貴州省煙草黑脛病菌對烯酰嗎啉的敏感性
2013-09-28 01:33:52桑維鈞張新強潘滴滴
植物保護 2013年1期
關鍵詞:煙草
蘇 凱, 桑維鈞*, 張新強, 潘滴滴, 王 慧
(1.貴州大學農學院,貴州山地農業病蟲害重點實驗室,貴陽 550000;2.甘肅省天水市秦州縣林業局,天水 741000;3.貴州省鎮遠縣農業局,鎮遠 557700)
煙草黑脛病是由煙草疫霉煙草變種(Phytophthoranicotianaevar.nicotianae)引起的毀滅性病害,是世界煙草生產中危害最為嚴重的病害。該病在我國各煙草產區大量發生,是僅次于煙草病毒病的病害,嚴重影響了煙草的產量和品質。該病害通常在潮濕、多雨條件下發病重,潛育期短,再次侵染次數多,傳播與蔓延迅速,嚴重時煙株常成片死亡。目前,防治煙草黑脛病主要通過采用抗病品種、栽培防治、化學防治等措施,其中化學防治是最普遍且實用的方法,但是病菌極易對殺菌劑產生抗性。
烯酰嗎啉是防治疫霉屬病害的高效內吸性殺菌劑,其作用為保護、治療和抑制孢子產生[1],對農作物上產生的霜霉病、疫霉病有很好的防治作用,其防治范圍為馬鈴薯、番茄晚疫病,黃瓜、油菜霜霉病和辣椒疫病,煙草黑脛病等由鞭毛菌亞門卵菌綱引起的病害。該藥對卵菌綱的作用特點是首先影響細胞壁分子結構的重新排列,然后干擾病原菌細胞壁聚合體的組裝,從而干擾細胞壁的形成和促使孢子囊壁的分解,使病原菌出現自動死亡[2],因此分析研究對貴州省煙草黑脛病病菌烯酰嗎啉的敏感性,明確貴州省各煙草產區抗藥菌的分布和抗藥性水平,為煙草黑脛病的防治提供科學依據具有重要的實踐意義。
1 材料與方法
1.1 病原菌的分離與純化
將貴州省境內各煙區采集的新鮮煙草黑脛病標本用自來水沖洗干凈,晾干,剖開莖基部發病部位,取病健交界處的病組織切成2 mm×2 mm左右大小,沿培養皿周緣直接置于含抗菌素的V8汁選擇性培養基表面,每皿(直徑9 cm)6~10塊。在28℃下培養3~7 d,待菌落形成后,進行鏡檢,選擇菌絲無隔膜的繼續培養,待產生孢子囊后,鏡檢觀察,鑒定。然后再從菌落邊緣切取2 mm×2 mm的菌絲塊轉移至盛有V8培養基的試管斜面上,25℃黑暗培養7 d后,常溫保存備用。
按照鄭小波的方法[3],將產生有大量孢子囊的菌絲或菌絲塊挑入盛有10 mL左右滅菌自來水的培養皿中,培養皿置于4~10℃冰箱中10 min左右后,取出置于適溫(20~25℃)下20 min,在顯微鏡下可以看到有大量游動孢子釋放。在冷卻凝固的0.2%V8培養基平板上(培養皿直徑9 cm)滴加游動孢子懸浮液100~200μL,約含游動孢子200~500個,均勻涂布于培養基平板表面。培養皿置于室溫下(25℃左右)、黑暗中培養24 h左右,使游動孢子休止萌發,并在培養基上形成微小菌落。并在顯微鏡下觀察;然后在顯微鏡下用滅菌刀片切取帶有單個已萌發游動孢子的瓊脂塊于選擇性培養基平板上,在25℃黑暗條件下培養2~3 d后可獲得單游動孢子的無性系。
1.2 煙草黑脛病菌菌絲生長對烯酰嗎啉的敏感性測定
供試烯酰嗎啉由江蘇輝豐農化股份有限公司生產,有效成分為50%水分散粒劑;先用無菌水配成濃度為10 mg/m L的母液備用。
將黑脛病菌接種于V8培養基上,25℃下黑暗培養5 d后,用直徑5 mm的滅菌打孔器在近菌落邊緣處打孔。取適量已配好的烯酰嗎啉母液,加入融化后即將凝固的V8培養基中,混勻,分別配制成濃度為0、0.5、1.0、2.0、4.0μg/m L V8培養基平板。待培養基冷凝后用接種針挑取菌碟,菌面向下接于含烯酰嗎啉的培養基中心,每皿1塊。接種后放入恒溫箱中,每處理4皿,試驗重復3次,25℃下培養5 d后用十字交叉法測量菌落直徑。
