結核疫苗研究進展
2013-10-09 06:11:28劉珊珊韓文瑜雷連成孫長江姚翠梅趙紅蕾
中國獸藥雜志 2013年3期
劉珊珊,韓文瑜,雷連成,馮 新,孫長江,姚翠梅,高 宇,趙紅蕾
(吉林大學農學部畜牧獸醫學院,長春 130062)
我國屬結核病高流行地區,結核病人數居全世界第二位,是全球22個結核病高負擔國家之一。世界衛生組織早在1993年宣布“全球結核病緊急狀態”。卡介苗(BCG)是牛型結核分枝桿菌強毒株經13年傳代培養獲得的減毒株,于1921年首次使用。自從1974年世界衛生組織正式將其列入免疫接種計劃以來,已經有約40億劑的卡介苗得到接種。目前卡介苗仍是唯一的結核疫苗,長期的臨床應用驗證其免疫效果大約在接種10~15后年逐漸減弱,對兒童以及青少年的保護性受到限制。臨床試驗也證實,BCG免疫不能對成年人提供良好的保護效果,更無法預防潛伏性結核感染。因此,臨床上迫切需要新的疫苗來控制結核病的發生發展。
結核疫苗研究的關鍵是對結核分枝桿菌(MTB)的致病機制有深入了解。MTB致病機理復雜,主要是誘導機體細胞免疫,因此結核病不能像其他疾病一樣應用傳統疫苗免疫手段得以有效控制,同時也不能用傳統免疫血清學方法進行有效診斷,目前為止在疫苗研制和檢測方面均未得到突破性進展。基于此,各國均在加大投入對結核病進行廣泛深入研究。本文對結核疫苗研制進展進行了闡述。
1 抗結核感染相關免疫反應
了解MTB感染免疫機制有利于合理設計抗結核疫苗。MTB是胞內感染菌,因其獨特的蠟質細胞壁結構,對于可溶性抗菌成分顯示出強烈抵抗作用,而肺泡內吞噬作用顯得尤為重要,機體抗MTB感染主要依賴細胞免疫應答,是一類T細胞介導的、以巨噬細胞為主要效應細胞的免疫反應,包括巨噬細胞吞噬MTB、抗原遞呈、T淋巴細胞對抗原的特異性識別結合、抗原增殖和分化、免疫細胞釋放相應細胞因子、巨噬細胞激活和殺菌等。調節性免疫反應對于結核感染至關重要,能夠提供長期的免疫保護以防止疾病復發。MTB潛伏感染后在機體免疫力下降的情況下常常會發生疾病復發甚至進入急性感染期。可以說機體免疫反應的強度很大程度上決定了帶菌感染者一生的健康。全世界每年20億結核感染者中,只有10%會發展為活動性結核,而剩下的90%無疑因為患者本身完善的免疫系統將疾病得以控制[1-2]。而免疫反應對于控制結核的有效作用在結核共感染HIV得到另一個方面的證實:共感染患者中一生中10%可能發展為活動性肺結核的幾率擴增到1年內就有10%可能發展為活動性肺結核[3]。
1.1 抗結核感染相關細胞免疫反應 MTB進入機體后被肺吞噬細胞和樹突狀細胞等抗原遞呈細胞吞噬,后者將抗原處理后遞呈給MHC限制性T細胞[4]。MHC II分子將抗原遞呈給CD4+T細胞,而CD4+T細胞輔助細胞分化成不同亞群,激發產生相應細胞因子,通常產生IFN-γ、IL-2和TNF-α來進一步激活巨噬細胞。而MHC I類分子將相關抗原遞呈給CD8+T輔助細胞,進而分化成IFN-γ和TNF-α來激活巨噬細胞[5],因MTB表面富含補體受體,感染部位瀕死的巨噬細胞吸引著單核細胞和其他免疫細胞從而形成局部損傷。IFN-γ信號同時還能促進吞噬體和溶酶體的融合,從而達到抵抗MTB的作用。