紫竹PPO基因的生物信息學(xué)及表達(dá)分析
2013-11-24 01:53:30郭小勤林新春
浙江林業(yè)科技 2013年2期
關(guān)鍵詞:分析
關(guān) 鷹,王 弋,郭小勤,林新春,方 偉
(浙江農(nóng)林大學(xué) 亞熱帶森林培育國(guó)家重點(diǎn)實(shí)驗(yàn)室培育基地,浙江 臨安 311300)
多酚氧化酶(polyphenol oxidase, PPO)是一種能與Cu鏊合的膜結(jié)合蛋白,于1895年被發(fā)現(xiàn),廣泛分布于動(dòng)物、植物、真菌和細(xì)菌中[1]。多酚氧化酶有三種類型,分別為酪氨酸酶、兒茶酚氧化酶和漆酶。植物中,多酚氧化酶參與生物合成、光合作用、植物抗逆及褐化等過(guò)程[2],尤其是在酶促褐變過(guò)程中,多酚氧化酶起關(guān)鍵作用[3],它能作用于底物酚類物質(zhì)而引起材料的褐化[4]。近幾十年來(lái),研究人員除了對(duì)多種植物的多酚氧化酶進(jìn)行研究外,還克隆了編碼基因,并對(duì)這些基因的結(jié)構(gòu)、表達(dá)情況及功能進(jìn)行了分析。
本實(shí)驗(yàn)從紫竹(Phyllostachys nigra)栽培類型一年紫中克隆了該基因的片段,對(duì)其序列分析顯示,該序列與此前從臺(tái)灣紫竹中獲得的另一個(gè)同源基因片段有一定的差異,顯示出該基因在不同栽培類型中具多態(tài)性。組織特異性分析結(jié)果顯示,該基因在一年紫幼嫩部位的表達(dá)量高于其他成熟組織。
1 材料與方法
1.1 材料
在浙江農(nóng)林大學(xué)翠竹園采集一年生紫竹的幼嫩葉片,采用CTAB法提取基因組DNA,-20℃保存?zhèn)溆谩T谕恢晟喜杉o、筍尖、全筍、鞭根、葉芽,采用Trizol法提取其總RNA,-70℃保存?zhèn)溆谩?/p>
1.2 紫竹PnPPO1基因片段的克隆
利用Primer軟件參照GeneBank上所登錄的PPO基因保守序列設(shè)計(jì)兼并引物,PnPPO1上游引物:5'-TTCGCGCTGCCGTTCTGGAA-3',下游引物:5'-GATGTGCCACATGCGGTCGA-3'。PCR反應(yīng)體系(20 μL)為:10×PCR buffer 2 μL,10 mmol/L dNTP 0.4 μL,上游引物(10 μmol/L)2 μL,下游引物(10 μmol/L)2 μL,Taq DNA聚合酶 0.15 μL,模板DNA 1.0 μL(100 ng),ddH2O 12.45 μL。PnPPO1反應(yīng)程序?yàn)椋?4℃:3 min,94℃:30 s,50℃:30 s,72℃:1 min,38個(gè)循環(huán),72℃:5 min,4℃保溫。瓊脂糖凝膠電泳檢測(cè)擴(kuò)增結(jié)果。用鼎國(guó)XA014-1膠回收試劑盒對(duì)目的片段進(jìn)行割膠回收,連接到pMD20-T載體(大連寶生物公司),連接產(chǎn)物轉(zhuǎn)化感受細(xì)胞DH5α,挑取白斑,提取質(zhì)粒,并進(jìn)行酶切檢測(cè),確定結(jié)果正確后進(jìn)行測(cè)序。
