索拉非尼對敏感人肝癌細胞株MAPK信號通路基因表達的影響*
2013-11-07 06:02:46殷曉煜肖偉鍇梁力建
中國病理生理雜志 2013年1期
陳 東, 趙 鵬, 殷曉煜, 陳 偉, 肖偉鍇, 梁力建△
索拉非尼(sorafenib;商品名:多吉美)是一個多靶點的分子靶向藥物,對受體輔助因子1(receptor accessory factor,Raf-1)激酶、血管內皮生長因子受體(vascular endothelial growth factor,VEGFR)2、VEGFR3、Fms樣酪氨酸激酶(Fms-like tyrosine kinase,FLT)3、Ret、C-kit、血小板源性生長因子受體(platelet-derived growth factor receptor,PDGFR)等靶點有抑制作用。目前已有多項國際多中心研究證明索拉非尼對晚期原發性肝癌(hepatocellular carcinoma,HCC)有明顯、確切的療效,能明顯延長晚期肝癌的生存時間[1-4]。目前,索拉非尼已作為晚期、不可切除肝癌的標準治療而廣泛應用。Liu等[5]利用細胞實驗發現索拉非尼可抑制Raf,下調絲裂原激活蛋白激酶激酶(mitogen-activated protein kinase kinase,MAPKK,又稱MEK)和細胞外信號調節激酶(extracellular signal-regulated kinase,ERK)表達,即通過抑制Raf/MEK/ERK(MAPK)信號通路的表達來抑制HCC細胞的增殖。另外,絲裂原激活蛋白激酶(mitogen-activated protein kinase,MAPK)信號通路也是VEGFR2、VEGFR3、PDGFR等靶點的下游信號通路,但索拉非尼抑制MAPK信號通路的具體分子機制目前仍不明確。本研究進行完善的實驗設計,利用流式細胞術檢測細胞凋亡和CCK-8(cell counting kit-8)實驗,篩選出對索拉非尼敏感的肝癌細胞株,利用MAPK信號通路PCR基因芯片,檢測索拉非尼作用后MAPK信號通路中具體分子的基因表達變化,確定了索拉非尼作用HCC過程中MAPK信號通路中發生變化的具體作用分子。
材料和方法
1 材料
1.1 細胞株 Huh7、HepG2、SMCC7721和 PLC 細胞均購自中國科學院上海細胞庫,HepG2.2.15細胞購自上海復祥生物科技有限公司,MHCC-97H細胞購自上海中山醫院肝癌研究所。
1.2 基因芯片 人 MAPK PCR Array(PAHS-061,Qiangen)。
1.3 儀器與試劑 37℃、5%CO2培養箱(QUEUE),流式細胞儀(Becton-Dickinson),紫外吸收分光光度計(NanaDrop? ND-1000),DMEM培養基(Gibco),胎牛血清(Gibco),二甲基亞砜(DMSO,上海生物試劑廠),EDTA(華美生物工程公司);青/鏈霉素雙抗(penicillin-streptomycin,P/S,上海新先鋒藥業有限公司),CCK-8試劑盒(Sigma),sorafenib(德國拜耳醫藥公司),Annexin-PE/7-AAD凋亡檢測試劑盒(Becton&Dickinson),Trizol試劑(Invitrogen),RNeasy? MinEluteTM純化試劑盒(Qiagen),2× SuperArray PCR master Mix(Cat.No.PA-112)。
2 方法
2.1 敏感肝癌細胞株的篩選
2.1.1 細胞培養 將各HCC細胞株接種在含10%小牛血清,1%青霉素、鏈霉素的DMEM培養基中,置于37℃、5%CO2培養箱中培養,細胞生長良好呈單層貼壁生長,48~72 h換液1次。傳代時用0.1%胰蛋白酶消化。
2.1.2 索拉非尼藥物配制 用手術刀片將索拉非尼藥片糖衣輕輕刮去,刮凈糖衣,在稱量紙上將藥片研成粉末狀,稱重后將粉末倒入試管中,加入適量DMSO液,配制成濃度為100 μmol/L的索拉非尼工作液,-20℃保存。