IFN-γ對于抵抗結核等胞內感染是必不可少的,因其能激活巨噬細胞,并產生毒性分子從而殺死MTB。激活的巨噬細胞分泌諸如TNF-α,IL-12,IL-18等細胞因子。TNF-α對于誘發其他炎癥趨化因子和相應受體是非常有效的,因此TNF-α和IFN-γ是關鍵細胞因子,能促進感染性吞噬過程同時也作為評價TH1反應的標準[6-7]。而 TNF-α 本身作為一個效應因子可以協同IFN-γ來激活記憶T細胞是機體預防細胞內病原菌再次感染的基礎。而IL-2的共表達對于抵抗病毒感染從而保護宿主而顯得尤為重要(圖1)[8-9]。MTB 在巨噬細胞和 DC 細胞中得以存活,對于結核的有效控制主要依靠巨噬細胞和T細胞亞群[10],誘導記憶T細胞產生是高效疫苗必備條件。
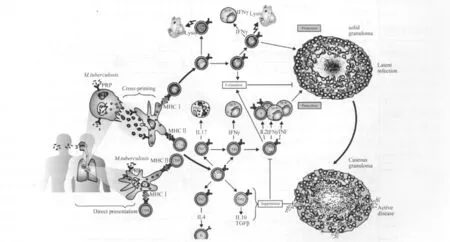
圖1 抗MTB感染相關的T細胞亞群[8]
1.2 抗結核感染相關體液免疫反應 針對MTB產生的抗體,由于無法接觸到胞內菌體,只能對釋放出巨噬細胞的細菌發揮作用,因此體液免疫反應只能起到輔助抗感染作用。體液免疫反應對于抗結核感染的作用仍沒有得到充分了解,最初研究中發現B細胞缺失的小鼠相對于野生鼠并未顯示出高易感性。然而近期研究不斷發現,在這些B細胞缺陷小鼠中,MTB滴度和因聚集過多的中性粒細胞而出現的病理損傷相對高于野生菌[11]。而抗體在某些情況下對于抵抗MTB感染顯示一定作用,在肺結核開放期,MTB因表面具有宿主抗MTB抗體從而改變其吞噬細胞內化機制,使巨噬細胞趨化作用提高,增強其吞噬溶酶體作用而有效殺死MTB。
1.3 MTB是通過氣溶膠形式傳播感染 多數病灶集中在感染者肺部,因此有效地開發區域黏膜免疫從而控制結核傳播顯得意義深重。一些研究已經驗證了黏膜免疫對于抗結核感染的顯著作用[12-13],而該免疫反應通常涉及較為復雜的免疫機制,包括先天免疫、體液細胞免疫等,同時分泌到肺部黏膜的抗菌肽抗菌機制也參與其中。
2 結核疫苗研制策略
為了減少全球結核病的廣泛蔓延,不斷研發新的疫苗策略,以期達到對成年人和兒童都能提供優于BCG疫苗的免疫保護作用。主要研發策略見圖3。后續疫苗不僅能夠控制最初感染,而且能夠減少MTB的復制代謝活性,從而將其引入休眠期[14-15]。因此,合適的感染后疫苗也是研發的熱點,尤其在高感染率地區,這樣的疫苗顯得尤為必需。
圖3 結核疫苗研發策略[11]
2.1 重組BCG疫苗和減毒活性結核疫苗 重組BCG疫苗是將一些重要抗原尤其是MTB表面蛋白相關毒力因子導入BCG菌株,如存在于BCG缺失區域RD1、RD2等一些具有高度免疫活性抗原基因,從而得到一個重組BCG疫苗,以期能取代傳統BCG 疫 苗,如 rBCGΔureC:Hly(VPM1002)和rBCG30目前已經進入臨床前期。而MTB減毒活疫苗是敲除MTB的一些基因從而得到基因缺失突變株,MTB減毒活疫苗因其能夠表達多種抗原從而能激活多功能T細胞群亞群,顯示出較強的免疫活性。減毒活疫苗比BCG具有更好的保護作用和安全性[16-17],目前主要的結核重組疫苗以及減毒活疫苗見表1。而其他一些減毒疫苗期待在未來幾年內陸續進入臨床前期,主要目標設置在基因敲除方面,比如MTBVAC01疫苗是通過敲除轉錄調控子phoP相關毒力基因而構建的[18]。

