抗水稻條紋病毒安全表達載體的構建及遺傳轉化
2013-09-11 07:23:16李紅偉王錫鋒
植物保護 2013年6期
關鍵詞:水稻
雷 陽, 李 羽, 王 彪, 李紅偉, 李 莉, 王錫鋒
(中國農業科學院植物保護研究所,植物病蟲害生物學國家重點實驗室,北京 100193)
由水稻條紋病毒(Rice stripe virus,RSV)引起的水稻條紋葉枯病是東亞稻區,尤其是中國、日本、韓國等的重要病毒病害,造成了巨大的經濟損失[1]。曾經于2001、2003、2004、2006和2008年在江蘇、山東、安徽、河南和上海等省份的粳稻種植區大規模暴發,2004年僅江蘇省的發病面積就達157萬hm2,占全省水稻種植面積的79%。流行年份,一般田塊減產20%~30%,嚴重田塊達到80%以上,甚至顆粒無收[2]。采取傳統治蟲防病策略對水稻條紋葉枯病有一定的防效,但大量使用化學農藥既污染環境,也易使害蟲產生抗藥性,而且介體灰飛虱(Laodelphax striatellus Fallén)傳毒具有瞬時性和持久性,使得防病效果欠佳,因此防治病毒病最有效的途徑是通過增強品種自身的抗性,達到主動預防病毒病害的目的。
RNA干擾(RNA interference,RNAi)機制的發現為植物抗病毒基因工程提供了一條全新的思路。RNAi是一種重要的基因沉默現象,可以通過雙鏈RNA(dsRNA)特異性降解對應序列的 mRNA,從而特異性地抑制相應基因的表達[3]。RNAi是一種古老的且進化上高度保守的防御機制,但是通過長期的共進化,病毒已經能夠從RNAi中逃逸,甚至產生抑制子來與之抗衡,植物體內天然的RNAi系統不能起到完全或較高水平地抵御某種病毒侵染的作用。利用基因工程手段將病毒的某一序列設計成雙鏈發夾結構,導入植物體使之表達,以便誘發RNA沉默,就有可能強化植物體內天然的RNA沉默病毒基因的能力。2007年代玉華等[4]構建了RSV全長CP基因RNAi載體,遺傳轉化水稻,獲得了穩定遺傳、高抗或免疫的植株。2011年Shimizu等[5]將RSV編碼的7個基因都構建成RNAi載體,分別導入水稻,接種試驗表明:轉CP、SP基因的水稻植株對RSV免疫,轉pC1基因的水稻植株抗病性顯著提高,但轉pC2和pC4基因的水稻植株抗病性沒有增加。
在抗病基因工程中,外源基因整合到植物基因組中的頻率很低,一般從千分之幾到百萬分之幾不等[6]。為了便于篩選到低頻率的轉化事件,常將外源目的基因與易于識別的選擇標記基因串聯,再通過標記基因篩選低頻率的轉化事件。然而,使用選擇性標記基因對植物細胞的生長和分化有不利的影響;對環境存在潛在危害,標記基因可能轉移到土壤微生物和其他植物,引起其他生物產生抗性;在食品上有一定的安全風險和消費心理障礙[7]。因此培育無選擇標記基因的安全轉基因植物已成為植物基因工程發展的趨勢。
目前獲得無選擇標記基因植物的方法主要有位點特異性重組、轉座子系統和共轉化法等。位點特異性重組系統能精確地引入外源基因,但需要進行二次轉化或雜交,費時、費力[8]。利用轉座子系統雖然能完整地刪除選擇標記基因,但需要通過有性繁殖分離目的基因和標記基因,周期長、效率低,且不穩定,難以定向優化[9]。共轉化法具有操作簡便,適用范圍廣,基因轉化效率較高等優點,被廣泛應用于水稻、煙草、油菜、大豆、玉米等植物中[10-12]。共轉化法就是把目的片段和選擇性擇標記基因分別構建于不同載體或同一載體的不同T-DNA區,期望借助于基因槍或農桿菌介導等轉化方法將選擇標記基因和目的基因同時導入同一受體細胞的不同染色體上。一方面利用選擇標記基因編碼的性狀,篩選出同時含有目的基因的細胞或再生植株。另一方面,通過轉基因植株后代自交,使目的基因和選擇標記基因發生分離,從而獲得不攜帶選擇標記基因的轉基因陽性植株。雖然已有利用共轉化法培育無選擇標記轉基因水稻的報道[13],但由于RNA干涉載體片段大,結構復雜,無抗性選擇標記基因植株的獲得率還不盡如人意。本研究應用RNAi原理,構建了RSV CP基因(RSV-HCP)安全表達載體,利用農桿菌介導的雙菌、雙質粒共轉化法,對轉化體系進行了優化,成功獲得了轉基因再生植株,經T1代自交分離、抗病性篩選,獲得了無選擇標記基因的抗RSV植株。
