中華卵索線蟲核型分析
2013-09-11 07:23:10劉建拓張麗紅王國秀
植物保護 2013年2期
劉建拓, 張麗紅, 王國秀
(華中師范大學生命科學學院,遺傳調控與整合生物學湖北省重點實驗室,武漢 430079)
染色體數目、形態、大小等特征具有種的特異性,即每種生物都具備自己特有的核型(karyotype)[1]。核型分析(karyotype analysis)是對整套生物染色體的形態特征進行綜合分析,是生物分類的重要依據[2],對生物的遺傳變異、發育機制以及物種的起源進化等研究有著重要理論價值[3]。同時,染色體中的性染色體與性別決定有關[4],因此核型分析對確定不同的性別決定機制也具有重要意義。
中華 卵 索 線 蟲 (Ovomermis sinensis Chen et al.)是一種昆蟲病原索科線蟲,其寄生前期幼蟲能主動侵染斜紋夜蛾[Prodenia litura(Fabricius)]、黏蟲[Mythimna separata (Walker)]、棉鈴蟲[Helicoverpa armigera (Hübner)]等鱗翅目幼蟲,寄生率即等于害蟲死亡率,具有很大的生物防治潛力和潛在應用價值[5]。為充分利用這一寶貴資源,研究人員試圖通過體外培養來解決其大量繁殖問題,但由于培養過程中性別沒分化,生殖腺不發育、未能獲得成功[6]。因此,中華卵索線蟲性別決定機制成為目前研究的重點。已有的生物學研究結果表明,中華卵索線蟲性別分化是由線蟲的感染強度及其宿主體內營養物質的豐度所決定。感染強度越大,每條線蟲吸收的營養越少,越有可能發育為雄蟲,反之,則為雌蟲[7]。這種特殊的營養決定性別分化機制,在動物界極其少見[8]。目前,本課題組從線蟲生理生化及分子生物學水平對該機制進行了初步研究,取得了階段性成果[9-11],并對中華卵索線蟲營養決定性別分化的分子機理有了一定的認識。但迄今,尚未有人從細胞水平研究這一特殊性別決定機制。為此,本試驗從染色體組學方面,對中華卵索線蟲染色體數目及核型進行了研究,探究性染色體的有無,旨在進一步了解中華卵索線蟲的性別決定機制。
1 材料與方法
1.1 試驗材料
中華卵索線蟲:2001年河南上蔡縣麥田采得,本實驗室通過宿主棉鈴蟲在(28±1)℃恒溫培養箱傳代培養,挑取活力良好的雌、雄成蟲為試驗材料。
棉鈴蟲(宿主):購于中國科學院病毒所(武漢),(28±1)℃恒溫培養箱飼養。
1.2 試劑
(1)線蟲生理鹽水:0.70%NaCl溶液。2.8g NaCl溶于ddH2O(雙蒸水)至400mL,4℃冰箱保存。
(2)秋水仙素溶液:秋水仙素由Sigma公司提供。以0.7%生理鹽水配制成濃度為0.4mg/mL溶液,置于棕色瓶中,高壓滅菌,4℃冰箱保存。
(3)低滲溶液:0.075mol/L KCl溶液。0.28g KCl溶解于ddH2O至50mL,室溫保存。
(4)分散液:V(冰乙酸)∶V(50%乳酸)=3∶1。先用ddH2O和乳酸按體積比配制成50%乳酸溶液,之后按比例加入冰乙酸,配制成分散液,室溫保存。
(5)固定液、磷酸緩沖液、Giemsa染色液配制:方法見鄭喜邦等[12]。
A.1/15mol/L 磷 酸 鹽 緩 沖 液 (PBS,pH=6.8):無水 Na2HPO49.465g,KH2PO49.047g,分別配成1000mL溶液,然后取 KH2PO4溶液510mL和Na2HPO4溶液490mL,混合即可,也可按比例配制PBS溶液。
B.Giemsa原液:Giemsa粉由Sigma公司分裝。Giemsa粉0.5g,甘油33mL,將少量甘油先與Giemsa粉混合后,用研缽進行研磨,直至顆粒消失為止,然后再將剩余甘油全部倒入,56℃水浴鍋中保溫2h,最后加入33mL的甲醇,保存于棕色瓶內,置于陰暗處,待用拿出。
C.0.91%的Giemsa應用液:按Giemsa原液和磷酸鹽緩沖液(PBS,pH=6.8)1∶10比例配制染液,置于陰暗處。
