兩種滸苔無機碳利用對溫度響應的機制
2013-12-16 08:20:34徐軍田王學文鐘志海姚東瑞
生態學報 2013年24期
關鍵詞:利用
徐軍田,王學文,鐘志海,姚東瑞,2
(1.淮海工學院海洋學院,連云港 222005;2.江蘇省中國科學院植物研究所,南京 210014)
溫度是影響生物過程最普遍的氣候因素。迄今為止向大氣排放的CO2已經使得地球比工業革命前變暖2.4℃,全球暖化的趨勢在本世紀還將進一步的加速 (IPCC 2001),預計本世紀末溫度將上升2—5℃,這將帶來許多生物種的滅絕[1]。大型海藻構成海洋總初級生產力的10%左右,是近岸海域重要的初級生產者,在近岸碳循環中起著很重要的作用[2]。溫度的變化對大型海藻的生長、代謝、碳平衡等產生重要影響,并進而影響到海藻季節與地理分布狀況[3]。
在自然海水中,在通常的pH值(8.0—8.2)和20℃條件下的濃度約為2mmol,而 CO2僅為10 μmol,由于Rubisco利用CO2的半飽和常數(Ks)為30—60 μmol[4]。在長期的進化過程中,許多大型海藻形成了無機碳濃縮機制(CCMs)來更有效地利用無機碳進行光合作用。除直接利用擴散進入細胞的CO2外,CCMs主要包括以下途徑:①依賴胞外碳酸酐酶(CA)的催化脫水形成 CO2,CO2進而被細胞吸收利用[5];②在細胞膜外酸性區域脫水而形成CO2,CO2進而被細胞吸收利用[6];③ 耗能的的直接吸收利用(此過程可被抑制劑DIDS所抑制)[7];④的協同運輸或的反向運輸[6]。這些機制得到了廣泛的研究,實驗主要以改變培養介質里的無機碳濃度以及pH值來測定這種機制的變化,但對于溫度的效應,卻未見報道。
滸苔(Ulva prolifera)和緣管滸苔(Ulva linza)是沿海潮間帶常見的大型綠藻,也是近幾年來綠潮爆發的主要種類,其生理特性對環境的變化得到了很好的研究[8-12],在此基礎上本文以這兩種滸苔為材料,研究無機碳利用機制對溫度的響應,為探討近岸大型海藻無機碳利用機制與溫度變化的關系提供一定的實驗基礎,同時對了解綠潮在響應全球變暖過程中的變化提供一定的數據支持。
1 實驗材料和方法
1.1 實驗材料
緣管滸苔(Ulva linza)和滸苔(Ulva prolifera)分別采自于江蘇連云港高公島和連島海域。材料用過濾海水清洗干凈后,選取色澤鮮綠,長度適中的健康藻體于玻璃水族箱內暫養,暫養的條件是溫度為15℃,光強為200μmol m-2s-1,光周期為12h∶12h;培養液為過濾的自然海水,每2d更換1次,每天24h通氣培養。
1.2 pH值漂移曲線的測定
稱取0.3g的緣管滸苔和滸苔藻體放于含有15ml過濾滅菌海水(pH值為8.0)的小玻璃瓶中,玻璃瓶放置于恒溫光照培養箱內,將藻體充分舒展后封閉,水體用磁力攪拌器均勻攪動,光照控制為200μmol m-2s-1,溫度為15℃。實驗設3個重復,每個小瓶口接一個pH計探頭,每隔一定的時間記錄pH的變化,當pH穩定后結束測定,這個穩定點就是pH值補償點。
1.3 光合作用的測定
用氧電極法(Hansatech,英國)測定緣管滸苔和滸苔的光合放氧速率。測定之前,把藻體用鋒利的小剪刀剪成約0.2 cm2大小的小片(緣管滸苔)和約1cm長的小條(滸苔),在室內低光下適應1—2h(溫度設定為測定時的溫度),最大程度的消除藻體損傷帶來的測定誤差。然后稱取大約20 mg鮮重的藻片(大約20片)放入裝有2 mL反應介質的反應槽中。用循環水浴精確控制反應槽內的溫度。飽和光強設置為600μmol m-2s-1。
1.4 無機碳利用機制的研究