1 材料與方法
1.1 轉基因受體、病毒分離物及傳毒介體材料
水稻模式品種‘愛知旭’(‘Aichiasahi’)以及對RSV高感、優質高產的品種‘武育粳3號’(‘Wuyujing 3’)和‘淮稻5號’(‘Huaidao 5’)用作轉基因的受體材料。
RSV分離物(RSV-JS)采集自江蘇南京自然發病的水稻(Oryza sativa Linnaeus)植株,經 ELISA和PCR鑒定、純化后保存在-70℃備用。攜帶RSV-JS的灰飛虱由江蘇省農業科學院植物保護研究所提供,飼養在高感品種‘武育粳3號’上(26℃,14h光照)。
1.2 菌株和質粒載體
大腸桿菌[Escherichia coli (Migula)Castellani et Chalmers]JM110、根瘤農桿菌 [Agrobacterium tumefaciens(Smith Townsend)Conn]EHA105和RNAi載體pMCG161均為本實驗室保存。載體pCAMBIA1301為澳大利亞CAMBIA中心產品,本實驗室保存。
1.3 載體構建
1.3.1 目的基因的克隆
按照已公布 RSV CP基因序列(GenBank.DQ299166.1)設計引物。上游引物 RSV-HCP-F:5′-AGACTAGT GGCGCGCCGACTATGTGCATCACCATGAG-3′,引入限制性酶切位點SpeⅠ和AscⅠ;下 游 引 物 RSV-HCP-R:5′-AGGAGCTCCCTAGGACAGCCATCTTAACACCAG-3′,引 入酶切位點SacI和Av rⅡ,預期擴增片段約256bp。
采用Trizol法(Invitrogen)提取水稻葉組織的總RNA為模板,以RSV-HCP-R為引物合成第一鏈cDNA。再以合成的cDNA為模板,以RSVHCP-F和RSV-HCP-R為引物對進行PCR擴增,反應總體積50μL(模板100ng,dNTP 1mmol/L,聚合酶5U,RSV-HCP-F 0.5μmol/L,RSV-HCPR 0.5μmol/L),反應條件:94℃4min;94℃40s,52℃40s,72℃1min,30個循環;72℃10min。將擴增到的RSV-CP基因部分片段命名為RSVHCP。試驗以健康植株為陰性對照。將RSV-HCP克隆于pMD18-T載體,經驗證序列正確的重組質粒命名為pMD18-T-HCP(基因的測序工作由上海生物工程公司完成)。
1.3.2 無潮霉素抗性基因載體pCMBIA1301-hpt(-)的獲得
用限制性內切酶XhoI酶切載體pCMBIA1301,經電泳檢測、回收、純化大片段,再經T4DNA連接酶16℃連接過夜,即可得到去除潮霉素標記基因的pCMBIA1301載體,經PCR和酶切驗證的載體命名為pCMBIA1301-hpt(-)。
1.3.3 RSV-HCP基因RNAi載體的構建
選擇含有來源于水稻Intron的RNAi載體pMCG161,通過酶切、T4連接酶作用,將RSV-HCP正向片段插入在載體的酶切位點Asc I、AvrⅡ間,將反向片段插入在載體的Spe I、Sac I間,獲得RNAi載體pMCG161+/-R。由于該載體含有氯霉素抗性基因,不適合根瘤農桿菌介導的遺傳轉化,因此將中間載體pMCG161+/-R上的發夾結構,用BamH I、HindⅢ雙酶切,回收6920bp的片段,在T4DNA連接酶作用下,與同樣經BamH I、HindⅢ酶切的載體pCMBIA1301-hpt(-)連接(16 ℃過夜),得到無潮霉素標記基因的RNAi載體,以antisense IP 5′(5′-TTCCTGGGCTAAAAGAATTGTTGATTTGGC-3′)和 antisense IP 3′(5′-CCGGTTCTGCCGCTTTTTTTAAAATTGGAT-3′)為 引物對,進行PCR鑒定,正確的質粒命名為pCMBIA1301+/-R。