2.1.3 流式細胞術檢測 每種細胞株設對照組、實驗組;細胞株用不含EDTA的胰酶消化;將濃度為2×108/L呈指數生長的細胞懸液接種于6孔板,每孔2.5 mL(即 5 ×105cells/well),孵育 24 h;對照組每孔加入0.5 mL DMSO,實驗組每孔加入索拉非尼0.5 mL 溶液(終濃度為 4 μmol/L),每組設 3 個復孔,孵育72 h;收集、離心,PBS液洗滌,重懸細胞,加入 5 μL Annexin V-FITC 和 5 μL propidium iodide(PI),混勻,室溫避光反應10 min,在1 h內上流式細胞儀檢測。
2.1.4 CCK-8法檢測 每種細胞設空白組(培養液+CCK-8)、對照組(細胞+培養液+CCK-8)、實驗組(細胞+培養液+索拉非尼+CCK-8);取對數生長期的Huh7、MHCC97、HepG2 、SMCC7721 和HepG2.2.15細胞,胰酶消化制成單細胞懸液,調整濃度為5×107/L,96孔板中每孔加入100 μL細胞懸液;各組設4個不同索拉非尼作用濃度,每個濃度設3個復孔;5%CO2、37℃孵育,吸凈培養液,加入無血清的DMEM液“饑餓細胞”24 h;吸凈培養液,對照組加入培養液200 μL,實驗組內加入不同量的索拉非尼溶液,余量液體用培養液補足,每孔液體量200 μL,孵育24 h;加入10 μL CCK-8溶液,孵育2 h,酶標儀在450 nm波長比色測定每孔吸光度(A)值,記錄結果;細胞存活率(%)=(Aa-Ab)/(Ac-Ab)×100%;a:實驗孔;b:空白孔;c:對照孔。
2.2 索拉非尼對敏感肝癌細胞株MAPK信號通路基因表達譜的影響
2.2.1 選擇上述實驗所篩選的敏感肝癌細胞株PLC細胞。
2.2.2 細胞分組 在3.5 cm直徑的培養盤中培養,分2組:PLC對照組(Pc組)和PLC+索拉非尼組(Pt組)。細胞貼壁后,Pc組加入DMSO液,Pt組加入索拉非尼,用藥濃度為5.25 μmol/L,加藥后繼續培養48 h。在培養盤中直接中加入1 mL Trizol試劑裂解細胞,裂解時用槍吸打幾次,然后分別放-80℃保存。置于干冰中運送至上海康成生物公司進行后續實驗。
2.2.3 PCR array操作 (1)RNA抽提:劇烈振蕩經Trizol裂解后樣本15 s,15~30℃孵育2~3 min。4℃下12 000×g離心15 min,將水相轉移到新離心管中。水相與異丙醇混合以沉淀其中的RNA,勻漿時加入1 mL Trizol試劑的同時加0.5 mL的異丙醇。混勻后15~30℃孵育10 min,4℃ 12 000×g離心10 min。移去上清液,加入75%乙醇,清洗RNA沉淀。振蕩后,4℃ 7 500×g離心5 min。去除乙醇溶液,干燥RNA沉淀5~10 min,溶解RNA,55~60℃孵育10 min,保存于-70℃。(2)cDNA合成:按試劑盒操作說明,每個純化柱的中心加入50 μL無RNA酶的H2O,放置2 min后8 000×g離心1 min,得到的溶液即純化的cRNA,暫時保存于冰中。(3)實時定量PCR:小心打開PCR array上的膜,加10 μL混合液到PCR array對應的每個孔中,小心蓋上蓋子密封PCR array,置于PCR儀進行PCR反應。
2.2.4 數據分析 采用ΔΔCt方法:(1)計算每個處理組中的每個通路相關基因的ΔCt:ΔCt(group 1)=average Ct- average of HK genes’Ct for group array;(2)計算2個PCR array(或2組)中每個基因的ΔΔCt:ΔΔCt= ΔCt(實驗組)-ΔCt(對照組);(3)通過2-ΔΔCt計算實驗組與對照組對應基因的表達差異。
3 統計學處理
數據采用SPSS 13.0軟件分析。數據以均數±標準差(mean±SD)表示,均數比較用t檢驗,以P<0.05為差異有統計學意義。
結 果
1 流式細胞儀檢測肝癌細胞株凋亡
