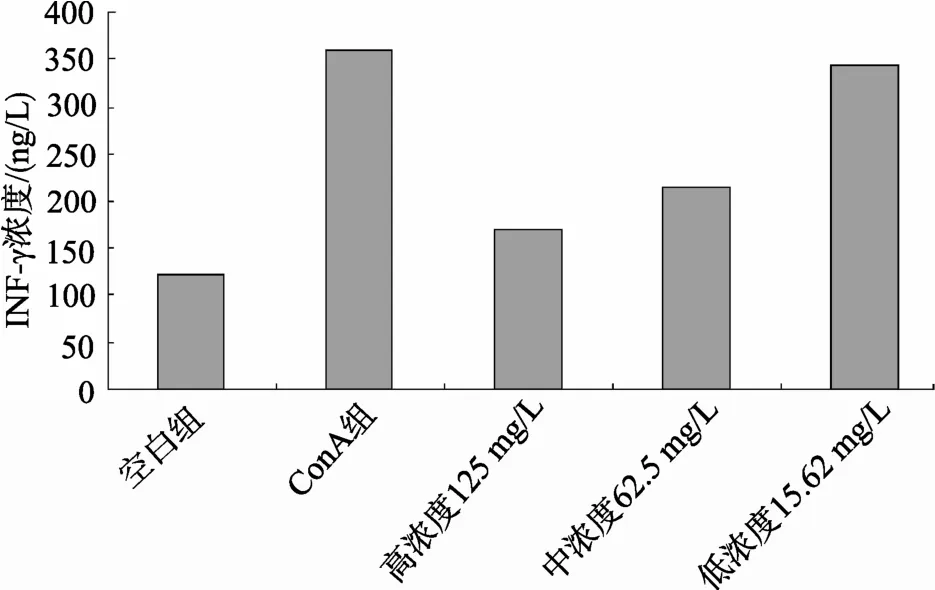
圖3 ELISA檢測(cè)牡蠣活性肽作用后對(duì)細(xì)胞上清中IFN-γ的影響 Fig. 3 The detection of IFN-γ in supernatant influenced by oyster bioactive peptides using ELISA method
細(xì)胞因子(cytokine)是一類(lèi)由抗原、有絲分裂原或其他刺激物活化的細(xì)胞分泌的、相對(duì)分子質(zhì)量為 15~30 kD的糖蛋白。包括淋巴細(xì)胞因子、單核細(xì)胞因子及其他細(xì)胞產(chǎn)生的細(xì)胞因子[12]。這些因子在體內(nèi)主要發(fā)揮抗感染、抗腫瘤作用、調(diào)節(jié)免疫、刺激造血細(xì)胞增殖分化以及參與和調(diào)節(jié)炎癥反應(yīng)等生物學(xué)功能。白細(xì)胞介素-2(IL-2), 主要由活化的CD4+T細(xì)胞和CD8+T細(xì)胞產(chǎn)生, 是所有T細(xì)胞亞群的生長(zhǎng)因子, 為調(diào)控免疫應(yīng)答的重要因子, 免疫抑制劑可以抑制其活性和生成。已有大量文獻(xiàn)報(bào)道, 在器官移植后的排斥反應(yīng)和自身免疫病如類(lèi)風(fēng)濕性關(guān)節(jié)炎、紅斑狼瘡、膜腎球腎炎、炎性腸病和免疫性溶血貧血等患者的血清或淋巴細(xì)胞培養(yǎng)上清液中, 炎癥性細(xì)胞因子IL-2的水平明顯升高[13]。IFN-γ可增強(qiáng)靶器官中抗原遞呈細(xì)胞或非抗原遞呈細(xì)胞異常表達(dá)組織相容性復(fù)合物分子, 促進(jìn)對(duì)自身抗原的識(shí)別和提呈功能, 激活自身反應(yīng)性T細(xì)胞, 從而誘發(fā)自身免疫性疾病。在自身免疫性肝炎、自身免疫性甲狀腺炎患者血清中IFN-γ明顯升高。本研究結(jié)果表明牡蠣蛋白活性肽可以抑制小鼠脾淋巴細(xì)胞分泌IL-2和TNF-γ, 由此推測(cè)牡蠣蛋白活性肽可能是通過(guò)抑制細(xì)胞因子的分泌來(lái)產(chǎn)生免疫抑制作用。
4 結(jié)論
近年來(lái), 器官移植的手術(shù)正以每年5000余例次遞增, 是器官衰竭治療的最后有效手段, 術(shù)后的特殊用藥——免疫抑制劑, 對(duì)器官移植患者存活時(shí)間、移植器官功能維持等方面起著舉足輕重的作用。而目前臨床上使用的免疫抑制劑多為進(jìn)口藥物, 價(jià)格昂貴, 此外臨床上常用的免疫抑制劑如糖皮素激素類(lèi)、環(huán)磷酰胺、甲氨喋呤、硫唑嘌呤、環(huán)孢菌素、抗淋巴細(xì)胞血清、抗白介素-1及2受體抗體等, 毒副作用較大。因此尋找開(kāi)發(fā)高效、低毒的免疫抑制劑, 是當(dāng)今器官移植藥物研究的熱點(diǎn)。
本研究發(fā)現(xiàn)在一定濃度范圍內(nèi)牡蠣蛋白活性肽能抑制小鼠脾淋巴細(xì)胞和混合淋巴細(xì)胞的增殖, 具有明顯的免疫抑制活性。研究結(jié)果顯示牡蠣活性肽有希望作為一種新的免疫抑制劑進(jìn)行開(kāi)發(fā)應(yīng)用研究, 在動(dòng)物皮瓣移植模型上進(jìn)一步探討其體內(nèi)抗排斥功效。
[1] 尹淑敏, 李鳳謙. 牡蠣的藥用[J]. 中國(guó)藥學(xué)雜志, 2001, 29(12): 751-752.