表1 MTB重組疫苗和減毒疫苗[16]
2.2 亞單位疫苗和活載體疫苗 對MTB亞單位疫苗研究起源于20世紀80年代,研究發現只有活的MTB才能夠提供免疫保護作用,而死菌則不能,原因在于死菌中缺乏一些毒力因子,而這些毒力因子常常是一些分泌蛋白,并且通過實驗證實,這些分泌蛋白往往通過佐劑能夠刺激機體產生強烈的免疫反應[19-20]。盡管隨著蛋白組學發展,不斷更新新的方法來分析MTB早期培養濾液,將近有100多種新的表面蛋白被發現[21],但目前主要亞單位疫苗候選抗原仍集中在少數幾個蛋白上,如Ag85B、ESAT-6和CFP-10等,顯然目前對于亞單位疫苗候選基因篩選工作只做了很小的一部分,大量工作等待我們去開展。目前MTB亞單位疫苗主要有兩種類型,一種是由2~3個MTB主要毒力相關蛋白構成;另一種是表達一個或是多個MTB蛋白的活載體疫苗。重組蛋白疫苗需要相關佐劑輔助來刺激產生Th1型免疫反應,而活載體疫苗則不需要相關的佐劑,目前研發的活載體疫苗能產生類似BCG免疫相關的免疫保護作用,甚至有些對于靈長類動物顯示出良好的免疫保護作用[22]。Ag85B-ESAT-6、Ag85B -TB -10.4(H4,AERAS-404)、M72、MVA85A、Ad35(AERAS -402)等都是這兩個類型的候選疫苗[23-26],有的也進入臨床一期。活載體疫苗技術的發展促使產生了更多的疫苗設計新思路,將痘病毒修飾后做為DNA載體來重組重要抗原,聯合BCG和禽痘重組Ag85A疫苗能激發出比BCG更強的免疫保護力。r-MVAAg85疫苗已經進入臨床一期階段。利用鼠傷寒沙門氏菌減毒株作為載體,將編碼MTB保護性抗原基因導入其中,可構建重組st疫苗,如rst-Ag85B;而將保護基因導入小鼠巨噬細胞或骨髓細胞則可構建結核活細胞疫苗,如J774-HSP65疫苗。目前主要的結核亞單位疫苗見表2。

表2 MTB亞單位疫苗[24]
2.3 DNA疫苗 向宿主體內注射DNA載體使宿主通過自身包裝表達機制來表達DNA載體中融合蛋白,從而避免了使用佐劑的副作用,這些抗原通過遞呈MHC I類相關分子,而誘導CD4+和CD8+T細胞免疫反應得到廣泛關注[27-28]。結核 DNA疫苗研究領域中 Ag85家族研究最為廣泛,其中Ag85A活性最強[29]。有研究甚至建立多價 DNA疫苗,融合 Ag85B、ESAT - 6、KatG、MPT8.4、MPT12、MPT63、MPT64 和 MPT83 蛋白[30]。目前主要的結核DNA疫苗見表3。

表 3 MTB DNA 疫苗[31]
有研究發現,分泌蛋白對于誘導DNA疫苗有效的保護作用是必需的,同時這種DNA疫苗對于高致病性耐藥臨床結核分離菌株有高效的保護作用[31]。Ag85A DNA疫苗也可以作為免疫增強劑,甚至是細胞因子刺激劑來提高機體免疫力。比如在給小鼠免疫BCG之前給予Ag85A DNA疫苗免疫,保護效果從單獨BCG免疫的0.67log到聯合免疫后的 1.12log,保護效果顯著增強[32]。有研究還將IL-12基因融合到Ag85A DNA疫苗中,從而將Ag85A DNA保護效率提高到與BCG免疫效果相當[33]。DNA疫苗的應用除了本身的安全問題,我們還必須考慮到這種類型疫苗是否可以應用到人類身上,而病毒編碼Ag85A疫苗對于BCG免疫的增強效果在人類身上也得到證實,這無疑是個好的開始,DNA疫苗對于人類至少可以成為有前景的免疫增強劑,而對于這種疫苗本身的恐懼并不足以抹殺其對人類免疫提高的顯著作用。