1.3 試驗方法
(1)秋水仙素處理:挑取活力好的中華卵索線蟲雌、雄成蟲,置于濃度為0.4mg/mL秋水仙素溶液處理2h。
(2)生殖腺的提取:將線蟲置于潔凈玻片上,在蟲體前1/3處截斷,用刀背將生殖腺擠壓出來,解剖鏡下用昆蟲針仔細挑出生殖腺。
(3)低滲處理:將生殖腺迅速置于裝有0.075mol/L KCl溶液的1.5mL EP管中,低滲處理1h(28℃)。
(4)固定:1.34×104r/min離心3min,棄上清,用新配制的卡諾氏固定液[V(無水甲醇)∶V(冰乙酸)=3∶1],固定20min,1.34×104r/min離心2min,重復3次,每次固定加入新配制固定液。
(5)分散與終固定:用昆蟲針挑出生殖腺,置于預冷無菌的玻片上,迅速滴入少量分散液[V(冰乙酸)∶V(50%乳酸)=3∶1],使細胞分散開來,然后再加上述固定液固定。
(6)染色制片:加一滴 0.91%Giemsa染液(pH=6.8,磷酸鹽緩沖液(PBS)稀釋配制),室溫下染色20min,流水沖洗,自然干燥。
(7)觀察拍照分析:顯微鏡下觀察、拍照,選取染色體分散良好的中期分裂相進行染色體統計分析,計算染色體的相對長度及臂比值,參照Levan標準進行染色體分類,即臂比值為1.0~1.7、1.7~3.0、3.0~7.0及7.0~∞分別代表中著絲粒染色體、亞中著絲粒染色體、亞端著絲粒染色體以及端著絲粒染色體。
2 試驗結果
中華卵索線蟲單倍體數目為6(圖1),二倍體數目為12(圖2),染色體相對長度最長為9.25,最短為5.71,臂比值介于1.18與2.10之間,其核型公式為2n=2x=3sm+3m,其中1、3、4號為亞中著絲粒染色體,2、5、6號為中著絲粒染色體(表1),其核型模式圖如圖3。試驗結果證實雌、雄中華卵索線蟲染色體數目一致,未發現異形性染色體與隨體。

圖1 中華卵索線蟲單倍染色體(10×100)Fig.1 The haploid chromosomes of O.sinensis(10×100)

圖2 中華卵索線蟲中期分裂相與核型圖(10×100)Fig.2 The metaphase chromosomes and karyogram of O.sinensis(10×100)
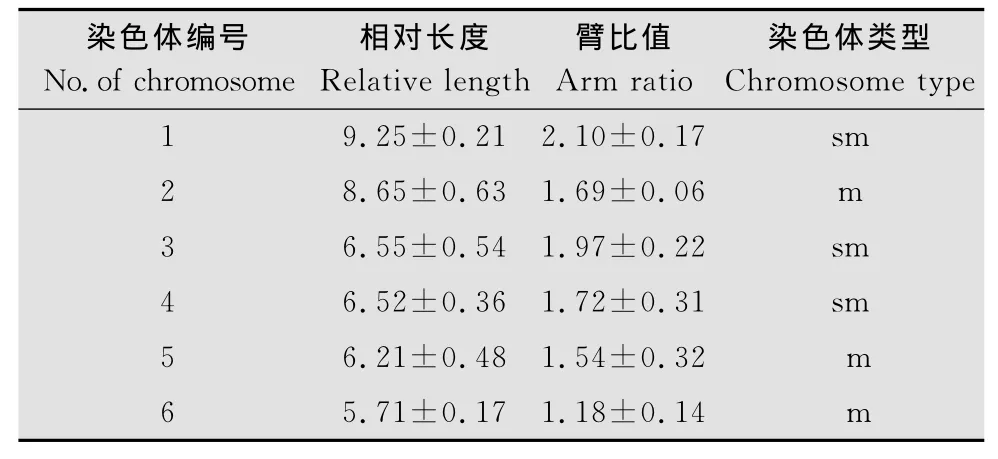
表1 中華卵索線蟲染色體相對長度及臂比值Table 1 Relative length and arm ratio of O.sinensis chromosomes

圖3 中華卵索線蟲核型模式圖Fig.3 The idiogram of O.sinensis karyotype
3 討論
3.1 染色體制備方法探討
核型分析最關鍵的是試驗材料,一般線蟲的核型分析是取材成蟲精巢和卵巢。但由于中華卵索線蟲的精巢或卵巢很難分離,所以本試驗以整條活力好的線蟲為試驗材料。本試驗進行了以下優化:(1)由于中華卵索線蟲體內充滿滋養體(脂肪粒),顯微鏡下無法看清其內部結構,使解剖提取其生殖腺的難度加大,也易導致精巢和卵巢丟失或活力下降。因此,本試驗采用先用秋水仙素處理整條線蟲,然后解剖并提取生殖腺;(2)處理過程中,秋水仙素濃度與處理時間同樣會影響到染色體制備。由于中華卵索線蟲體壁很厚,低濃度(0.02mg/mL)秋水仙素溶液很難滲透到蟲體內發揮作用,所以必須提高秋水仙素濃度,經試驗反復摸索,當濃度為0.4mg/mL,處理時間2h時,其效果達到最好。
3.2 核型分析與性別決定機制
國內關于昆蟲病原索科線蟲的研究,主要停留在形態分類、資源調查、生物學特性等[13]。近年開始涉及分子生化方面研究,但有關此類線蟲細胞遺傳學方面的研究,還從未有報道。中華卵索線蟲的核型分析,從細胞遺傳學角度為其營養決定性別分化機制提供了重要科學依據。