為精確研究溫度對緣管滸苔和滸苔無機碳利用的影響,采用藻類碳酸酐酶的抑制劑AZ(acetazolamide)和EZ(6-ethoxyzolamide)(Sigma)(用0.05 mol/L NaOH配制成5mmol/L的原液),最終反應介質中的濃度為100μmol。AZ不能滲透細胞而只抑制胞外CA酶的活性[13-14],而EZ能滲透細胞從而抑制胞內和胞外CA兩者的活性[15]。通過和不加抑制劑的藻樣進行對比,從光合作用的光響應曲線來分析溫度的效應。
1.5 統計與分析
試驗數據采用one-way ANOVA(turkey)分析(采用Origin 7.0分析軟件),設顯著水平為P<0.05,所有實驗處理均為3個重復。
2 結果
pH漂移曲線表明這兩種滸苔都有很強的無機碳利用能力,其pH補償點分別高達10.19±0.01(緣管滸苔)和10.08±0.01(滸苔)(圖1)。
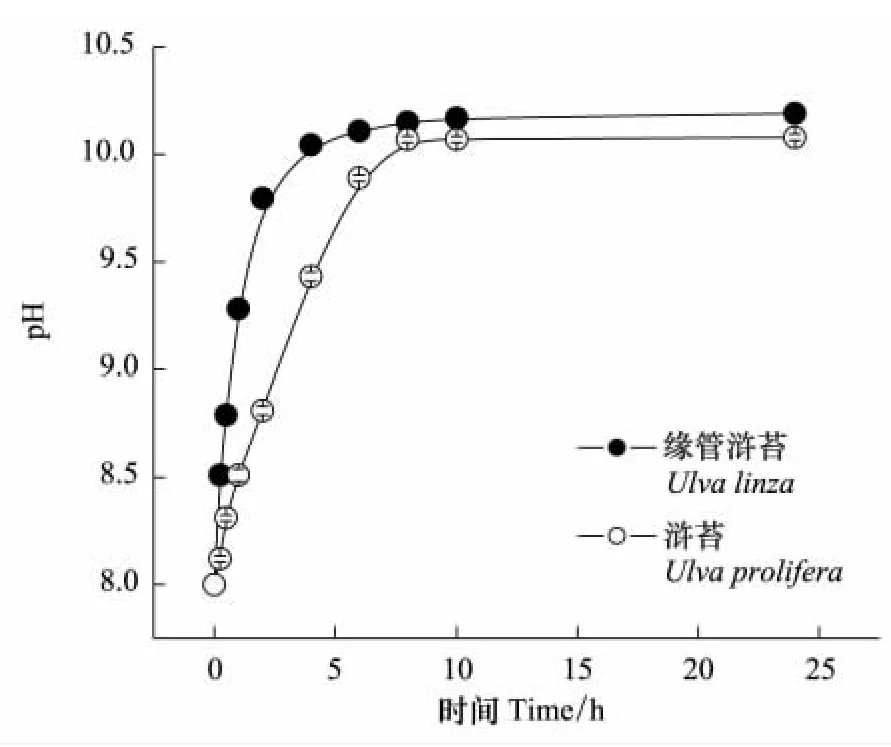
圖1 緣管滸苔和滸苔在15℃、光強為200μmol m-2s-1的pH漂移曲線Fig.1 The pH-drift curves of Ulva linza and Ulva prolifera under 20℃ and 200μmol m-2s-1condition
為了探討溫度對兩種滸苔無機碳利用能力的影響,測定了藻體在不同溫度下和加入不同抑制劑情況下的最大放氧速率,從圖中可以看出,緣管滸苔的最大光合作用速率(Pmax)隨著溫度的升高而顯著增加,到25℃達到最大,但在30℃顯著下降,下降比率為56%。當加入抑制劑AZ后,除在15℃外,其他溫度下的Pmax受到顯著的抑制。其Pmax隨著溫度的變化模式為,隨著溫度的升高,Pmax也顯著增加,但到15℃后,Pmax維持在一個恒定的水平,直到30℃才顯著下降。而加入抑制劑EZ后,藻體的Pmax在所有的溫度下與加入AZ相比受到了更顯著的抑制,其Pmax隨溫度的變化模式為,隨著溫度的升高,Pmax也顯著增加,在30℃具有最大的Pmax(圖2)。通過抑制劑對光合作用的抑制率來看,AZ處理的藻體在5℃和30℃最大,達到70%,而在15℃最小,抑制率為零。整個抑制率隨著溫度的變化呈“V”字形變動。而EZ處理的藻體,抑制率隨溫度的變化基本維持在85%的水平,只是在30℃時光合作用的抑制水平下降為80%(圖2)。
滸苔的最大光合作用速率(Pmax)隨著溫度的升高而顯著增加,到25℃達到最大,但在30℃顯著下降,其下降比率與緣管滸苔相比要小,僅為20%。當加入抑制劑AZ后,所有溫度下藻體的Pmax都受到顯著的抑制,其隨著溫度的變化趨勢和對照情況下一致,都是隨著溫度的升高而顯著增加,在25℃達到最大,但在30℃顯著下降。加入抑制劑EZ后,藻體的Pmax在所有的溫度下與加入AZ相比受到了更顯著的抑制,其Pmax隨溫度的變化模式為,5℃下滸苔具有最低的Pmax,10℃處理下滸苔的Pmax約是5℃的2.5倍,但隨著溫度的增加,Pmax卻一直保持在10℃時的水平上,其處理間沒有顯著差異(圖3)。通過抑制劑對光合作用的抑制率來看,AZ處理的藻體在5℃時最低,僅為16%,在10℃時其抑制率上升為55%,隨著溫度的上升,抑制率卻保持穩定的水平,除5℃外,其他所有溫度間的抑制水平沒有顯著差異。而EZ處理的藻體,抑制率隨溫度的變化在5℃和10℃較低,約為70%,隨著溫度的增加,其他溫度處理下的抑制率基本維持在85%的水平(圖3)。
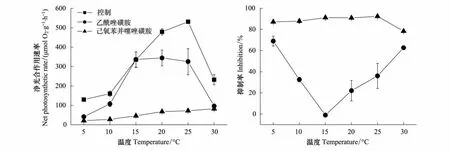
