2010-2011年度四省小麥區試品種遺傳多樣性和抗條銹性分析
2013-09-28 01:44:22王冬梅王鳳濤藺瑞明徐世昌
植物保護 2013年1期
王冬梅, 馮 晶, 王鳳濤, 藺瑞明, 徐世昌
(中國農業科學院植物保護研究所,植物病蟲害生物學國家重點實驗室,北京 100193)
隨著近代育種的開展,育種家們過多對產量、抗病性、品質等的關注,造成育種的遺傳基礎日益變窄,品種的遺傳多樣性逐年下降[1],給育種工作帶來了重要的瓶頸。對于遺傳多樣性的研究,前人有很多報道,可以從表型性狀[2-3]、生化標記[4-5]、分子標記[6-8]等方面進行研究。但這些一方面耗時比較長,另一方面就是缺乏對整個基因組進行研究,會受到抽樣誤差的影響。親緣系數(coefficient of parent-age)是用于度量兩個個體間親緣關系的一個指標,等于兩個個體的基因組中相同且同源基因的比例,其不受環境因素的影響,已有研究表明,對于已知系譜信息的自花授粉植物小麥,COP分析是一種簡便的評價遺傳多樣性方法[9],近年來,根據長期育種進程中所形成的系譜信息,用親緣系數度量品種間的親緣關系,前人已獲得一些成果。徐曉丹等[10]利用親緣系數對57個河南小麥主栽品種進行分析,探討了河南小麥主栽品種的遺傳信息豐富度;王江春等[11]對山東省的66個小麥品種的親緣系數進行分析,探討了新中國成立以來山東省小麥品種的遺傳多樣性。Souza和 Mercado都指出[12-13],當有完整系譜信息時,系譜距離分析是評價遺傳多樣性的簡便方法,陳玉清[14]對四川小麥種質資源采用數量性狀及隨機擴增多態性DNA標記對不同時期育成品種的遺傳多樣性變化的分析結果與用親緣系數分析的結果一致,也驗證了當有完整系譜信息時,系譜距離分析是探討遺傳多樣性有效而簡便的方法。1949年以來,我國小麥品種經歷了4~5次大的更換,尤其是以河南、山東、河北、四川為代表至今育成品種繁多。
本研究采用親緣系數對河南、河北、四川、甘肅2010-2011年參加全國區試的92個小麥品種進行遺傳多樣性分析,以期研究其對種質資源的利用情況,為今后培育優良新品種提供依據。四川、甘肅是小麥條銹病菌源的發源地,其育種目標是抗病性,河北、河南是我國小麥的主產區,研究四省小麥的遺傳多樣性,對抗源的合理布局具有重要意義。
1 材料與方法
1.1 材料
試驗材料選用2010-2011年度小麥參試品種中四川、甘肅、河北、河南四個省的92個小麥品種,追溯其系譜信息至三代以上。其品種名稱、系譜信息列于表1。
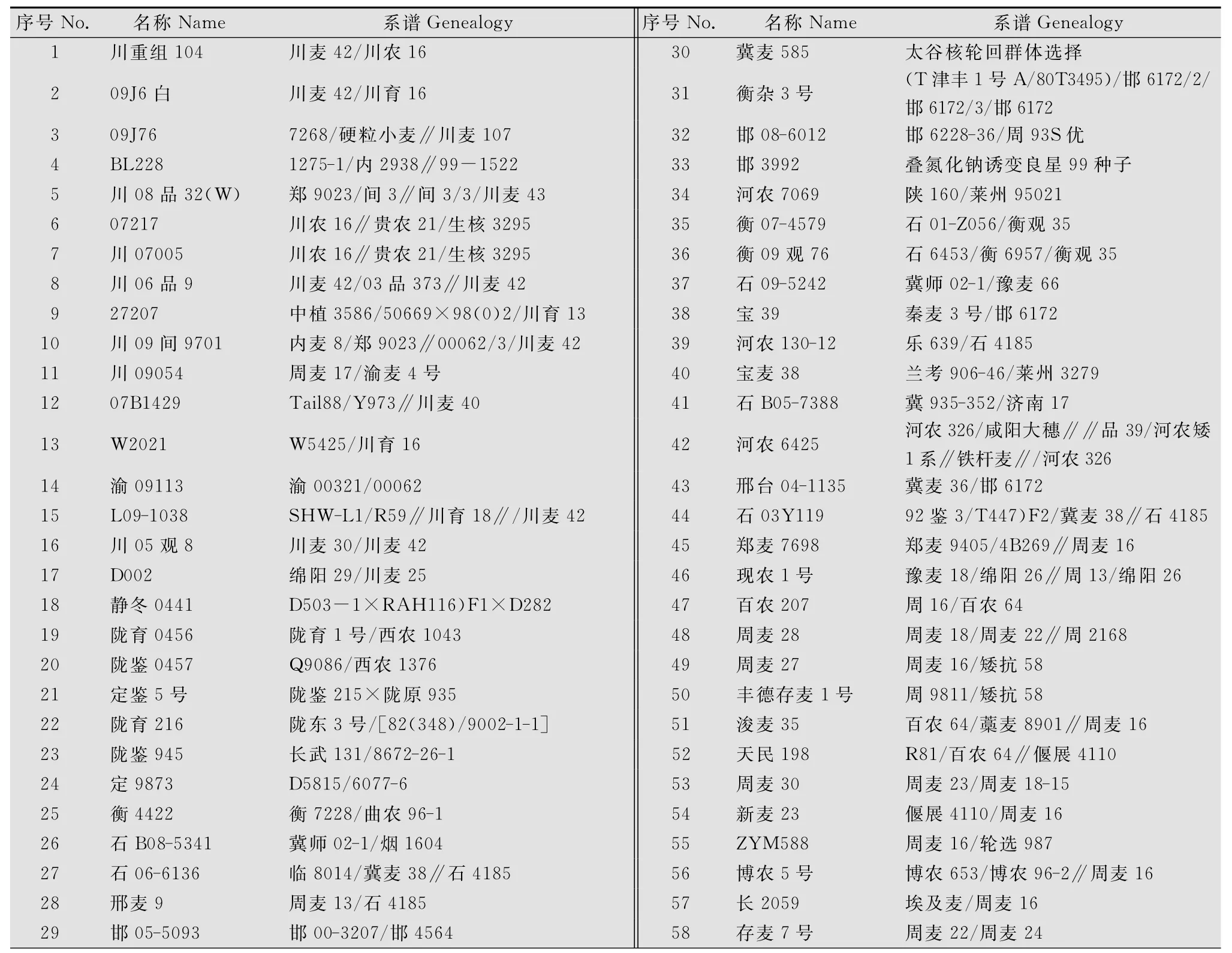
表1 供試品種的基本信息1)Table 1 Basic data of the cultivars tested

續表1 Table 1(Continued)
1.2 方法
1.2.1 親緣系數分析