1.3 紫竹PnPPO1基因的組織特異性表達(dá)分析
以所提取的莖,筍尖,全筍,鞭根,葉芽的總RNA為材料,參照試劑說(shuō)明書(shū)分別進(jìn)行反轉(zhuǎn)錄,采用半定量PCR法對(duì)紫竹上述部位PnPPO1基因的表達(dá)進(jìn)行分析。以β-actin基因?yàn)閮?nèi)標(biāo)。β-actin反應(yīng)程序?yàn)椋?4℃預(yù)變性3 min;94℃變性30 s,50℃退火30 s,72℃延伸1 min,27個(gè)循環(huán),72℃延伸5 min。PnPPO1反應(yīng)程序如上,循環(huán)數(shù)為28循環(huán)。
2 結(jié)果與分析
2.1 PnPPO1基因片段的克隆及序列分析
以紫竹一年紫嫩葉 cDNA為模板,以PnPO1引物進(jìn)行擴(kuò)增,將擴(kuò)增產(chǎn)物回收,轉(zhuǎn)化,測(cè)序,測(cè)序結(jié)果顯示該片段長(zhǎng)438 bp,核苷酸序列比對(duì)結(jié)果發(fā)現(xiàn),該序列為PPO同源基因,該基因與水稻(Oryza sativa)、小麥(Triticum aestiv)、蓮(Nelumbo nucifera)、荔枝(Litchi chinensis)、玉米(Zea mays)等植物的PPO基因序列有很高的同源性,其中與小麥的同源性達(dá)到 75%,故將其命名為PnPPO1。DNAMAN分析顯示,該片段編碼146個(gè)氨基酸(圖1),蛋白分子量約為36.2 kD,等電點(diǎn)為5.03。Blastp結(jié)果顯示,該蛋白屬于酪氨酸酶(圖2),具有PPO蛋白典型的特征,含有1個(gè)保守的CuB結(jié)合區(qū),進(jìn)一步分析發(fā)現(xiàn),在該區(qū)域有4個(gè)保守的H(His)和一個(gè)F(Phe)(圖1)。
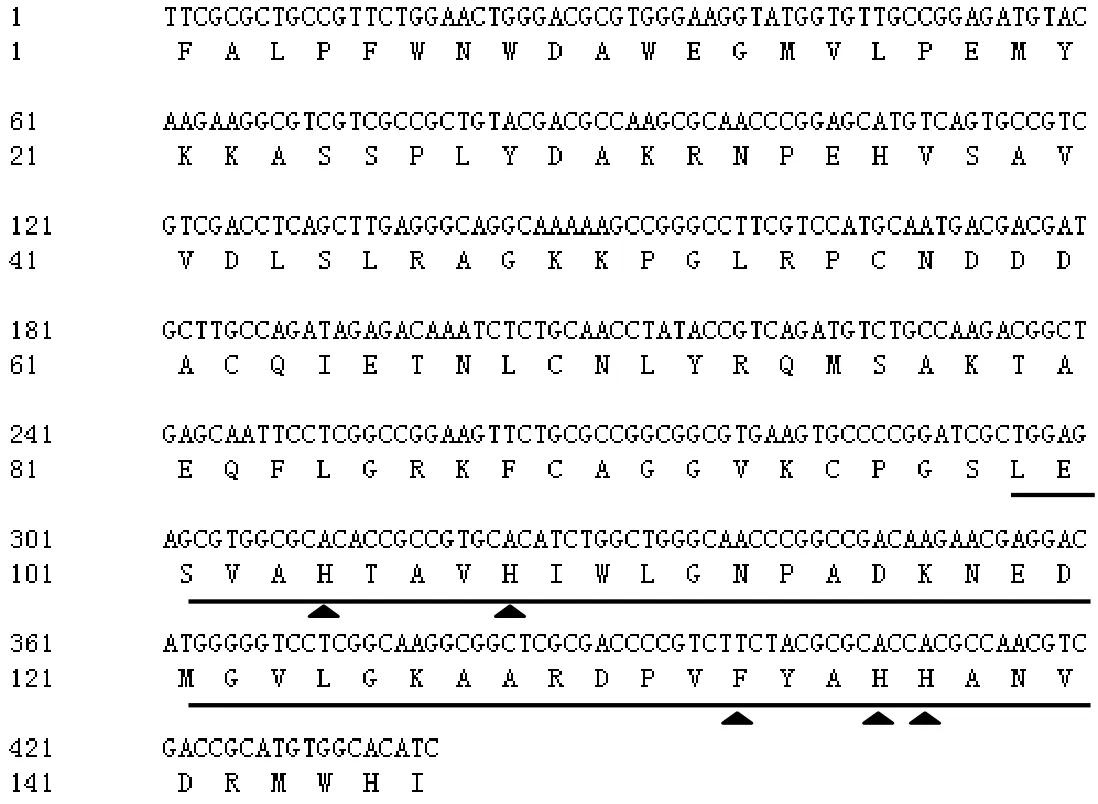
圖1 PnPPO1 cDNA及氨基酸序列Figure1 cDNA and amino acid sequence of PnPPO1

圖2 PnPPO1基因編碼的部分氨基酸序列的比對(duì)結(jié)果Figure2 The blastp of PnPPO1partial amino acid sequence
