豬流行性腹瀉病毒P55野生毒株主要結構蛋白遺傳分析
2013-12-18 10:00:54高杏丹陳曦楊金先俞伏松李英英林天龍宋鐵英
福建畜牧獸醫 2013年2期
高杏丹陳 曦楊金先俞伏松李英英林天龍宋鐵英*
(1.福建師范大學生命科學學院 福州 350108;2.福建省農業科學院生物技術研究所 福州 350003)
豬流行性腹瀉病毒(porcine epidemic diarrhea virus,PEDV),是一種引發豬消化道黏膜發炎和糜爛等流行性腹瀉病癥的病毒。其臨床特征表現為發熱、厭食、嘔吐、腹瀉以及脫水等,且各種年齡和不同品種的豬群都有易感性,尤其對哺乳仔豬危害最嚴重,死亡率高。PED于1971年在英國首次被報道[1],此后日本、韓國及泰國等多個國家都報道了PEDV的發生[2-4]。PEDV基因組是單股正鏈具有感染性的RNA,全長27 000~33 000個核苷酸,編碼非結構蛋白(復制酶多聚蛋白1ab,ORF3蛋白)和4種主要結構蛋白。結構蛋白從5’→3’端依次為:纖突蛋白(Spike,S)、小膜蛋白(sma11Membrane,sM)、膜蛋白(Membrane,M)和核衣殼蛋白(Nuc1eocapsid,N)。
近年來,PED在我國呈現了不斷擴散和暴發的態勢[5-7]。特別是2010年以來,在全國包括福建地區均出現了大面積的暴發,給養豬業發展造成沉重的打擊[8]。2010-2012年,本研究室對福建省內20多個豬場進行了分子流行病學調查,篩選到一株標號為P55的野生毒株,其在與病毒毒力相關的ORF3區域的412到465位置出現了與CH/GSJIII/07(GeneBank:GU372743)相似的大片段缺失[9]。為研究P55片段中是否存在有其他影響病毒毒力的變異,需要對該株的其余結構基因展開研究。本文以野生變異毒株P55株為研究對象,通過對M蛋白和N蛋白的核苷酸和氨基酸序列與參考毒株的比較分析,同時參考先期研究中的S1和ORF3蛋白的結果[10],充分評估該毒株的變異情況,為進一步的功能性研究奠定基礎。
1 材料與方法
1.1 病料收集與處理 病料樣品采自福建省某豬場內表現急性腸炎和水樣腹瀉仔豬的部分腸組織。樣品采集后,置于研缽研磨,加入pH7.4的 PBS制成1:9的組織勻漿,在渦旋儀上振蕩混勻,1 700×g離心10 min,吸取上清液,-80℃凍存備用。
1.2 引物的設計與合成 參照GenBank中己發表的PEDV CV777 M基因和N基因序列,設計特異性擴增引物,該引物由寶生物大連(TaKaRa)有限公司合成。見表1。

表1 M與N基因對應的特異性引物
1.3 總RNA的提取 取200 uL樣品上清液,加入800 uL RNAiso P1us Reagen(t寶生物工程有限公司)振蕩混勻,室溫靜置5 min后,使用氯仿和異丙醇進行抽提、離心,所得沉淀用乙醇洗滌,自然干燥后加入RNase-free水溶解沉淀,即為所提取RNA模板,-40℃凍存備用。
1.4 PEDV M、N基因的擴增 使用PrimescriptOne Step RT-PCR Kit Ver.2(寶生物工程有限公司)進行基因擴增。反應總體系為25 uL:包含PrimescriptOne Step Enzyme Mix 1 uL、2×1 Step Buffer 12.5 uL、上游引物和下游引物(10 pmo1/uL)各 1 uL、RNA模板0.5 uL、雙蒸水9 uL。反應條件為:50℃反轉錄30 min,94℃預變性2 min,94℃變性30 s、50℃退火30 s、72℃延伸1 min,30個循環。PCR產物進行1.5%瓊脂糖凝膠電泳分析,確認結果后產物送交Takara公司測序。
1.5 序列比對及進化樹分析 將所得P55毒株的M與N基因序列與選擇的GeneBank中的參考序列,置入MegA1ign4.0(DNA Star)程序中,采用C1usta1W方法加以比較,得到各毒株間的遺傳距離,以此為基礎繪制其遺傳系統進化樹,結合前期研究結果中S1和ORF3區的進化樹,進行P55與各毒株間的遺傳距離計算和分析。
2 結果與分析
2.1 PEDV目的片段的RT-PCR鑒定 采用兩組特異性引物(見表1)擴增所得的RT-PCR結果見圖1,對應的目的片段M、N的長度分別為746 bp和1 464 bp。