2.4 滅活疫苗 滅活菌體疫苗感染相關疾病的危險性相對較小,目前結核疫苗研制在這個領域主要采取兩個策略,一是滅活BCG載體疫苗,二是選用一些其他分枝桿菌載體,如熱滅活弱毒性分枝桿菌如Mycobacterium vaccae,具有安全性同時還能作為抗結核的免疫治療劑,而滅活Mycobacterium vaccae疫苗已經完成臨床3期安全實驗。臨床試驗顯示,這類型疫苗能夠提高放射療法和化學療法的療效,大大縮短治療時間[33]。
2.5 佐劑研發 佐劑的功能在過去疫苗研制過程中有所忽視,最近很多研究表明佐劑除了可以增強疫苗免疫原性(如DC靶向提呈),促進免疫細胞分化成熟,還在誘導免疫應答傾向(Th1或Th2),甚至改變特異性T細胞克隆群等過程中都發揮了重要作用,這對于抗結核感染顯得意義重大。在過去的幾年里,實驗驗證了一些佐劑作為結核疫苗有效佐劑所具備的有效性和安全性[35],其中單磷酸酯質A(MPL)被廣泛應用,尤其與DNA疫苗聯合使用時顯示更強的保護作用,而新一代的氨基烴基類MPL佐劑的藥效研究正積極展開。目前在結核疫苗研制領域由此也掀起了一場開發新型佐劑的激烈戰。
2.6 未來結核疫苗研究方向 未來結核疫苗研究主要著重兩個方向,一是感染后疫苗,靶向潛伏期MTB,以期來完全消滅潛伏期MTB,或是降低其復制感染幾率,從而控制疾病的復發,而潛伏表達相關抗原就成為該類疫苗的候選基因[36]。另一個方向就是開發BCG免疫增強劑,以期增強BCG免疫效果,而篩選的增強劑候選基因必須是能完全消滅MTB的相關抗原,而不是僅僅將其靶向潛伏期。
3 新的疫苗評價標準
對于疫苗評價主要是針對已經進入臨床前期的相關疫苗(表4),通過深入了解MTB相關免疫機制和致病機理,我們必須考慮到這些疫苗的有效性和安全性。有效性至少要比BCG強,安全性至少可以用于HIV高流行地區,而具體的評定標準見表5。

表4 處于臨床階段的結核疫苗及其接種方法[36]

表5 結核疫苗評價標準[36]
4 展望
隨著來自公共基金、政府機構和各項社會投資的增加,MTB致病機理、疫苗研發、疾病診斷等方面研究的不斷深入,預計在2015年以前會有20種左右新結核疫苗進入臨床一期試驗,到2015年會獲得一種有效、安全、廉價的新結核疫苗,得以在全球廣泛使用。我們有理由相信,經過數代科學家們持之以恒的努力,到2050年有可能達到控制和逐步消滅結核病的偉大目標[37]。
[1] Young D DYE C.The development and impact of tuberculosis vaccines[J].Cell,2006,124(4):683 -687.
[2] Kaufmann SH E.Recent findings in immunology give tuberculosis vaccines a new boost[J].Trends in Immunology,2005,26(12):660-667.