圖2 緣管滸苔在正常和抑制劑AZ和EZ存在情況下最大光合作用速率隨溫度的變化圖以及這兩種抑制劑對藻體最大光合作用速率的抑制率Fig.2 The change of the maximum photosynthetic rate of Ulva linza as function of temperatures with or without AZ and EZ inhibitors and the inhibition ratio of AZ and EZ for the maximum photosynthetic rate
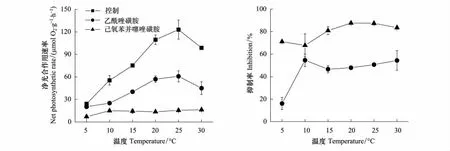
圖3 滸苔在正常和抑制劑AZ和EZ存在情況下最大光合作用速率隨溫度的變化圖以及這兩種抑制劑對藻體最大光合作用速率的抑制率Fig.3 The change of the maximum photosynthetic rate of Ulva prolifera as function of temperatures with or without AZ and EZ inhibitors and the inhibition ratio of AZ and EZ for the maximum photosynthetic rate
3 討論
本實驗結果表明,緣管滸苔和滸苔的pH漂移曲線說明這兩種海藻具有很強的無機碳利用能力,一般認為一種海藻的pH補償點達到9.2以上就表示其具有有效的利用能力[16-17],而這兩種海藻的pH補償點可以高達10以上,這表明這兩種海藻存在一種或者幾種高效的體外無機碳轉運形式。研究表明,緣管滸苔和滸苔具有明顯的胞外和胞內碳酸酐酶活性,這是大型海藻胞外CA酶催化的利用方式,是大型海藻無機碳利用的最普遍和最基本的方式[18-19]。而胞外CA酶活性本身僅僅是加速與CO2之間的相互轉化,而不能影響CO2的平衡濃度,并且形成的CO2主要是通過擴散作用而被動的進入細胞。這就意味著,這種方式的利用,在高pH(>9.4)時就不能很好地起作用,因為這時的CO2平衡濃度極低[20-21]。因此這表明,除胞外碳酸酐酶催化利用的方式外,緣管滸苔和滸苔必定還存在另外的利用方式。這在同屬的腸滸苔(Ulva intestinalis)中得到證實,其有兩種利用能力,即通過胞外CA酶催化的利用,以及通過陰離子交換蛋白抑制劑(4,4'-diisothiocyanatostilbene-2,2'-disulfonic acid,DIDS)敏感機制的直接吸收[21]。
通過比較緣管滸苔在不同溫度下AZ對其最大光合作用速率抑制率,得到在溫度最低的5℃和最高的30℃下,其抑制水平達到70%左右,這表明在這兩個溫度下無機碳的轉運主要是通過胞外碳酸酐酶的催化作用,而其他的轉運方式僅占很少的比例。而在15℃,加入胞外碳酸酐酶的抑制劑對藻體光合作用速率沒有顯著影響,這表明在這個溫度下其他的轉運方式可以彌補碳酸酐酶轉運方式所起到的作用。而其他的溫度下碳酸酐酶和其他轉運方式各占有一定的比例。這些實驗結果表明,除碳酸酐酶外的轉運方式對溫度具有很強的依賴性,在溫度很低或者很高的情況下,其效率與碳酸酐酶相比受到明顯的抑制,其最適的溫度為15℃,在這個溫度下其高的效率完全可以替代碳酸酐酶來為藻體進行有效的轉運。
而對于滸苔來說,無機碳利用機制對溫度的響應和緣管滸苔有顯著的差異,AZ在溫度最低的5℃具有最低的Pmax抑制率,其抑制水平僅為16%,這表明在低溫的情況下,除胞外碳酸酐酶的活性外,其他的轉運方式占有很高的比例。而從10℃開始,這種抑制水平就達到一個很高的值,并且一直保持在這個水平上(55%),這表明,當溫度升高到10℃以后,碳酸酐酶催化轉運所占的比率顯著增加,其他的轉運方式在無機碳利用過程中的作用與5℃相比顯著下降,并且在10℃以后直到30℃,這種比率保持恒定。