Huh7、MHCC97H、HepG2 、SMCC7721、HepG2.2.15和PLC肝癌細胞株經索拉非尼作用后,細胞凋亡均較對照組明顯增加(P<0.05),見圖1。從凋亡細胞百分比可以看出,對索拉非尼較敏感細胞株為PLC細胞和 HepG2.2.15細胞(P < 0.01),對其相對不敏感細胞株為SMCC7721細胞和MHCC97H細胞,見圖2。
2 索拉非尼對各肝癌細胞株存活率的影響
由各細胞系的IC50值可看出,對索拉非尼相對敏感細胞株為PLC細胞,不敏感細胞株為SMCC7721細胞,見圖3。
3 MAPK PCR array實驗結果
與對照組比較,實驗組MAPK信號通路中有8個差異表達基因,其中>2.0倍有2個,≤0.5倍有6個。其中,細胞周期蛋白E1(cyclin E1,CCNE1)、周期素依賴性激酶 6(cyclin-dependent kinase 6,CDK6)和周期素依賴性激酶抑制因子1C(cyclin-dependent kinase inhibitor 1C,CDKN1C)與細胞周期的調節相關;jun原癌基因(jun proto-oncogene)、CREB結合蛋白(CREB-binding protein,CREBBP)、MAP 激酶相互作用絲/蘇氨酸激酶1(MAP kinase-interacting serine/threonine kinase-1,MKNK1)和MAP激酶激活的蛋白激酶3(MAP kinase-activated protein kinase 3,MAPKAPK3)與轉錄因子的活化有關,見表1。
討 論
本研究以索拉非尼作用不同肝癌細胞株Huh7、MHCC97H、HepG2、SMCC7721、HepG2.2.15 和 PLC,然后通過流式細胞儀檢測發現,索拉非尼實驗組對上述各種肝癌細胞的促凋亡作用均明顯強于對照組(P<0.05)。用不同濃度的索拉非尼作用于這些細胞,結果發現細胞不同其IC50值有差異,通過流式細胞術和CCK-8實驗,篩選出對索拉非尼敏感細胞株和不敏感細胞株分別為PLC和SMCC7721。篩選索拉非尼敏感和不敏感肝癌細胞株對于后續索拉非尼相關基礎研究有重要意義,可供同行參考。
研究表明,基因和蛋白質較少單獨起作用,它們往往通過復雜的網絡交互作用共同影響生物系統的功能。索拉非尼作用于肝癌涉及的信號通路尚包括STAT3[6]、PI3k/Akt/mTOR 信號通路等[7]。由于索拉非尼主要通過抑制Raf-1抑制MAPK信號通路,MAPK信號通路同時也是索拉非尼設計的其它作用靶點如VEGFR2、VEGFR3、PDGFR等分子下游的主要信號通路,因此,本研究重點深入研究MAPK信號通路具體分子基因的表達變化。
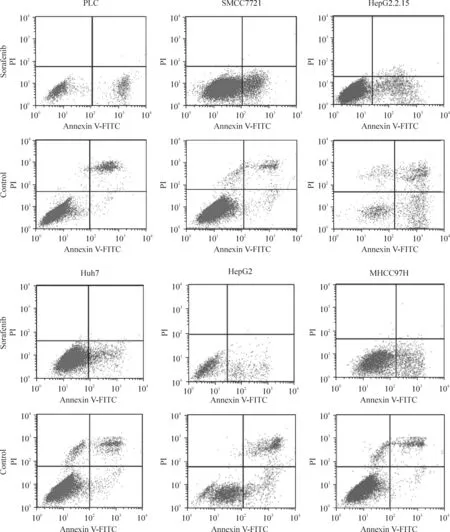
Figure 1.Cell apoptosis determined by flow cytometry.PLC,SMMC7221,HepG2.2.15,Huh7,HepG2 and MHCC97H cells were exposed to sorafenib for 72 h,then 5 μL Annexin V-FITC and 5 μL propidium iodide(PI)were added for 10 min of incubation.圖1 流式細胞術檢測細胞凋亡
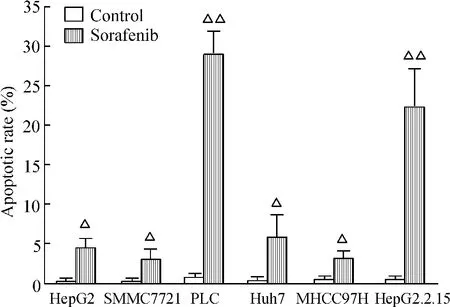
Figure 2.Apoptotic rates determined by flow cytometry.Mean ±SD.n=3.△P < 0.05,△△P < 0.01 vs control group.圖2 流式細胞術檢測凋亡細胞百分比
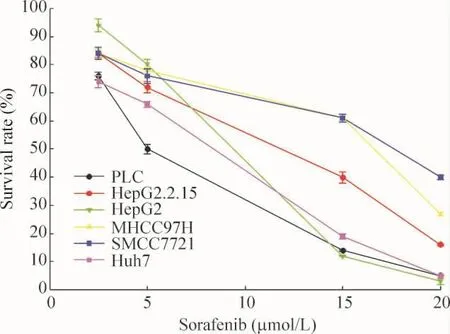