[3] Kaufmann SH E,Mcmichael A J.Annulling a dangerous liaison:Vaccination strategies against AIDS and tuberculosis[J].Nature Medicine,2005,11(5):578 -578.
[4] Dorhoi A,Kaufmann S H E.Fine - tuning of T cell responses during infection[J].Current Opinion in Immunology,2009,21(4):367-377.
[5] Kaufmann SH E.Essay- Envisioning future strategies for vaccination against tuberculosis[J].Nature Reviews Immunology,2006,6(9):699 -704.
[6] Newport M J,Huxley C M,Huston S,et al.A mutation in the interferon-gamma-receptor gene and susceptibility to mycobacterial infection[J].N Engl JMed,1996,335(26):1941 -1949.
[7] Flynn JL,Goldstein M M,CHAN J,etal.Tumor necrosis factoralpha is required in the protective immune response against Mycobacterium tuberculosis in mice[J].Immunity,1995,2(6):561-572.
[8] Kaufmann SH,Hussey G,Lambert PH.New vaccines for tuberculosis[J].The lancet,2010,375(5):2110 -2119.
[9] Foulds K E,Wu C Y,Seder R A.Th1 memory:Implications for vaccine development[J].Immunological Reviews,2006,211:58-66.
[10] Russell D G.Who puts the tubercle in tuberculosis?[J].Nat Rev Microbiol,2007,5(1):39 - 47.
[11] De valliere S,Abate G,Blazevic A,et al.Enhancement of innate and cell-mediated immunity by antimycobacterial antibodies[J].Infection and immunity,2005,73(10):6711 -6720.
[12] Goonetilleke N P,Mcshane H,Hannan C M,et al.Enhanced immunogenicity and protective efficacy against Mycobacterium tuberculosis of bacille Calmette-Guerin vaccine using mucosal administration and boosting with a recombinantmodified vaccinia virus Ankara[J].Journal of Immunology,2003,171(3):1602 -1609.
[13] Teitelbaum R,Glatman - freedman A,CHEN B,et al.A mAb recognizing a surface antigen of Mycobacterium tuberculosis enhances host survival[J].Proc Natl Acad Sci U S A,1998,95(26):15688-15693.
[14] VoskuilLM I,Schnappinger D,Visconti K C,et al.Inhibition of respiration by nitric oxide induces a Mycobacterium tuberculosis dormancy program[J].Journal of Experimental Medicine,2003,198(5):705-713.
[15] Betts JC,Lukey P T,Robb L C,et al.Evaluation of a nutrient starvation modelof Mycobacterium tuberculosis persistence by gene and protein expression profiling[J].Molecular Microbiology,2002,43(3):717 -731.
[16] Torchinsky M B,Garaude J,Martin A P,et al.Innate immune recognition of infected apoptotic cells directs T(H)17 cell differentiation[J].Nature,2009,458(7234):78 -82.
[17] Winau F,Weber S,Sad S,et al.Apoptotic vesicles crossprime CD8 T cells and protect against tuberculosis[J].Immunity,2006,24(1):105 -117.
[18] Martin C,Williams A,Hernandez- pando R,et al.The live Mycobacterium tuberculosis phoPmutant strain ismore attenuated than BCG and confers protective immunity against tuberculosis in mice and guinea pigs[J].Vaccine,2006,24(17):3408 -3419.
[19] PalP G Horwitz M A.Immunization with extracellular proteins of Mycobacterium tuberculosis induces cell-mediated immune responses and substantial protective immunity in a guinea pig model of pulmonary tuberculosis[J].Infect Immun,1992,60(11):4781-4792.
[20] C a- z,IS,Jm G,et al.The secreted antigens of Mycobacterium tuberculosis and their relationship to those recognized by the available antibodies[J].J Gen Microbiol,1988,134(2):531-538.
[21] Covert B A,Spencer J S,Orme I M,et al.The application of proteomics in defining the T cell antigens of Mycobacterium tuberculosis[J].Proteomics,2001,1(4):574 -586.