線蟲性別決定機制是復雜多樣的,但總體上主要分為兩類:基因決定型(genotypic sex determination,GSD)和環境決定型(environmental sex determination,ESD)。研究表明,昆蟲病原索科線蟲性別是由營養環境決定的[14],這種機制在動物界極其特殊。為深入了解這一機制,本試驗對中華卵索線蟲的染色體進行了研究,探究其有無性染色體。結果表明,中華卵索線蟲二倍體染色體數目為12,且雌、雄蟲染色體數目一致,這首先排除了XX/XO性別決定類型;通過對其核型分析發現,配對染色體之間形態特征差異不顯著,未發現異形性染色體(見圖2),不存在典型的XX/XY性別決定類型。以上結果表明,中華卵索線蟲性別決定應該為環境決定型,這也與前人的研究結果相一致[14]。盡管如此,染色體形態學結果也不能完全排除中華卵索線蟲存在性染色體的可能,由于物種進化比較低級,性染色體分化程度低,雖然性染色體有一定的分化,但在形態上分辨不出來,這種情況在魚類就有發現,有研究表明,大部分魚類核型無異形性染色體,但通過進一步研究,發現一些魚類存在性染色體,如通過C-帶技術,科學家證明了美洲齒鯉(Poecilia sphenops)和Pseudatocindus tetensis的性別決定類型分別為ZZ/ZW 和 XX/XY[4]。
綜上所述,本試驗首次從細胞水平對中華卵索線蟲染色體進行了形態學研究,對其染色體數目、類型進行了分析,未發現異形性染色體,這一結果從細胞的角度佐證了該線蟲環境決定性別分化機制。
[1] 閆素麗,安玉麟,孫瑞芬,等.染色體核型分析及染色體顯微分離技術研究進展[J].生物技術通報,2008(4):71-74.
[2] 吳甘霖.核型分析在細胞分類學中的應用[J].生物學雜志,2006,23(1):39-42.
[3] 林明敏,朱香萍.石鰈染色體核型分析[J].青島農業大學學報(自然科學版),2009,26(2):128-130.
[4] 常重杰,杜啟艷.魚類的性別決定和性染色體[J].淡水漁業,2002,32(2):56-58.
[5] 岳華梅,汪江一,王洪濤,等.中華卵索線蟲的研究與應用[J].昆蟲知識,2006,43(6):762-766.
[6] 王國秀,陳曲侯.中華卵索線蟲的體外培養[J].動物學報,2001,47(2):235-239.
[7] Robin J S,Moeen A H,Randy G.Sex ratio and the infection process in entomopathogenic nematodes:are males the colonizing sex[J].Journal of Invertebrate Pathology,1998,72:288-295.
[8] 任爽,王偉娜,趙娜娜,等.中華卵索線蟲的性別分化分子機理研究進展[J].應用昆蟲學報,2011,48(3):716-719.
[9] 高原,王國秀,陳思禮.中華卵索線蟲雌雄成蟲可溶性蛋白雙向電泳分析[J].動物學報,2004,50(1):141-144.
[10]賀俊飛,王偉娜,周青春,等.中華卵索線蟲tra-1基因cDNA片段的克隆及實時定量表達分析[J].植物保護學報,2009,36(4):324-328.
[11]任爽,陳沖,劉緒生,等.中華卵索線蟲vasa基因的克隆及其表達模式分析[J].植物保護學報,2011,38(4):333-338.
[12]鄭喜邦,何寶祥.尼克紅雞染色體核型分析[J].中國家禽,2003,25(3):8-10.
[13]鐘玉林,王國秀.我國昆蟲寄生索科線蟲研究近況[J].中國生物防治,2001,17(1):35-39.
[14]Andre P S.Evolution of the control of sexual identity in nematodes[J].Seminars in Cell & Developmental Biology,2007,18(3):362-370.