利用電擊法將質粒pCMBIA1301+/-R和pCAMBIA1301分別轉化到農桿菌EHA105感受態細胞中,得到的陽性重組子分別命名為E13R和 E1301[10]。
1.4 不同水稻品種對共轉化率的影響
分別將水稻品種‘愛知旭’、‘武育粳3號’和‘淮稻5號’的成熟種子滅菌后置于愈傷誘導培養基上,28℃黑暗培養10~15d。剝下成熟胚盾片長出的愈傷組織,進行繼代培養,每個品種各取長勢良好的愈傷900個,分為3個重復。將E13R和E1301分別在YM培養基(酵母膏0.3%,胰蛋白胨0.5%,麥芽糖0.5%,pH=9.0;Kana 50μg/mL;利福平50μg/mL)平板上劃線,28℃黑暗培養2~3d,挑取單菌落,220r/min振蕩培養。當A600至0.8時,4000r/min離心收集菌體,用AAM+100μmol/L AS培養基重新懸浮沉淀至A600為0.4。將E13R和E1301的懸浮液按照體積比為1∶1混合,分別對3個水稻品種共轉化。水稻愈傷組織誘導培養、與農桿菌的共培養、抗性愈傷組織的篩選再生及所用培養基配方等參照易自立[11]的方法。對T0代再生植株進行PCR檢測,分別統計出抗性愈傷數、苗數和陽性植株數,并計算出愈率、再生率和共轉化率。計算公式如下:

1.5 不同菌液配比對共轉化率的影響
將E13R和E1301的懸浮液按照體積比3∶1、2∶1、1∶1和1∶2混合,以水稻品種‘愛知旭’為受體,分別進行共轉化。對T0代再生植株進行PCR檢測后,分別統計出抗性愈傷數、苗數和陽性植株數,并計算出愈率、再生率和共轉化率。
1.6 轉基因植株的PCR檢測
以改進的CTAB法[14]提取轉基因植株幼嫩葉組織總DNA,采用兩重PCR反應體系,同時擴增RSV-HCP和hptⅡ基因。反應總體積50μL(模板100ng,dNTP 1mmol/L,聚合酶5U,RSV-HCP-F 0.3 μmol/L,RSV-HCP-R 0.3 μmol/L,N-hpt-F 0.3μmol/L,N-hpt-R 0.3μmol/L),反 應 條 件 同1.3.1節,預期PCR產物RSV-HCP片段為265bp,hptⅡ基因片段為1042bp。反應產物經1%瓊脂糖凝膠電泳檢測。hptⅡ擴增引物對為N-hpt-F和N-hpt-R。引 物 N-hpt-F 序 列 為 5′-CCCATGGGATGAAAAAGCCTGAACTCACCGCGAC-3′,N-hpt-R為5′-CCCATGGGCTATTTCTTTGCCCTCGGACGAGTGC-3′。
1.7 轉基因植株的抗病性分析
對水稻條紋葉枯病的抗性鑒定試驗在本所防蟲溫室內進行(溫度26~30℃,光照14h),以‘鎮稻88’為抗病對照,‘武育粳3號’為感病對照,管理按常規方法進行。接種和調查方法參照張世賢等[15],水稻條紋葉枯病分級標準參照盧百關等[16]。
1.8 數據處理
以ANOVA/MANOVA統計方法分析數據,并進行多重比較。
2 結果與分析
2.1 RNAi載體的構建
2.1.1 RSV CP基因部分核苷酸片段的克隆
以水稻葉組織總RNA為模板,以RSV-HCP-F和RSV-HCP-R為引物,通過 RT-PCR 擴增 RSVHCP,1%瓊脂糖電泳分析結果表明:從RSV侵染的水稻樣品中擴增到約260bp的預期片段,陰性對照則沒有擴增到任何片段。將該片段克隆到pMD18-T載體,得到重組質粒pMD18-T-HCP,對其測序結果表明目的片段長度為256bp,與GenBank No.DQ299166.1序列的201~434bp序列完全一致,說明獲得了RSV CP基因的部分核苷酸片段RSVHCP。通過在NCBI上BLAST發現,與RSV-HCP核苷酸序列一致性達100%的分離物達42個,這也說明RSV-HCP序列是高度保守性,將有利于通過RNAi獲得抗RSV的轉基因植株。
2.1.2 中間載體的構建