按下式計算不同濃度藥劑對黑脛病菌菌絲生長的抑制率:

計算抑制率幾率值和濃度對數值并建立毒力回歸方程,計算烯酰嗎啉對黑脛病菌各菌株的抑制中濃度EC50及相關系數R[4]。數據統計用農藥室內生物測定數據處理系統(武漢市蔬菜科學研究所,農業部農藥檢定所)。對42個菌株的EC50用SPSS程序進行正態分布檢驗,是否符合正態分布。根據王文橋等[5]的方法將菌株分為抗性菌株(EC50>10μg/m L)、中間型菌株(0.1μg/m L≤EC50≤10μg/m L)和敏感菌株(EC50<0.1μg/m L)。
1.3 煙草黑脛病菌游動孢子囊形成對烯酰嗎啉的敏感性測定
參照Matheron等[6]的方法,從生長在V8平板上的黑脛病菌落邊緣處打直徑為5 mm的菌碟4塊,然后分別置于含有0.0、0.5、1.0、2.0、4.0μg/mL烯酰嗎啉的9 cm培養皿中(加入藥液時要使液面剛剛沒過菌碟塊),25℃下光暗交替培養4 d后倒去無菌水,在菌碟上滴加少量酸性品紅,1 min之后從菌碟的一側慢慢滴加乳酸直至洗去大量品紅[7]。在顯微鏡下可以看到游動孢子囊呈現紅色,在菌碟上隨機選取3個視野,在顯微鏡下對游動孢子囊進行計數(包括少量未被染紅的游動孢子已出來的孢子囊),每處理3皿,重復3次,數據處理方法同菌絲生長抑制試驗,計算菌株的EC50并進行正態分布檢驗。
2 結果與分析
2.1 煙草黑脛病菌的分離
從貴州省興仁市屯腳鎮、雨樟鎮,興義烏沙鎮、白碗窯,晴隆縣光照鎮,開陽縣羊場鎮,福泉市黎山鄉,余慶縣松煙鎮、熬溪鎮,湄潭縣抄樂鄉、興隆鎮,遵義縣三岔鎮,銅仁市,大方縣雙山鎮,納雍縣,長順縣,安龍縣,水城縣,黔西縣林泉鎮,畢節市等煙草黑脛病經常發生地區采集發病樣本,經分離、純化和鑒定后,總共得到47個煙草黑脛病菌菌株。
2.2 煙草黑脛病菌對烯酰嗎啉的敏感性水平
2.2.1 煙草黑脛病菌菌絲生長對烯酰嗎啉的敏感性
采用菌絲生長速率法測定單孢分離得到的47個單孢菌株的EC50,結果(表1)發現大多數菌株屬于中間型菌株,煙草黑脛病菌自然種群對烯酰嗎啉沒有明顯的抗藥性。
在黑脛病菌對藥劑的敏感性變化范圍內,將EC50等分為8個階段,統計出每個階段菌株的個數和頻率,然后以每個階段EC50的中值為橫坐標,頻率為縱坐標,作出頻率分布圖(圖1),圖中的峰代表病原菌群體對藥劑的敏感性分布,發現煙草黑脛病菌絲對烯酰嗎啉的敏感性呈單峰曲線,采用SPSS敏感性分布進行了K-S正態性檢驗,得到Z=0.784,P=0.571,表現連續性,則表明47個煙草黑脛病菌株對烯酰嗎啉敏感性的頻率分布符合正態分布。

表1 煙草黑脛病菌菌絲生長對烯酰嗎啉的敏感性(貴州,2011年)Table 1 Sensitivity of mycelial growth of Phytophthora nicotianae var.nicotianae to dimethomorph(Guizhou,2011)
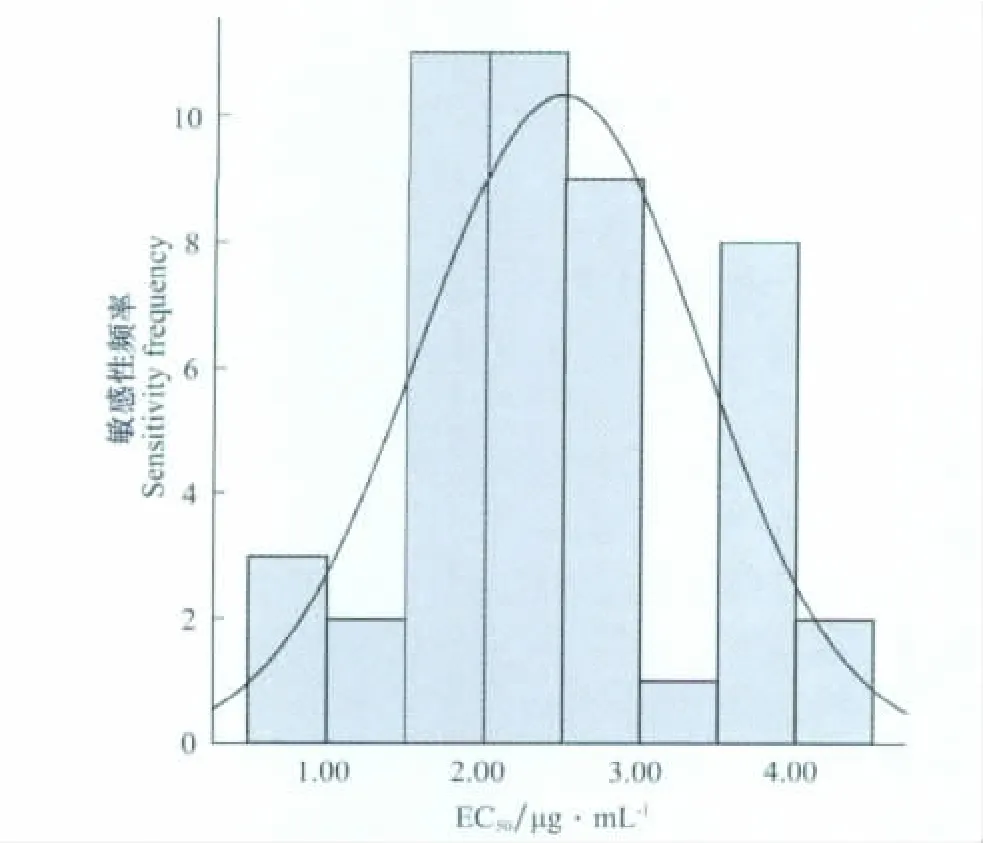
圖1 煙草疫霉病菌47個菌株菌絲生長對烯酰嗎啉敏感性的頻率分布Fig.1 Frequency distribution of sensitivity of 47 P.nicotianae var.nicotianae isolates to dimethomorph
2.2.2 不同地區煙草黑脛病菌菌株對烯酰嗎啉敏感性差異