Figure 3.Survival rates of different hepatocarcinoma cell lines treated with sorafenib(2.5,5,15 and 20 μmol/L)for 24 h were determined by CCK-8 assay.Mean ±SD.n=3.The IC50 values of sorafenib were 5.25 μmol/L for PLC cells,5.30 μmol/L for HepG2.2.15 cells;6.80 μmol/L for Huh7 cells,7.01 μmol/L for HepG2 cells,11.7 μmol/L for MHCC97H cells and 15.0 μmol/L for SMCC7721 cells.圖3 索拉非尼作用后各肝癌細胞株的存活率
MAPK信號通路PCR芯片包含了與MAPK信息轉導通路有關的全部84種基因。實驗組MAPK信號通路中有8個差異表達基因,其中>2.0倍的有2個,≤0.5倍的有6個。其中,CCNE1、CDK6和 CDKN1C與細胞周期的調節相關;jun、CREBBP、MKNK1和MAPKAPK3與轉錄因子的活化有關。CCNE1基因編碼cyclin E1,CDKN1C編碼p57Kip2。一般而言,cyclin E1和周期蛋白依賴性激酶(cyclin-dependent kinase,CDK)2結合,成為細胞從G1進入S期的關鍵激酶復合物,磷酸化相關底物蛋白而發揮作用;而p57Kip2作為抑制因子,可結合cyclin E1-CDK2使其失去活性[8]。已有報道,p57Kip2和HCC的發生發展有關系,p57Kip2失活與低分化的HCC密切相關,是HCC預后的一個獨立預測因子[9]。本研究發現,索拉非尼作用PLC細胞株后,CDKN1C(p57)上調,CCNE1(cyclin E1)和CDK6下調,但索拉非尼影響p57-cyclin E1-CDK6細胞周期調控機制的具體過程,值得進一步研究。由于索拉非尼在應用過程中可發生耐藥,部分患者在應用的后期對索拉非尼失去反應,腫瘤因此發生進展,導致治療效果下降。因此,目前有提倡聯合用藥,提高索拉非尼治療HCC的療效的相關研究[10]。另有研究表明,miR-195在 85.7% 的HCC中顯著下調,而CDK6則是miR-195的一個作用靶點[11]。索拉非尼有無可能通過miR-195-CDK6機制抑制HCC細胞,有待進一步研究。目前關于microRNA、索拉非尼和 HCC三者關系的研究較少[12-13],而microRNA是未來藥物研究的一個方向。我們的實驗結果為未來的索拉非尼研究提供了相關的線索。
本研究發現索拉非尼作用后MAPK信號通路中的jun上調,這與最近Cervello等[14]的研究一致。他們運用全基因組芯片,利用索拉非尼作用HepG2細胞株,同樣發現jun基因上調;他們的研究還發現,索拉非尼作用后,HRK mRNA和c-Jun顯著上調且HepG2細胞發生凋亡,而HRK正是c-Jun作用的的一個基因靶點,另外抑制c-Jun信號通路可與索拉非尼發揮協同效應。本研究所采用的細胞系和Cervello等[13]的不同,但有相似的結果,提示本研究的實驗結果較為可靠,c-Jun上調可能是索拉非尼抑制HCC細胞時發生的共同現象;同時,也提示在索拉非尼抑制肝癌的研究領域中,有必要進一步對c-Jun信號通路在其中的具體機制進行研究。

表1 Sorafenib處理PLC細胞后MAPK信號通路的差異表達基因Table 1.Differential expression of MAPK signal genes in sorafenib-treated PLC cells
總之,本研究篩選出對索拉非尼敏感的HCC細胞系,并利用此敏感細胞系,通過MAPK信號通路基因芯片,鑒定出索拉非尼作用后MAPK信號通路中發生變化的具體分子,這些分子主要與細胞周期調控和轉錄因子活化相關,為進一步的研究提供了線索和方向。
[1] Abou-Alfa GK,Schwartz L,Ricci S,et al.Phase II study of sorafenib in patients with advanced hepatocellular carcinoma[J].J Clin Oncol,2006,24(26):4293-4300.