構建RNAi載體需要一個兩側帶有多酶切位點的合適Intron序列,以便將RSV-HCP的正反義鏈插入在Intron的兩翼,形成發夾結構。本研究選擇了具有水稻Intron的載體pMCG161,將RSV-HCP的正義鏈插入在酶切位點Asc I和AvrⅡ間,將反義鏈插入在酶切位點Spe I和Sac I間。當片段正向、反向均連入載體后,此時的載體含有兩個AscⅠ酶切位點,它們分別位于正義片段和負義片段的兩端,因此采用AscⅠ對重組質粒進行單酶切時,陽性質粒得到兩個片段,其中1650bp的小片段為有效的 發 夾 結 構,包 括 正 向 片 段 (256bp)、intron(1138bp)和反向片段(256bp);而作為陰性對照的空質粒,只得到一條約12913bp的大片段條帶(圖1),說明RNAi載體構建成功,命名為pMCG161+/-R,該載體具有氯霉素抗性。

圖1 重組質粒pMCG161+/-R的酶切鑒定(Asc I)Fig.1 Restriction analysis of recombinant plasmid pMCG161+/-R with AscⅠ
2.1.3 適合農桿菌轉化的安全RNAi載體構建
為了獲得適合農桿菌遺傳轉化水稻的去潮霉素抗性基因的RNAi載體,采用限制性內切酶BamH I和HindⅢ將RNAi載體pMCG161+/-R上有效的發夾結構切下,并在T4連接酶的作用下,連入到去除潮霉素抗性基因的載體pCMBIA1301-hpt(-)上獲得了重組載體pCAMBIA1301+/-R,該載體具有卡那霉素抗性。以 antisense IP5′/antisense IP3′為引物,單克隆為模板進行菌落PCR鑒定,結果表明:部分待檢樣品和陽性對照(pMCG161+/-R)擴增出1121bp的特異條帶,與預期片段大小一致,而陰性對照(pCMBIA1301-hpt(-))未擴增出任何條帶。pCAMBIA1301+/-R的酶切鑒定結果與pMCG161+/-R相似,同樣得到了1650bp的條帶,證明無標記基因的RNA干涉載體構建成功。進一步測序結果也說明RSV-HCP發夾結構已連入載體pCMBIA1301-hpt(-)中,成功獲得了適合農桿菌轉化,且在T-DNA區內不含有抗生素標記基因的安全RNAi載體,命名為pCAMBIA1301+/-R(圖2)。

圖2 RNAi載體pCMBIA1301+/-R的T-DNA區示意圖Fig.2 The construct of pCMBIA1301+/-R T-DNA
2.2 不同水稻品種的共轉化率
采用重組農桿菌E13R和E1301(按照1∶1濃度配比),選擇水稻模式品種‘愛知旭’和對RSV高感、但優質高產的水稻品種‘武育粳3號’和‘淮稻5號’等進行了共轉化。在轉化過程中,3個品種的成熟胚最初產生愈傷組織的能力無明顯的差別,但在重組農桿菌侵染后,經潮霉素篩選,‘愛知旭’產生的抗性愈傷組織最多,出愈率最高(25.6%);其次是‘武育粳3號’,出愈率為23.3%;而‘淮稻5號’產生的抗性愈傷數最少,僅為7.9%。在隨后成苗過程中,‘愛知旭’成苗需要時間最短,約45d左右,成苗數也最多,再生率為38.3%;而‘武育粳3號’的成苗時間稍晚,需要50d左右,同時成苗數也略少(35.7%)。3個品種中‘淮稻5號’的成苗時間最晚,80d后才出現少量再生苗,同時成苗數也最少,僅5.6%的再生率。對再生植株進行PCR檢測后,對陽性植株數進行統計,計算共轉化率,采用ANOVA/MANOVA統計分析數據,并進行多重比較,3個品種的共轉化率有顯著差異。可以看出水稻品種‘愛知旭’的共轉化率最高,達9.07%;推廣品種‘武育粳3號’的共轉化率略低于‘愛知旭’,達到6.59%;而‘淮稻5號’的所有再生植株均未檢測到外源目的基因(表1)。由此可見,本研究所用的遺傳轉化體系最適合模式品種‘愛知旭’的轉化,其次是‘武育粳3號’,而不適合‘淮稻5號’的轉化。

表1 不同水稻品種的轉化效率1)Table 1 Co-transformation efficiencies of different rice varieties
2.3 不同菌液配比對共轉化率的影響