計算四川、甘肅、河南、河北四省共92個品種成對組合的親緣系數,方法同Cox等[15],略有改變,計算原則如下:①一個品種分別從其雙親得到一半的基因;②所有祖先種、親本及其后代品種都是純合同型的;③沒有系譜信息的祖先種是不相關的;④系選品種、自然突變品種、誘變品種及核不育輪回群體選擇品種從親本中得到的基因是親本的3/4;⑤同一親本組合的后代兩品種間的COP=(0.75)2=0.562 5;⑥一個品種與其自身的COP=1。計算類內及類間的平均COP值。用(1-COP)矩陣表示系譜遺傳距離矩陣[16],采用SPSS16.0軟件的歐氏距離法對矩陣(92×92)親緣系數矩陣進行聚類分析。
1.2.2 抗條銹病鑒定
利用條銹菌混合優勢小種在田間分別對供鑒品種進行人工接種鑒定。供試的條銹混合圃鑒定菌系包括中國農業科學院植物保護研究所、甘肅省農業科學院植物保護研究所和四川省農業科學院植物保護研究所提供的CYR33、CYR32、CYR31、CYR30、CYR29、CYR17、Su-1和V26,接種后,待對照品種‘銘賢169’充分發病后,對區試品種進行田間調查,鑒定方法和調查記載標準《中華人民共和國農業行業標準(NY/T1443.1-2007)-小麥抗條銹病評價技術規范》。
2 結果與分析
2.1 供試品種組合間的親緣系數分析
四省參加全國區試的92份小麥品種,共組成4 186對組合,有618對組合間存在親緣關系,親緣系數變化范圍在0.0000~0.750 0之間,總COP值為95.793 91,平均值為0.022 9。表明四省參加全國區試的小麥品種部分存在遺傳相似性,但其遺傳程度不高,不管其是否通過區試試驗,均可以用這些品種組合培育優良品種。
從表 2 可 以 看 出,‘09J76’、‘07B1429’、‘渝09113’、‘靜冬0441’、‘隴鑒0457’、‘定鑒5號’、‘隴育216’、‘定9873’、‘衡4422’、‘邯05-5093’、‘邯08-6012’、‘邯3992’、‘河農7069’、‘石B05-7388’、‘河農6425’、‘鄭育麥518’、‘豫同69’、‘太空7號’、‘天寶98-7-5’與其余品種的COP均為0,其中‘靜冬0441’、‘隴鑒0457’、‘定鑒5號’、‘隴育216’、‘定9873’為甘肅省品種,說明甘肅在培育新品種時,能較廣泛地利用抗源。其余品種一方面是系譜信息追查不夠,另一方面是利用誘變育種的結果,另外也說明這些品種與其余供試品種的遺傳差異較大,利用這些品種培育優良品種的潛力很大。除去這19個品種,其余73個品種間的平均COP值為0.035 0,表明這19個品種與其余供試的大多數品種間無親緣關系,系譜距離上的遺傳差異很大,因此大大降低了總體水平上的遺傳相似性,增大了總體遺傳差異與遺傳多樣性。
‘金粒88’與其余品種組合間的COP總和為7.960 9,可見‘金粒88’與其余品種間的親緣關系密切,主要是其親本含有‘周麥13’與‘周麥16’,而‘周麥13’與‘周麥16’是姊妹系,均含有‘豫麥2號’的血緣,而河南省的48個小麥品種中含有‘豫麥2號’血緣的有31個小麥品種,也說明‘豫麥2號’仍是河南省小麥生產利用品種的主要骨干親本。‘周麥27’、‘百農207’與其余品種組合間的COP總和分別為6.825 0、6.164 1,同樣也是由于骨干親本中含有‘豫麥2號’的結果。