圖1 PEDV目的片段的RT-PCR擴增
2.2 M基因的分析 試驗株P55獲得的M基因由746個核苷酸組成,(G+C)含量為45.08%,編碼226個氨基酸,蛋白分子量約為25.38 kD,GenBank登錄號為JQ723736。選取與P55同源性較高(Max score>1 210)的20株毒株為參考進行類比。比較結果顯示,P55毒株M基因序列無特有的片段缺失和插入,也未出現特有的堿基點突變。此外,在P55M基因序列的啟動子上游11個核苷酸的位置,存在有高度保守的轉錄調控因子(TRSs),其核心區由6個核苷酸組成(5’-ATAAAC-3’)[11]。
采用MegA1ign4.0(DNA Star)軟件得到的氨基酸進化分析結果顯示:M蛋白總變異度為2.4。氨基酸序列分析結果未發現P55存在特有突變,其具有的潛在的天冬酰胺酸糖基化結合位點的數量(3個),與經典株CV777一致。在氨基酸進化樹分組中P55與從中國南方的廣東、湖北、上海分離的5株病毒(CHGD-01、GD-A、AJ1102、ZJCZ4、LC) 遺傳距離較近。見圖2。

圖2 P55株M基因遺傳進化樹
2.3 N基因的分析 試驗株P55獲得的N基因由1 464個核苷酸組成,(G+C)含量為47.51%,編碼有441個氨基酸,蛋白分子量約為48.95 kD,GenBank登錄號為JQ723731。從GeneBank中選取選取與P55同源性較高(Max score>2 300)的 22株毒株為參考進行類比。結果顯示,P55毒株N基因序列無特有的片段缺失和插入,但出現了4個該株特有的點突變,影響到3個氨基酸的翻譯,位點見表2。這些變異并未改變其在啟動子上游9個核苷酸處存在的轉錄調控因子,該因子的核心區為 5’-AGAAAC-3’[11]。

表2 P55株N蛋白變異情況
同樣采用MegA1ign4.0(DNA Star)軟件得到的氨基酸進化分析結果顯示,N蛋白總變異度為3.7。此外,P55毒株N蛋白的氨基酸序列存在6個潛在的天冬酰胺酸糖基化結合位點,與CV777毒株的N蛋白一致。在氨基酸進化樹分組中P55與CHGDS-09、CH-IMT-06、CH-S、attenuated DR13 遺傳距離接近,其中attenuated DR13來源于韓國,其余毒株均從中國分離(見表3、圖3)。
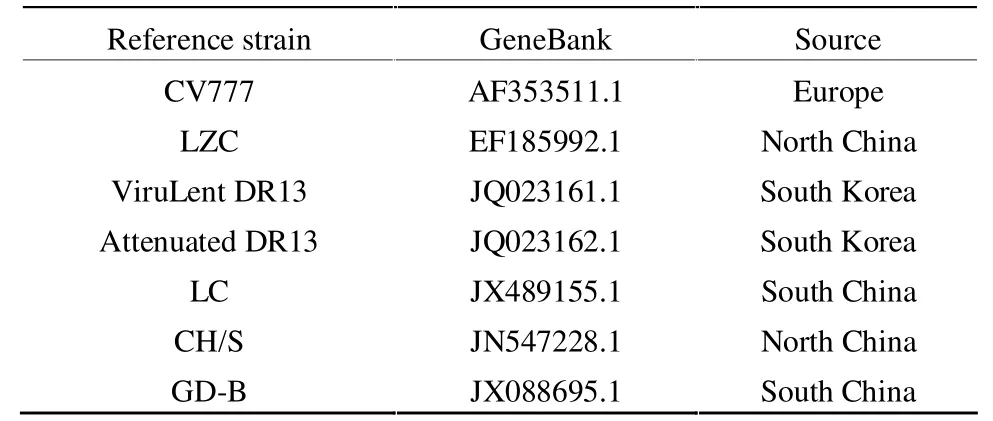
表3 具有全基因組序列的PEDV參考株

圖3 P55株N基因遺傳進化樹
2.4 M、N、S1、ORF3蛋白的遺傳距離分析 抽選在P55毒株的4組蛋白比較中共有的參考毒株進行分析,并對各組值進行方差分析(見表4)。結果顯示,遺傳距離:S1蛋白>ORF3蛋白>N蛋白>M蛋白。
3 討論
1)20世紀90年代以來,雖然各種滅活疫苗和弱毒疫苗已經被應用于PED的防控,但其在中國的流行性暴發仍時有發生,特別是2010年以來PED大規模暴發對我省養豬業造成的重大的經濟損失[8,12],警示我們有必要對PEDV毒株的變異情況進行調查。