選用最適合RNAi載體pCAMBIA1301+/-R的共轉化受體品種‘愛知旭’,分別用菌液E13R與E1301按照3∶1、2∶1、1∶1、1∶2混合對350個愈傷進行了共培養,對抗性愈傷、再生植株數等的統計,發現當E1301菌液濃度較高時(E13R:E1301為1∶2),盡管抗性愈傷和再生苗數都相對較多,但hptⅡ和RSV-HCP的共轉化率較低(5.97%),而當 E13R菌液濃度較高時(2∶1和3∶1)hptⅡ和 RSV-HCP的共轉化率較高(13.56%~16.33%),最終獲得的轉基因再生陽性植株數目較多(表2)。

表2 不同農桿菌菌液濃度對轉化效率的影響Table 2 Effects of different density ratios on co-transformation efficiency
2.4 無潮霉素抗性基因植株的獲得
將獲得的33個轉基因再生植株,通過帶毒灰飛虱人工接種后,其中5株未表現癥狀(0級),9株表現輕微癥狀(Ⅰ級),5株表現中等癥狀(Ⅱ級),12株癥狀嚴重(Ⅲ級),而受體‘愛知旭’發病嚴重,全部為Ⅱ級或Ⅲ級,發病率為100%。PCR檢測結果表明:表現為0~Ⅰ級的轉基因再生植株均擴增到潮霉素抗性基因(hptⅡ)和RSV-HCP。
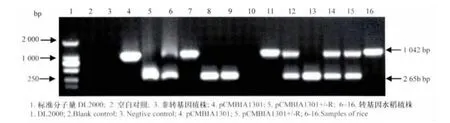
圖3 T1代植株的PCR檢測Fig.3 Analysis of T1transformation of hptⅡ gene and RSV-HCP fragment
對33個株系的T1代植株進行帶毒灰飛虱人工接種,篩選到14個株系的87株不表現病害癥狀的植株,通過雙重PCR檢測潮霉素抗性基因(hptⅡ)和RSV-HCP(圖3),其中16株(5個株系)為不攜帶hptⅡ的陽性植株。說明通過本研究構建的載體成功獲得了不攜帶選擇標記基因的轉基因抗病毒植株,為進一步育種應用打下了良好的基礎。
3 討論
在植物轉基因過程中,選擇標記基因的使用使得大量篩選抗性愈傷和再生植株成為可能。傳統的做法是將標記基因和目的基因串聯起來,置于同一載體(T-DNA)內,以便通過標記基因篩選低頻率的轉化事件[1,4,17],但后期刪除標記基因卻非常困難。隨著人們環保意識的加強,對標記基因的安全性提出了質疑,并引起了國際上的普遍關注,不同的國家也制定了相關的條款和法規加以限制。因此,在獲得轉基因植株后,是否能成功去除標記基因對轉基因材料的育種應用和安全釋放至關重要。在國內外,已有通過RNAi技術使水稻獲得抗RSV特性的報道。Park[1]利用 RSV 的全長 CP,Zhou[17]利用 RSV 的全長SP分別構建了RNAi載體,遺傳轉化水稻,均獲得了高抗或免疫的植株。Shimizu[5]更是將RSV編碼的7個基因的部分核苷酸序列(500bp)都構建成RNAi載體,分別導入水稻模式品種‘日本晴’,比較含不同基因的轉基因植株的抗病性差異,試驗表明轉化RSV CP和RSV SP的植株具有良好的抗性。但上述試驗更多的是在做機制層面上的研究,為了便于篩選,其轉基因植株均攜帶有選擇標記基因,難以直接投入生產或進行育種應用。另外,已有研究表明僅60~200bp的病毒基因片段就能誘導RNA沉默從而獲得抗病毒抗性[18],所以只要選擇具有高度保守性的RSV基因片段,就能達到全長基因的效果,這樣不僅大大簡化了構建植物表達載體的工作量,還縮小了T-DNA區的長度,更利于遺傳轉化和減小對水稻基因組的影響。
雙菌、雙質粒共轉化法作為一種簡便易行的去除選擇標記基因的轉基因方法,目前得到了廣泛的應用,而利用雙菌、雙質粒共轉化法轉化RNAi載體還未見報道。由于目的基因和選擇標記基因分別位于兩種質粒上,再加上RNA干涉載體的T-DNA區片段大,結構復雜,使其共轉化效率難以達到預期結果。本文利用RSV CP基因(969bp)的高度保守區段(RSV-HCP,234bp)構建了RNA干涉載體,與全長CP基因相比,既能保證對RSV CP基因的RNA干涉效果,還大大縮短了外源插入片段,使得轉化率(轉化率=再生苗數/共培養的愈傷數×100%)最高可達到19.14%,相比單質粒轉全長CP基因的轉化率(15%)[4]有明顯提高。