[22] Verreck F A W,Vervenne R A W,Kondova I,et al.MVA.85A boosting of BCG and an attenuated,phoP deficient M.tuberculosis vaccine both show protective efficacy against tuberculosis in rhesusmacaques[J].PLoSOne,2009,4(4):e5264.
[23] Von eshen K,Morrison R,BraunM,et al.The candidate tuberculosis vaccine Mtb72F/AS02A Tolerability and immunogenicity in humans[J].Human Vaccines,2009,5(7):475 -482.
[24] Lingnau K,RiedlL K,Von Gabain A.IC31(R)and IC30,novel types of vaccine adjuvant based on peptide delivery systems[J].Expert Review of Vaccines,2007,6(5):741 -746.
[25] Dietrich J,Aagaard C,Leah R,et al.Exchanging ESAT6 with TB10.4 in an Ag85B fusion molecule - based tuberculosis subunit vaccine:Efficient protection and ESAT6-based sensitive monitoring of vaccine efficacy[J].Journal of Immunology,2005,174(10):6332-6339.
[26] McshaneH,PathanAA,SanderCR,etal.Recombinantmodified vacciniavirusAnkaraexpressingantigen85AboostsBCG-primed andnaturallyacquiredantimycobacterialimmunityinhumans[J].NatureMedicine,2004,10(12):1397 -1397.
[27] KirmanJR,SederRA.DNAvaccination:theanswertostable,protectiveT - cellmemory[J].CurrentOpinioninImmunology,2003,15(4):471 -476.
[28] HuygenK.OntheuseofDNAvaccinesfortheprophylaxisof mycobacterialdiseases[J].InfectionandImmunity,2003,71(4):1613-1621.
[29] UlmerJB.TuberculosisDNAvaccines[J].ScandinavianJournal ofInfectiousDiseases,2001,33(4):246 -248.
[30] LiZM,HowardA,KelleyC,etal.ImmunogenicityofDNAvaccines expressingtuberculosisproteinsfused to tissue plasminogen activatorsignalsequences[J].InfectionandImmunity,1999,67(9):4780-4786.
[31] BaldwinSL,DsouzaCD,OrmeIM,etal.Immunogenicityand protectiveefficacyofDNAvaccinesencodingsecretedandnonsecretedformsofMycobacteriumtuberculosisAg85A[J].Tuber LungDis,1999,79(4):251 -259.
[32] FengCG,alendiraU,DemangelC,etal.PrimingbyDNA immunizationaugmentsprotectiveefficacyofMycobacteriumbovis bacilleCalmette - Guerinagainsttuberculosis[J].Infectionand Immunity,2001,69(6):4174 -4176.
[33] KamathAT,HankeT,BruscieH,etal.Co - immunizationwith DNAvaccinesexpressinggranulocyte-macrophagecolonystimulatingfactorandmycobacterialsecretedproteinsenhances T -cellimmunity,butnotprotectiveefficacyagainstMycobacterium tuberculosis[J].Immunology,1999,96(4):511 -516.
[34] DlugovitzkyD,FirenzaG,FarroniM,etal.Immunologicalconsequencesofthreedosesofheat-killedMycobacteriumvaccaein theimmunotherapyoftuberculosis[J].RespiratoryMedicine,2006,100(6):1079 -1087.
[35] LindbladEB,ElhayMJ,SilvaR,etal.Adjuvantmodulationof immuneresponsestotuberculosissubunitvaccines[J].Infect Immun,1997,65(2):623 -629.
[36] DanielFH.Tuberculosisvaccinedevelopment:Goals,immunological design,andevaluation[J].Thelancet,2008,372(5):164 -175.
[37] DraftstrategicplanofthestoppartnershipworkinggrouponTB vaccinedevelopment.2ndgilbalplantostopTB[J].GPIIStatus 2005:2006-2015.