利用ProtParam對(duì)PnPPO1氨基酸序列的理化性質(zhì)進(jìn)行在線預(yù)測(cè),該氨基酸序列不穩(wěn)定指數(shù)較高(表1)。利用TMHMM2.0在線預(yù)測(cè)了PnPPO1的跨膜情況,結(jié)果顯示,PnPPO1蛋白在該區(qū)段沒(méi)有發(fā)現(xiàn)跨膜位點(diǎn)并且膜外概率高于膜內(nèi),PPO蛋白有可能是一個(gè)膜外蛋白(圖3)。
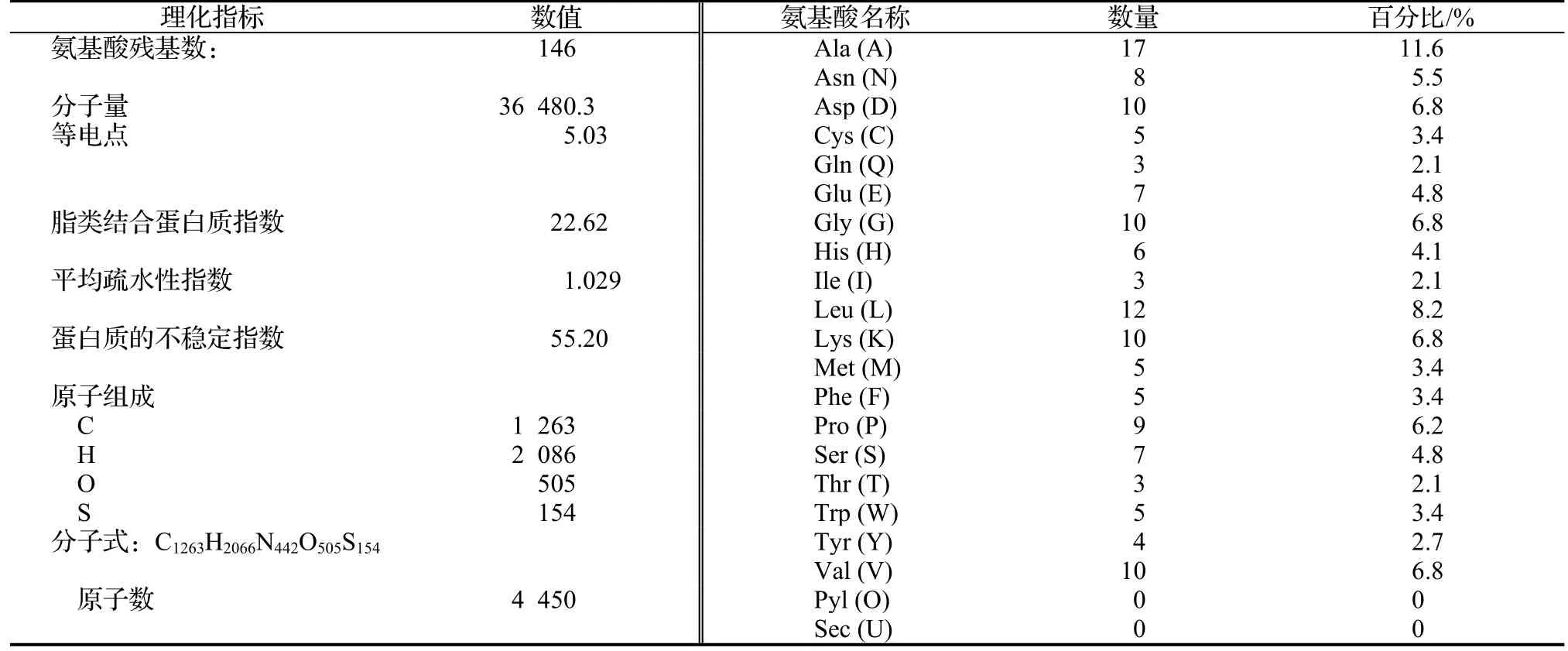
表1 一年紫PnPPO1氨基酸殘基理化性質(zhì)分析Table1 Physio-chemical characteristics analysis of PnPPO1 amino acids
運(yùn)用ProtScale對(duì)PnPPO1的氨基酸殘基進(jìn)行疏水性分析顯示,總體上親水氨基酸明顯多于疏水氨基酸(圖4),說(shuō)明一年紫PnPPO1蛋白在該區(qū)域顯示較高的親水性。
另外,所擴(kuò)增到的該氨基酸片段具有α-螺旋(含量10.28%),β-折疊(4.79%)且α-螺旋串通不同長(zhǎng)度的無(wú)規(guī)則卷曲(84.93%)。
對(duì)從一年紫和臺(tái)灣紫中獲得兩條PPO序列進(jìn)行分析,比對(duì)結(jié)果顯示,兩者同源性為86.45%,兩序列間除了片段缺失外,還存在明顯的 SNP位點(diǎn),多態(tài)性頻率為 1 SNP/27.3bp,其中以A/G轉(zhuǎn)換類型最多,占81.25%。由于單堿基的變異,一年紫和臺(tái)灣紫中酶切位點(diǎn)也會(huì)發(fā)生變化,從圖4中可以看出,由于單核苷酸的變異,有6處酶切位點(diǎn)發(fā)生了變化,其中5處為新酶切位點(diǎn)的產(chǎn)生,1處表現(xiàn)為酶切位點(diǎn)的變換(圖5)。
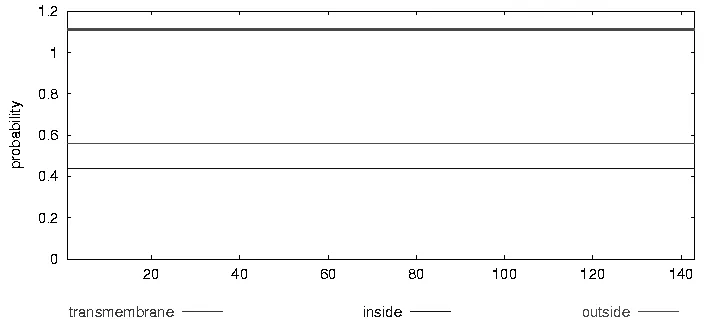
圖3 一年紫PnPPO1跨膜預(yù)測(cè)Figure3 Predicted transmembrane of PnPP01
2.2 PnPPO1基因的組織特異性表達(dá)
為研究PnPPO1基因在一年紫不同組織部位中的表達(dá)情況,以β-actin基因?yàn)閷?duì)照,對(duì)PnPPO1進(jìn)行RT-PCR分析,結(jié)果如圖6。由圖6可知,在所檢測(cè)的5個(gè)組織中,該基因在葉芽中的表達(dá)量最高,在鞭根中微量表達(dá),而在其余的組織未檢測(cè)到其表達(dá)。
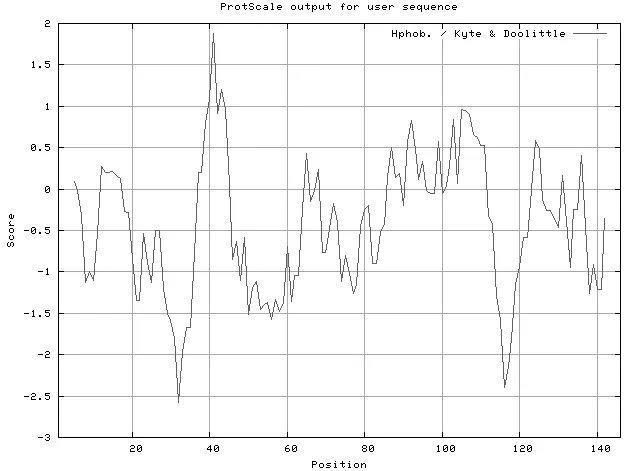
圖4 一年紫PnPPO1基因保守區(qū)推斷的氨基酸序列疏水圖Figure4 Hydrophobic analysis of on amino acids in the PnPPO1 sequence