表2 單一品種與其余品種的COP1)Table 2 COP of mono-cultivars and the other cultivars
2.2 親緣系數聚類分析
用(1-COP)值表示系譜遺傳距離,采用SPSS16.0軟件的歐氏距離法對92×92親緣系數矩陣進行聚類分析,四川、甘肅、河北、河南四省參加區試的92個小麥品種可以明顯地分為A、B、C、D、E、F、G、H,共8類(圖1)。A類包括‘現農5號’、‘平麥108’等一類利用‘周麥9號’選育的品種;B類包括‘現農1號’、‘周麥30’、‘豐德存麥5號’等,這類品種主要是以‘豫麥2號’為主要親本選育而成,‘豫麥2號’是一個遺傳基礎廣泛的品種,從系譜上可以看出它含有意大利、荷蘭、朝鮮等國外品種的優良種質資源,并且具有豐產性好的地方品種的基因信息,具有適應性強、豐產性好、品質優良等特點[17],從聚類分析結果也可以看出河南省小麥育種中對‘豫麥2號’的重視以及對其衍生品種的成功利用;C類包括‘邢麥9’、‘周麥28’兩個品種,這兩個品種是親本中都含有‘周麥9號’;D類包括‘09J76’、‘W2021’、‘07B1429’、‘隴育0456’、‘豫同69’、‘鄭育麥518’等39個品種,這類品種遺傳基礎復雜,遺傳背景豐富,并且親緣關系較遠,育種中可適當加大對這些品種的使用力度;E類包括‘衡雜3號’、‘寶39’、‘邢臺04-1135’等以‘邯6172’、‘矮抗58’為親本選育的8個品種,‘邯6172’是2003年國家農作物品種審定委員會審定的越冬抗寒性好,耐后期高溫,熟相好。慢條銹病,中抗紋枯病,高感赤霉病,高感葉銹病和白粉病,對稈銹病免疫的小麥品種,屬半冬性中熟小麥品種,普遍表現高產穩產、抗凍耐旱、抗倒抗病;F類包括‘川重組104’、‘09J6白’、‘BL228’、‘川08品32(W)’、‘川06品9’、‘L09-1038’、‘川05觀8’,從系譜信息看這7個品種主要是以‘川麥42’、‘川麥43’這兩個品種同一親本組合的后代品種;G類包括‘07217’、‘川07005’,這兩個品種是同一親本組合的后代品種,遺傳相似性比較高;H類包括‘冀麥585’、‘漯6073’,這兩個品種都是通過核不育輪回選擇的,細胞核雄性不育可以用于發展雜種優勢利用和群體改良,是近些年來很受重視的研究領域之一。
2.3 不同省份參試品種遺傳多樣性分析
不同省份育成品種系譜遺傳相似性分析見圖2,從中可見,河南省參加區試品種間遺傳相似性最高,各品種組合間的平均COP為0.067 8,遺傳基礎相對較窄,遺傳多樣性相對最小;四川省、河北省參加區試品種次之,平均 COP分別為0.023 5、0.015 9,遺傳相似性較低,遺傳基礎相對較廣,表明這兩個省在近幾年的育種工作中廣泛利用優良種質資源,提高了育種材料的遺傳多樣性;甘肅省參加全國區試的品種遺傳相似性最小,平均COP為0.006 0,遺傳多樣性最高,說明甘肅省對種質資源的利用率較高。