緣管滸苔光合作用能力最強的溫度是在25℃,而加入抑制劑EZ后,最高光合作用的溫度為30℃,但這表明光合作用體系(包括光合作用電子傳遞、固碳以及碳獲取的各個組分)里酶的最適溫度是不相同的,EZ抑制了胞內外碳酸酐酶的活性,導致胞內無機碳庫產生向Rubisco供應的CO2過程受到抑制[15],藻體利用的CO2是胞內碳酸鹽系統中自然解離的產物,這樣藻體的光合作用能力取決于RUBISCO酶的活性以及CO2的解離速度。而正常情況下,光合作用能力的大小必須要考慮無機碳轉運體系里各種酶的活性以及效率。我們實驗結果說明在30℃情況下,無機碳轉運體系受到了一定程度的抑制,導致光合作用能力顯著下降。
但對滸苔來說,其光合作用能力最強的溫度也為25℃,加入抑制劑EZ后,最高光合作用速率從15℃開始就保持恒定的水平,這表明,在不同溫度下影響光合作用能力的主要是無機碳轉運體系所起的作用,當這些轉運體系受到抑制后,藻體僅僅是利用海水中和體內自然化學解離形成的CO2,溫度對其光合作用過程中各種酶的活性沒有顯著的影響。
綜上所述,溫度對緣管滸苔和滸苔的無機碳利用轉運體系的影響是不同的,他們之間存在著明顯的種間差異,緣管滸苔對溫度的依賴性要強于滸苔,其對溫度的適應范圍要窄于滸苔,尤其是在高溫下(30℃),緣管滸苔的最大光合作用能力與最適溫度相比下降了56%,而滸苔僅為20%,這表明在高溫的情況下,滸苔比緣管滸苔具有更強的生存適應能力,這是滸苔能夠在綠潮藻占有絕對優勢的原因之一,并且在北方海域滸苔綠潮的調查研究中得到證實[9]。對于緣管滸苔和滸苔的無機碳利用機制來說,其他轉運的方式是在正常海水pH下就可以起到有效的作用,還是當胞外碳酸酐酶受到抑制后的誘導使其產生作用,這還需要進一步的研究。
[1] Romm J.Cleaning up on carbon.Nature Reports Climate Change,2008,doi:10.1038/climate.2008.59.
[2] Leigh E G,Paine R T,Quinn J F,Suchanek T H.Wave energy and intertidal productivity.Proceedings of the National Academy of Sciences of the United States of America,1987,84(5):1314-1318.
[3] Barry J P,Baxter C H,Sagarin R D,Gilman S E.Climate-related,long term faunal changes in a California rocky intertidal community.Science,1995,267(5198):672-675.
[4] Cook C M,Colman B.Some characteristics of photosynthetic inorganic carbon uptake of a marine macrophytic red alga.Plant Cell and Environment,1987,10(3):275-278.
[5] Smith R G,Bidwell R G S.Mechanism of photosynthetic carbon dioxide uptake by the red macroalga Chondrus crispus.Plant Physiology,1989,89(1):93-99.