[2] 王穎, 馬安倫, 張惠珍, 等. 牡蠣提取物抗腫瘤作用的實(shí)驗(yàn)研究[J]. 中國(guó)海洋藥物, 1997, 16(1): 18-22.
[3] 曹棄元, 李永強(qiáng), 陳誠(chéng)欽, 等. 牡蠣提取物體外放射增敏作用[J]. 中國(guó)海洋藥物, 1993, 12(2): 11-13, 29.
[4] 陳艷輝, 李超柱, 吳磊, 等. 廣西產(chǎn)牡蠣多糖的制備和抗腫瘤活性初步研究[J]. 中國(guó)現(xiàn)代醫(yī)學(xué)雜志. 2010, 20(7): 1004-1005.
[5] 桑希生, 李慶云, 徐強(qiáng), 等. 牡蠣湯對(duì)免疫性肝損傷小鼠肝組織NO、IL-和TNF-α影響[J]. 中醫(yī)藥信息, 2007, 24 (5): 68-69.
[6] 黃大川, 李祺福, 李鵬, 等. 牡蠣低分子活性物質(zhì)對(duì)人肺腺癌A549細(xì)胞的生物學(xué)效應(yīng)[J]. 廈門(mén)大學(xué)學(xué)報(bào)(自然科學(xué)版), 2002, 41(5): 438-479.
[7] 吳繼衛(wèi), 何海倫, 路敬濤, 等. 海洋生物蛋白的酶解及酶解產(chǎn)物的抗氧化活性[J]. 海洋科學(xué). 2005. 29(3): 76-80.
[8] Yamamoto N, Ejiri M, Mizuno S. Biogenic peptides and their potential use [J]. Curr Pharm Des, 2003, 9(16): 1345-1355.
[9] Kong Z L, Chiang L C, Fang F. Immune bioactivity in shellfish toward serum free cultured human cell lines [J]. Bioscience Biotechnology and Biochemistry 2006, 61(1): 24-28.
[10] 陳艷輝, 李超柱, 吳磊, 等. 動(dòng)物蛋白酶酶解制備廣西產(chǎn)牡蠣肉抗腫瘤活性肽的實(shí)驗(yàn)研究[J]. 食品工業(yè)科技, 2010, 331(8): 167-169.
[11] Angela S.Wermerskirchen, Dorian H.LaTocha, Benjamin L.Clarke.Adrenocortropid hormone controls Concanavalin A activation of rat lymphocytes by modulating IL-2 production.Life Sciences 2000, 67: 2117-2187.
[12] 仲人前. 細(xì)胞因子與自身免疫性疾病的關(guān)系[J]. 中華檢驗(yàn)醫(yī)學(xué)雜志, 2000, 23: 4.
[13] Choy E H S, Panayi G. Cytokine pathways and joint inflammation in rheumatoid arthtitis.N Engl J Med 2001, 344: 907-915.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫(huà)刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂(lè)園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