采用Duncan氏新復極差法檢驗了烯酰嗎啉對47個不同地區菌株EC50值的差異顯著性。結果顯示:47個菌株之間EC50值有極顯著性差異(見表2)。EC50最高為興仁屯腳的TJ1菌株,為4.232 7μg/mL,而開陽的KY2菌株EC50最低,只有0.792 6μg/mL,最高值與最低值相差5.34倍,供試47個菌株的平均EC50為2.476 8μg/m L(見表1)。表明貴州不同地區的黑脛病菌菌株對烯酰嗎啉的敏感性不同。當烯酰嗎啉的濃度為0、0.5、1.0、2.0、4.0μg/m L時,對耐藥性最強菌株TJ1的抑制率分別為9.28%、21.20%、36.72%、46.00%,而對最敏感菌株 KY2的抑制率分別為:47.40%、51.39%、53.65%、60.59%,差異顯著。

表2 烯酰嗎啉對不同地區煙草黑脛病菌菌株EC50值的比較1)Table 2 The EC50 values of dimethomorph to different isolates of P.nicotianae var.nicotianae
2.2.3 不同地理來源菌株對烯酰嗎啉敏感性水平的系統聚類分析
烯酰嗎啉對47個供試菌株EC50值的離差平方和系統聚類分析結果(圖2)表明:47個菌株的EC50值可分為由低到高的4個聚類組,第1組包括11個菌株(林泉2個,烏沙2個,屯腳3個,興隆2個,遵義三岔1個);第2組包括3個菌株(開陽2個,抄樂的1個);第3組包括10個菌株(水城2個,長順1個,遵義三岔1個,大方雙山1個,屯腳2個,白碗窯1個,松煙1個,雨樟1個);第4組包括23個菌株(雨樟1個,安龍2個,大方雙山2個,福泉3個,畢節1個,白碗窯3個,松煙3個,晴隆2個,長順2個,光照2個,抄樂1個)。大部分菌株出現在第3、4聚類組中,只有少部分菌株單獨成一類,表明菌株對烯酰嗎啉的敏感性和其地理來源有一定的相關性,從各系列菌株對烯酰嗎啉敏感性的變化程度看,福泉、大方雙山最大,林泉、開陽、屯腳、烏沙、興隆的變化幅度最小,與Duncan新復極差法的分析結果一致[8]。
2.2.4 煙草黑脛病菌游動孢子囊形成對烯酰嗎啉的敏感性
烯酰嗎啉對47個煙草黑脛病菌游動孢子囊形成的EC50為0.394 5~4.625 3μg/m L(表3),平均EC50為2.040 0μg/m L。不同濃度的烯酰嗎啉對煙草黑脛病菌孢子囊形成的影響表明:當烯酰嗎啉濃度較低時,對不同菌株的產孢抑制率差異明顯,如當烯酰嗎啉濃度為0.5μg/m L時對菌株BWY3的抑制率為26.3%,而對菌株FQ4的抑制率則為40.4%。隨著烯酰嗎啉濃度的提高,菌株之間對烯酰嗎啉的敏感性差異縮小。當烯酰嗎啉濃度為8μg/m L時,其對煙草黑脛病菌的產孢抑制率已達100%,表明其可以完全抑制孢子囊的形成,隨著處理濃度的提高,其抑制率差異逐漸縮小,結果見表3。

圖2 不同地理來源菌株對烯酰嗎啉敏感性水平的系統聚類分析Fig.2 Systemic cluster analyses of sensitivity level of isolates with different geographical origin to dimethomorph
采用SPSS敏感性分布進行了K-S正態性檢驗,得Z=1.127,P=0.076,同樣表明游動孢子囊對烯酰嗎啉的敏感性呈現出正態分布,結果表明烯酰嗎啉對黑脛病菌的游動孢子囊有抑制作用,比對黑脛病菌菌絲的活性低或大致相等,但也出現了值相反的情況,其中BWY3、FQ1、KY1、QL1和全部的CL系列對游動孢子的EC50值明顯大于對菌絲的,初步判斷煙草黑脛病菌部分菌株的游動孢子已在低濃度下產生了耐藥性。