表4 P55株與參考株M、N、S1、ORF3蛋白的遺傳距離
2)本研究中,P55的M蛋白未發現特殊變異,進化樹變異度和遺傳距離的方差結果顯示,M蛋白在主要結構蛋白中是最為保守的。由于M蛋白在病毒粒子的組裝和出芽過程中具有重要作用[13],且能介導機體產生干擾素[14],因此,研究結果再次證明可以將M基因作為PEDV基因工程疫苗的候選基因。另外,在進化樹中,P55所在分支的所有毒株均來自于中國,推測在M蛋白區域,P55表現出一定的地域保守性。
3)相較M蛋白,P55的N蛋白變異度增加,其N蛋白基因的ORF均由1326 nt組成,無核苷酸缺失和插入現象,但是堿基序列出現了4個點突變,從而影響了3個氨基酸的變異,由于N蛋白參與病毒的復制和轉錄[15],該變異是否會影響其功能尚待研究。
4)將前期結果中P55毒株的S1和ORF3蛋白與本研究的M、N蛋白遺傳距離的方差分析結果進行比較,由于方差表現的是各毒株與P55之間遺傳距離的變化程度,該結果揭示了各蛋白保守程度的差異,M和N蛋白的變異度較S1和ORF3小[10]。同時,P55與各毒株之間的平均遺傳距離結果表明,P55毒株和來源于韓國的attenuated DR13親緣關系較近。
5)Song等證明,ORF3區域的缺失或變異會導致病毒毒力的改變[16]。但近期的相關研究中陸續報道了4株與P55的ORF3區域存在類似突變的野生毒株,其中3株分離自中國,說明類似的缺失變異毒株在我國出現并已開始流行擴散[9,17](CH/BJ/2011 GenBank:JQ027019;Zhejiang-08 GenBank:JX0027 03)。因此,研究病毒毒力的分子機制對預防和控制該類變異毒株至關重要。對P55的M和N蛋白區域基因序列與參考序列進行比較,未發現特殊的缺失和變異。綜上所述,P55毒株中是否存在有除了M、N、S1之外的其他影響病毒毒力因素,尚待進一步對病毒的如S2蛋白區和PPo1區等進行分析研究。
[1] OLDHAMJ.Pig Farming(October Supp1)[M].1972:72-73.
[2] TAKAHASHI K,OKADA K,OHSHIMA K.An outbreak of swine diarrhea of a new-type associated with coronavirus-1ike partic1es in Japan [J].Nihon Juigaku Zasshi,1983,45(6):829-832.
[3] PURANAVEJA S,POOLPERM P,LERTWATCHARASARAKUL P,et a1.Chinese-1ike strain of porcine epidemic diarrhea virus,Thai1and [J].Emerg Infect Dis,2009,15(7):1112-1115.
[4] PARK SJ,MOON H J,YANG J S,et a1.Sequence ana1ysis of the partia1 spike g1ycoprotein gene of porcine epidemic diarrhea viruses iso1ated in Korea [J].Virus Genes,2007,35(2):321-332.
[5] 劉暢.目前豬傳染性腹瀉疾病的流行情況與防控方案[J].今日畜牧獸醫,2012(8):15-16.
[6] 李思銀,楊亮宇,楊玉艾.豬流行性腹瀉的實驗室診斷方法[J].豬業科學,2010(12):54-57.
[7] 林忠武.豬流行性腹瀉的流行特點及其防治措施[J].福建畜牧獸醫,2011,33(4):14-15.
[8] 張世忠,江斌.2011年福建省豬流行性腹瀉的流行特點及其防治措施[J].福建畜牧獸醫,2012,34(2):23-25.
[9] CHEN J,WANG C,SHI H,et a1.Mo1ecuLar epidemio1ogy of porcine epidemic diarrhea virus in China.[J].Arch Viro1,2010,155(9):1471-1476.
[10] Chen X,Yang J,Yu F,et a1.Mo1ecuLar characterization and phy1ogenetic ana1ysis of porcine epidemic diarrhea virus(PEDV)samp1es from fie1d cases in Fujian,China[J].Virus Genes,2012,45(3):499-507.
[11] PENZES Z,GONZALEZ J M,CALVO E,et a1.Comp1ete genome sequence of transmissib1e gastroenteritis coronavirus PUR46-MAD c1one and evo1ution of the purdue virus c1uster[J].Virus Genes,2001,23(1):105-118.
[12] 徐國棟,李峰,張廣峰.國內豬流行性腹瀉防治概況[J].畜牧與獸醫,2011,43(12):88-93.
[13] DE HAAN C A,KUO L,MASTERS P S,et a1.Coronavirus partic1e assemb1y:primary structure requirements of the membrane protein [J].J Viro1,1998,72(8):6838-6850.
[14] LAUDE H,GELFI J,LAVENANT L,et a1.Sing1e amino acid changes in the vira1g1ycoprotein M affect induction of a1pha interferon by the coronavirus transmissib1e gastroenteritis virus[J].JViro1,1992,66(2):743-749.
[15] Jian-Qiang Li,Ji-Xing Liu,Xi Lan,et a1.C1oning the Structure Genes and Expression the N Gene of Porcine Epidemic Diarrhea Virus DX[J].中國病毒學:英文版,2009,24(3):179-186.
[16] SONG D S,YANG J S,OH J S,et a1.Differentiation of a Vero ce11 adapted porcine epidemic diarrhea virus from Korean fie1d strains by restriction fragment 1ength po1ymorphism ana1ysis of ORF3 [J].Vaccine,2003,21(17/18):1833-1842.
[17] PARK SJ,KIM H K,SONG D S,et a1.Mo1ecuLar characterization and phy1ogenetic ana1ysis of porcine epidemic diarrhea virus(PEDV)fie1d iso1ates in Korea[J].Arch Viro1,2011,156(4):577-585.