[6] Lucas W J.Photosynthetic assimilation of exogenousby aquatic plants.Annual Review of Plant Physiology,1983,34(1):71-104.
[7] Raven J A,Lucas W J.The energetics of carbon acquisition//Lucas W J,Berry J A,eds.Inorganic Carbon Uptake by Aquatic Photosynthetic Organisms.Rockwell:American Society of Plant Physiologists,1985:305-324.
[8] Gao S,Chen X J,Yi Q Q,Wang G C,Pan G H,Lin A P,Peng G,Peer W A.A strategy for the proliferation of Ulva prolifera,main causative species of green tides,with formation of sporangia by fragmentation.PloS One,2010,5(1):e8571,doi:10.1371/journal.pone.0008571.
[9] Zhang X W,Xu D,Mao Y Z,Li Y X,Xue S Y,Zou J,Lian W,Liang C W,Zhuang Z M,Wang Q Y,Ye N H.Settlement of vegetative fragments of Ulva prolifera confirmed as an important seed source for succession of a large-scale green tide bloom.Limnology and Oceanography,2011,56(1):233-242.
[10] Xu J,Liu S,Zou D,Guo G L,Li X S,Liu S X.Responses of Enteromorpha linza to the copper exposure under different N growth conditions.Marine Science Bulletin,2010,29(6):643-648.
[11] Lin A P,Wang C,Qiao H J,Pan G H,Wang G C,Song L Y,Wang Z Y,Sun S,Zhou B C.Study on the photosynthetic performances of Enteromorpha prolifera collected from the surface and bottom of the sea of Qingdao sea area.Chinese Science Bulletin,2009,54(3):399-404.
[12] Xu J,Gao K.Future CO2-Induced ocean acidification mediates the physiological performance of a green tide alga.Plant Physiology,2012,160(4):1762-1769.
[13] Haglund K,Pedersén M.Growth of the red alga Gracilaria tenuistipitata at high pH:influence of some environmental factors and correlation to an increased carbonic-anhydrase activity.Botanica Marina,1992,35(6):579-587.
[14] Axelsson L,Mercado J,Figueroa F.Utilization ofat high pH by the brown macroalga Laminaria saccharina.European Journal of Phycology,2000,35(1):53-59.
[15] Sültemeyer D F,Schmidt C,Fork H.Carbonic anhydrases in higher plants and aquatic microorganisms.Physiologia Plantarum,1993,88(1):179-190.
[16] Maberly S C,Madsen T V.The contribution of air and water to the carbon balance of Fucus spiralis.Marine Ecology Progress Series,1990,62(1/2):175-183.
[17] Johnston A M,Maberly S,Raven J A.The acquisition of inorganic carbon by four red macroalgae.Oecologia,1992,92(3):317-326.
[18] Mercado J M,Figuroa F L,Niell F X,Axelsson L.A new method for estimating external,carbonic anhydrase activity in macroalgae.Journal of Phycology,1997,33(6):999-1006.
[19] Larsson C,Axelsson L.Bicarbonate uptake and utilization in marine macroalgae.European Journal of Phycology,1999,34(1):79-86.
[20] Axelsson L,Ryberg H,Beer S.Two modes of bicarbonate utilization in the marine green macroalga Ulva lactuca.Plant Cell and Environment,1995,18(4):439-445.
[21] Larsson C,Axelsson L,Ryberg H,Beer S.Photosynthetic carbon utilizatin by Enteromorpha intestinalis(Chorophyta)form a Swedish rockpool.European Journal of Phycology,1997,32(1):49-54.
參考文獻:
[10] 徐軍田,鄒定輝,朱明,郭贛林,李信書,劉樹霞.不同N水平下緣管滸苔對重金屬銅的響應.海洋通報,2010,29(6):643-648.
[11] 林阿朋,王超,喬洪金,潘光華,王廣策,宋厲蕓,王志遠,孫松,周百成.青島海域漂浮和沉降滸苔的光合作用研究.科學通報,2009,54(3):294-298.
猜你喜歡
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
中學生數理化(高中版.高考數學)(2021年6期)2021-07-28 06:19:08
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
小學生學習指導(低年級)(2019年11期)2019-11-25 07:31:44
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
數學小靈通·3-4年級(2017年6期)2017-06-22 11:28:50
工業設計(2016年5期)2016-05-04 04:00:33
河北遙感(2015年4期)2015-07-18 11:05:06