圖5 一年紫PnPPO1與臺(tái)灣紫PnPPO的序列比對(duì)Figure5 Alignment sequence between PnPPO1and PnPPO1

圖6 PnPPO1的組織特異性表達(dá)Figure6 Expression pattern of PnPPO1
3 結(jié)論與討論
本研究利用同源克隆技術(shù)從一年紫嫩葉中獲得PnPPO1的cDNA序列,與之前從臺(tái)灣紫中獲得的序列的同源性為 86.45%,兩序列間除了堿基缺失外,還存在有豐富的SNP位點(diǎn),且以A/G型轉(zhuǎn)換位點(diǎn)最豐富。研究表明,PPO蛋白具有3個(gè)Cu結(jié)合區(qū)域(CuA、CuB、CuC),CuA與PPO的水溶性有關(guān),CuB是底物結(jié)合的最關(guān)鍵區(qū)域,CuC與分子氧相連且調(diào)節(jié)葉綠體中有害光氧化反應(yīng),參與電子傳遞,起能量轉(zhuǎn)換作用[5]。在實(shí)驗(yàn)獲得的146個(gè)氨基酸殘基中存在有CuB區(qū)域,該區(qū)域包含有保守的5個(gè)H(His),一個(gè)F(Phe),二級(jí)結(jié)構(gòu)分析發(fā)現(xiàn),本實(shí)驗(yàn)中所獲得的一年紫PnPPO1蛋白片段中含2個(gè)α-螺旋(含量10.27%),1個(gè)β-折疊(4.79%)串通不同長(zhǎng)度的無(wú)規(guī)則卷曲(84.93%)。
PPO是一種質(zhì)體酶,它在植物中的分布有兩個(gè)特點(diǎn):一是常存在于光合組織和非光合質(zhì)體中,二是在植物的幼嫩部分含量比較高[6]。研究表明,在不同物種、組織和發(fā)育階段,PPO的表達(dá)水平存在較大差異[7],如番茄中,該基因僅在葉片發(fā)育后期的柵欄細(xì)胞中表達(dá),而蛋白主要存在于嫩葉、花、根形態(tài)建成等時(shí)期的表皮細(xì)胞中,隨著葉片的發(fā)育成熟,幾乎檢測(cè)不到該蛋白的存在[8]。在馬鈴薯中,該基因僅在頂端的四個(gè)葉節(jié)點(diǎn)上能檢測(cè)到表達(dá),而其蛋白存在于所有的葉節(jié)點(diǎn)上。在蓮藕中,該基因在整個(gè)供試組織中都有表達(dá),但在莖尖和幼葉中表達(dá)量最高,其次為新鮮的蓮藕切片,而在花瓣和莖稈中表達(dá)量最低[9]。本研究結(jié)果表明,一年紫中PnPPO1在葉芽中表達(dá)量最高,在鞭根中微量表達(dá),而在其余組織中未檢測(cè)表達(dá),推測(cè)PnPPO1先在葉芽中表達(dá),然后再分配到各組織中,在特定的時(shí)空下被激活并擔(dān)負(fù)相應(yīng)的功能。