圖1 品種間(1-COP)聚類樹狀圖Fig.1 Clustering diagram of different cultivars

表3 92個區試品種混合譜抗條銹性鑒定結果1)Table 3 Mixed-spectrum resistance of 92regional trial wheat cultivars to wheat stripe rust
甘肅、四川兩省的遺傳相似性較低,主要是由于甘肅、四川是小麥條銹病發病的菌源地,其品種選育尤其注重抗原的利用,否則一旦大面積種植品種對條銹病失去抗性,將造成全國小麥條銹病大流行,給小麥生產帶來不可估量的損失;河北省的遺傳相似性比較大,分析可能與系譜追溯不夠有關;河南省參加區試的小麥品種遺傳相似性較高,分析主要是河南省是小麥的主產區,小麥條銹病的發生比較輕,對小麥產量影響較小,育種家在育種時更多關注的是產量,所以主要選擇產量較高的‘豫麥2號’、‘周麥9號’作為親本進行選育,導致遺傳基礎相對狹窄。

圖2 不同省份參試品種遺傳多樣性比較Fig.2 Genetic diversity of the tested cultivars from different provinces
2.4 小麥抗條銹病鑒定結果
利用條銹菌混合優勢小種在田間對四省參加區試的92個小麥品種進行抗病性鑒定,結果見表3。甘肅省的品種除了‘隴育0456’外,‘靜冬0441’、‘隴鑒0457’、‘定鑒5號’、‘隴育216’、‘隴鑒945’、‘定9873’均表現高度抗病性(反應型為0型);四川省的區試品種‘09J6白’、‘川08品32’(W)、‘07B1429’、‘W2021’、‘渝09113’、‘D002’因被凍死而缺苗,沒有鑒定結果,品種‘09J76’、‘BL228’表現中度抗病反應;河北省的區試品種‘邯3992’、‘河農7069’、‘衡07-4579’、‘石03Y119’表現抗病反應(反應型為0型),‘石B08-5341’、‘邢麥9’、‘河農130-12’表現感病,嚴重度及普遍率都達到60以上,‘石06-6136’、‘邯05-5093’、‘冀麥585’、‘寶麥38’等品種均對條銹病表現中抗,反應型2~3,嚴重度及普遍率比較低;河南省的參試品種‘鄭麥7698’、‘現農1號’、‘鄭育麥9988’等表現出抗病反應(反應型為0型),其余品種‘陽光68’、‘零43’等品種對小麥條銹病表現中抗。
3 討論