[2] Cheng AL,Kang YK,Chen Z,et al.Efficacy and safety of sorafenib in patients in the Asia-Pacific region with advanced hepatocellular carcinoma:a phase III randomised,double-blind,placebo-controlled trial[J].Lancet Oncol,2009,10(1):25-34.
[3] Llovet JM,Ricci S,Mazzaferro V,et al.Sorafenib in advanced hepatocellular carcinoma [J].N Engl J Med,2008,359(4):378-390.
[4] 闞和平,譚永法,周 杰.索拉非尼治療肝移植術后肝癌復發的療效與腫瘤組織Ki-67表達及微血管密度的關系[J].中國病理生理雜志,2011,27(8):1549-1551.
[5] Liu L,Cao Y,Chen C,et al.Sorafenib blocks the RAF/MEK/ERK pathway,inhibits tumor angiogenesis,and induces tumor cell apoptosis in hepatocellular carcinoma model PLC/PRF/5 [J].Cancer Res,2006,66(24):11851-11858.
[6] Tai WT,Cheng AL,Shiau CW,et al.Signal transducer and activator of transcription 3 is a major kinase-independent target of sorafenib in hepatocellular carcinoma[J].J Hepatol,2011,55(5):1041-1048.
[7] Chen KF,Chen HL,Tai WT,et al.Activation of phosphatidylinositol 3-kinase/Akt signaling pathway mediates acquired resistance to sorafenib in hepatocellular carcinoma cells[J].J Pharmacol Exp Ther,2011,337(1):155-161.
[8] Liang Y,Li WW,Yang BW,et al.Aryl hydrocarbon receptor nuclear translocator is associated with tumor growth and progression of hepatocellular carcinoma[J].Int J Cancer,2012,130(8):1745-1754.
[9] Nakai S,Masaki T,Shiratori Y,et al.Expression of p57KIP2in hepatocellular carcinoma:relationship between tumor differentiation and patient survival[J].Int J Oncol,2002,20(4):769-775.
[10] Huynh H,Ngo VC,Koong HN,et al.Sorafenib and rapamycin induce growth suppression in mouse models of hepatocellular carcinoma[J].J Cell Mol Med,2009,13(8B):2673-2683.
[11] Xu T,Zhu Y,Xiong Y,et al.MicroRNA-195 suppresses tumorigenicity and regulates G1/S transition of human hepatocellular carcinoma cells [J].Hepatology,2009,50(1):113-121.
[12] Bai S,Nasser MW,Hsu SH,et al.MicroRNA-122 inhibits tumorigenic properties of hepatocellular carcinoma cells and sensitizes these cells to sorafenib[J].J Biol Chem,2009,284(46):32015-32027.
[13] Shimizu S,Takehara T,Hikita H,et al.The let-7 family of microRNAs inhibits Bcl-xL expression and potentiates sorafenib-induced apoptosis in human hepatocellular carcinoma[J].J Hepatol,2010,52(5):698-704.
[14] Cervello M,Bachvarov D,Lampiasi N,et al.Molecular mechanisms of sorafenib actioin in liver cancer cells[J].Cell Cycle,2012,11(15):2843-2855.
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
天津醫科大學學報(2021年1期)2021-01-26 00:57:10
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
電子制作(2018年11期)2018-08-04 03:25:42
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國繼續醫學教育(2015年3期)2016-01-06 01:36:44
分子影像學雜志(2015年3期)2015-12-04 03:28:58
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