表3 煙草黑脛病菌游動孢子囊形成對烯酰嗎啉的敏感性(貴州,2011年)Table 3 Sensitivity of zoosporangia formation of P.nicotianae var.nicotianae to dimethomorph(Guizhou,2011)

圖3 煙草黑脛病菌47個菌株游動孢子囊形成對烯酰嗎啉敏感性的頻率分布Fig.3 Sensitivity frequency distribution of zoosporangia formation of 47 P.nicotianae var.nicotianae isolates to dimethomorph
3 結論與討論
試驗結果表明:絕大部分菌株為烯酰嗎啉一般耐藥型,表明貴州煙草黑脛病菌對烯酰嗎啉已產生了不同程度的耐藥性,推測有2種可能:一種是該地區煙田間存在天然的耐藥性菌株;另一種是由于煙農單一使用烯酰嗎啉,導致病原菌產生適應性變異而出現了耐藥性群體。烯酰嗎啉作為生產上單一使用防治煙草黑脛病的藥劑具有一定的風險性,建議采用新型殺菌劑,同時開展其他殺菌劑的敏感性試驗,為減少病菌抗藥性的產生提供理論依據。
對來自貴州省主要煙區47個菌株抗性的差異顯著性檢驗結果及菌株的抗性系統聚類分析結果均表明,不同地區的煙草黑脛病菌對烯酰嗎啉敏感性存在顯著差異。
煙草黑脛病發病初期病部在高濕條件下可形成大量游動孢子囊并釋放游動孢子,所以在田間易形成發病中心。游動孢子囊和游動孢子都可成為再侵染源侵染健康煙株。試驗結果表明,烯酰嗎啉對黑脛病菌菌絲生長的抑制效果與對游動孢子囊的抑制效果基本相同或略好,當烯酰嗎啉的濃度達到8.0μg/mL時可以完全控制煙草黑脛病菌孢子囊的形成。因此,發病初期,在田間使用8.0μg/m L以上濃度的烯酰嗎啉可及時對游動孢子囊及游動孢子的萌發起抑制作用,進而有效地抑制病原菌的再侵染,并對病害的傳播和流行起到較好的控制作用。
[1]黃青春,葉鐘音.烯酰嗎啉(DMM)的特性及其作用機制[J].農藥科學與管理,2000,21(5):28-31.
[2]Stein J M,Kirk W W.The generation and quantification of resistance to dimethomorph inPhytophthorainfestans[J].Plant Disease,2004,88(9):930-934.
[3]鄭小波,陸家云.建立掘氏疫霉單游動孢子無性系方法的研究[J].南京農業大學學報,1991,14(4):135.
[4]趙善歡.植物化學保護[M].第2版.北京:農業出版社,1983:518-520
[5]王文橋,馬志強,張小風,等.致病疫霉抗藥性、交配型和適合度[J].植物病理學報,2002,32(3):278-283.
[6]Matheron M E,Porchas M.Impact of azoxystrobin,dimethom orph,fluazinam,fosetylal,and metalaxyl on growth,sporulation,and zoospore cyst germ ination of threePhytophthoraspp.[J].Plant Disease,2000,84(4):454-458.
[7]鄭小波.疫霉菌及其研究技術[M].北京:中國農業出版社,1997.
[8]馬國勝.煙草黑脛病菌生理生態及對甲霜靈抗性監測與遺傳研究[D].合肥:安徽農業大學,2002:86-87.
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