致謝:感謝浙江農(nóng)林大學(xué)特聘教授鄭康樂(lè)在實(shí)驗(yàn)和論文寫(xiě)作中提出的寶貴建議和意見(jiàn)。
[1]王乃棟,張麗霞,向勤锃,等.茶樹(shù)多酚氧化酶基因的生物信息學(xué)分析及原核表達(dá)[J].茶葉科學(xué),2011,31(1):33-39.
[2]郭小勤,李犇,阮曉賽,等.用ACGM分子標(biāo)記研究10個(gè)毛竹不同栽培變種的遺傳多樣性[J].林業(yè)科學(xué),2009,45(4):28-32.
[3]楊杰,曹卿,王軍,等.水稻多酚氧化酶基因功能標(biāo)記的開(kāi)發(fā)與應(yīng)用[J].中國(guó)水稻科學(xué),2011,25(1):37-42.
[4]王曼玲,胡中立,周明全,等.植物多酚氧化酶的研究進(jìn)展[J].植物學(xué)通報(bào),2005,22(2):215-222.
[5]Mayer A M.Polyphenol oxidases in plants and fungi: Going places? A review[J].Phytochemistry, 2006,67(21):18-31.
[6]SUN Youwei, HE Zhonghu, MA Wujun,et al.Alternative splicing in the coding region of Ppo-A1 directly influences the polyphenol oxidase activity in common wheat (Triticun aestivum L.) [J].Funct Integr Genomics, 2011, 11(1):85-93.
[7]王弋,郭小勤,婁永峰,等.紫竹多酚氧化酶基因克隆及其表達(dá)[J].浙江林學(xué)院學(xué)報(bào),2010,27(5):781-785.
[8]WANG Hong, JIANG Yuping, YU Haijing,et al.Light quality affects incidence of powdery mildew expression of defence-related genes and associated metabolism in cucumber plants[J].Eu J Plant Pathol,2010,127(1):125-135.
[9]張躍進(jìn),郝曉燕,梁宗鎖,等.郭宏波 藕多酚氧化酶基因(PPO)的克隆與表達(dá)分析[J].農(nóng)業(yè)生物技術(shù)學(xué)報(bào),2011,19(4):634-641.
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