隨著現代農業的迅速發展,農業集約化程度越來越高。為了滿足現代農業規模化、商業化的要求,農業生產中傳統的地方品種被遺傳改良的高產優質品種所替代,并且推廣應用的品種越來越單一,遺傳基礎越來越狹窄。在小麥育種和生產中,由于嚴重的遺傳侵蝕和長期單一使用骨干親本為育種關鍵材料,全球小麥品種的遺傳多樣性出現降低趨勢,一些關鍵的抗性基因幾乎來源于幾個甚至同一個親本[18]。因此研究小麥的種質資源,引入小麥優良基因和遺傳變異,對于豐富現代栽培小麥的遺傳多樣性和進一步開展小麥高產、抗病、優質育種具有重要意義。本研究結果表明,2010-2011年四川、甘肅、河北、河南四省參加區試的92個小麥品種可聚為8個類群,其中 A、B、C、E、F、G、H 類都為河南、河北等骨干親本及其衍生品種,D類為親緣關系較遠的品種。
陳玉清等[14]對四川小麥親緣系數聚類分析表明,40個供試品種聚為6個類群,其中2個引進品種單獨聚類,育自同一祖先種的大多數品種聚為一類,祖先中有‘繁6’、‘綿陽11’及其延伸品種的聚為一類,有‘阿勃’及其延伸品種的聚為一類,聚類結果很好地反映出育種資源的利用情況及遺傳多樣性程度。而本研究中,四川省的小麥品種大多聚為一類且抗病性基本相同,其原因為祖先品種中基本含有‘川麥42’。‘川麥42’是利用硬粒小麥——節節麥人工合成的高抗條銹病小麥新品種,張颙等[19]對‘川麥42’進行了抗條銹病遺傳分析,結果表明‘川麥42’對條銹菌CYR31由1對顯性基因控制,Li等[20]研究表明,‘川麥42’的抗條銹性受1對顯性基因控制(暫定名為YrCH42)。‘川麥42’現已被國內育種單位廣泛引用[21],可以作為高產、抗條銹的基因資源。
甘肅省的7個小麥品種都聚為D類群,這類品種遺傳基礎復雜,遺傳背景豐富,并且親緣關系較遠,且這7個品種基本都表現高度抗病性,這表現出甘肅省在小麥遺傳育種中充分考慮了對小麥條銹病的抗性,這與甘肅省是小麥條銹病的發源地有關,一旦源頭發病,并得不到有效控制,小麥條銹病將發生大流行。
從河南參加區試的小麥品種的系譜看,主要是利用前一個時期選育的品種作親本,特別是主要推廣品種作為親本的利用率較高,大多數小麥品種中含有‘豫麥2號’的血緣,究其原因‘豫麥2號’是很好的抗銹源,能抵抗銹病的發生,品質較好,與其他小麥品種雜交后,還表現出遺傳力高的特點。河南省小麥品種中聚為一類的大部分都是由‘周麥16’、‘周麥13’組成,‘周麥16’和‘周麥13’是由同一親本組合的兩后代品種,且中抗條銹病,所育成的品種對小麥條銹病均表現出中抗或感病,但是對河南省而言,小麥條銹病的輕度發生并不會造成嚴重的產量損失也不會影響全國小麥條銹病的大流行。
從不同省份間小麥品種的遺傳多樣性分析可以看出,河南、河北、四川的品種遺傳多樣性相對較小,其遺傳基礎相對狹窄,這與小麥條銹病抗病性的選育有關。
在目前開展的超級麥優質專用小麥育種工作中,應采用系譜分析田間農藝性狀生理性狀和分子標記等方法有目的地進行親本引入并篩選遺傳多樣性較高的新種質,同時要利用遠緣雜交、航天誘變、基因工程等高新技術創造新種質進一步加強對資源的創新研究,提高對親本選擇的預見性,改變小麥品種遺傳多樣性狹窄的局面,提高種質資源的利用率。
[1] 劉三才,鄭殿升,曹永生,等.中國小麥選育品種與地方品種的遺傳多樣性[J].中國農業科學,2000,33(4):20-24.
[2] 毛盛遠,劉來福,黃遠樟,等.冬小麥數量性狀遺傳差異及其在作物育種上的應用[J].遺傳,1979,1(5):26-30.
[3] Van Beuningen L T,Busch R H.Genetic diversity among North American spring wheat cultivars:Ⅲ cluster analysis based on quantitative morphological traits[J].Crop Science,1997,37:981-988.
[4] Metakovsky E V,Branlard G.Genetic diversity of French common wheat germplasm based on gliadin alleles[J].Theoretical and Applied Genetics,1998,96:209-218.
[5] Metakovshy E V,Kezevic D,Javomik B.Gliadin allele composition of Yugoslav winter wheat cultivars[J].Euphytica,1991,54:285-295.
[6] 孫其信,黃鐵誠,倪中福.小麥雜種優勢群研究I:利用RAPD標記研究小麥品種間遺傳差異[J].農業生物技術學報,1996,4(2):103-109.
[7] Siedler H,Messmer M M,Schachermayr G M,et al.Genetic diversity in European wheat and spelt breeding materials based on RFLP data[J].Theoretical and Applied Genetics,1994,88:994-1003.
[8] 耿惠敏,劉紅彥,宋玉立,等.40個河南省審定小麥品種遺傳多樣性的SSR標記分析[J].西北農業學報,2005,14(2):27-32.
[9] Mercado L A,Souza E,Kephart K D.Origin and diversity of North American hard spring wheats[J].Crop Science,1985,25:1058-1063.
[10] 徐曉丹,馮晶,藺瑞明,等.河南小麥主栽品種親緣系數分析[J].麥類作物學報,2011,31(4):653-659.
[11] 王江春,胡延吉,余松烈,等.建國以來山東省小麥品種及其親本的親緣系數分析[J].中國農業科學,2006,9(4):664-672.
[12] Souza E,Sorrells M E.Pedigree analysis of North American oat cultivars released from 1951-1985[J].Crop Science,1989,29:595-601.
[13] Mercado L A,Souza E,Kephart K D.Origin and diversity of North American hard spring wheats[J].Theoretical and Applied Genetics,1996,93:593-599.
[14] 鄭有良.四川主栽小麥品種遺傳多樣性研究[D].雅安:四川農業大學,2000.
[15] Cox T S,Lookhart G l,Walker D E,et al.Genetic relationship among hard red winter wheat cultivars as evaluated by pedigree analysis and gliadin polyacrylamide gel electrophoretic patterns[J].Crop Science,1985,25:1058-1063.
[16] Rebecca L B,Susan P L,Roy J M,et al.Predicting progeny variance from parental divergence in hard spring wheat[J].Crop Science,1998,38:243-248.
[17] 王翠玲,張燦軍,王書子,等.河南省優質小麥系譜追溯及遺傳改良分析[J].中國農學通報,2002,18(2):80-82.
[18] Porceddu E,Ceoloni C,Kiflandra D.Genetic resources and plant breeding:problems and prospects[M]∥ Miller T E.Koebner R M D,eds.Proc 7thInt Wheat Genet Symp.Cambridge,UK:Institute of Plant Science Research,1988:7-21.
[19] 張颙,楊武云,彭云良,等.小麥新品種川麥42抗條銹病遺傳分析[J].植物保護學報,2006,33(3):287-290.
[20] Li G Q,Li Z F,Yang W Y,et al.Molecular mapping of stripe rust resistance gene YrCH42in Chinese wheat cultivar Chuanmai 42and its allelism with Yr24and Yr26[J].Theoretical and Applied Genetics,2006,112:1434-1440.
[21] 張颙,楊武云,胡曉蓉,等.源于硬粒小麥——節節麥人工合成種的高產抗病小麥新品種川麥42主要農藝性狀分析[J].西南農業學報,2004,17(2):141-145.