在雙菌雙質粒轉化中,通常的做法是將重組農桿菌菌液濃度按照1∶1配比,本試驗首先比較了在該配比下轉化水稻模式品種‘愛知旭’及生產上的推廣品種‘武育粳3號’和‘淮稻5號’的差異。結果表明,‘愛知旭’的共轉化率最高,‘武育粳3號’次之,而‘淮稻5號’沒有獲得任何陽性植株,不同品種間的差異較為顯著。這主要是由于在植物的遺傳轉化中,決定轉化成功與否的重要因素為材料的再生能力和效率,而這兩者最終是由品種的基因型決定的。此外,本研究結果表明在一定范圍內,含有目的基因的重組農桿菌濃度越高轉化效率越高,當E13R與E1301配比為3∶1時,共轉化率最高,濃度配比為2∶1時次之,濃度比為1∶2時最低,可能的原因是pCMBIA1301+/-R 的 T-DNA 區(11297bp)相對于pCMBIA1301的 T-DNA 區(5576bp)要 大 很多,這就造成了發卡結構相對于潮霉素抗性基因更難插入水稻的基因組中。本文通過雙菌雙質粒共轉化方法轉化水稻模式品種‘愛知旭’和生產上推廣品種‘武育粳3號’,均獲得了無標記基因抗RSV的轉基因植株,為轉基因材料進一步的推廣應用鋪平了道路。
[1] Park H M,Choi M S,Kwak D Y,et al.Suppression of NS3 and MP is important for the stable inheritance of RNAi-mediated rice stripe virus(RSV)resistance obtained by targeting the fully complementary RSV-CP gene[J].Molecules and Cells,2012,33(1):43-51.
[2] 陳潔,吳麗娟,周彤,等.江蘇省主栽水稻品種對條紋葉枯病與灰飛虱的抗性評價[J].南京農業大學學報,2010,33(4):105-108.
[3] Ding S W,Voinnet O.Antiviral immunity directed by small RNAs[J].Cell,2007,130(3):413-426.
[4] 代玉華,王錫鋒,李莉,等.一種適合水稻農桿菌轉化的RNAi載體的構建和潮霉素對水稻轉化的影響[J].植物保護,2007,33(2):37-40.
[5] Shimizu T,Nakazono-Nagaoka E,Uehara-Ichiki T,et al.Targeting specific genes for RNA interference is crucial to the development of strong resistance to rice stripe virus[J].Plant Biotechnology Journal,2011,9(4):503-512.
[6] De Vetten N,Wolters A M,Raemakers K,et al.A transformation method for obtaining marker-free plants of a cross-pollinating and vegetatively propagated crop[J].Nature Biotechnology,2003,21(4):439-442.
[7] Komamine Heksemsekya.Systems for the removal of a selection marker and their combination with a positive marker[J].Plant Cell Reports,2001,20(4):383-392.
[8] 邱淑萍,陳在杰,王鋒.Cre/loxp位點特異性重組系統在轉基因植物中的應用[J].福建農業學報,2008,77(2):211-217.
[9] 金維正,段瑞君,張帆,等.利用Ac/Ds轉座子系統在水稻中獲得無選擇標記轉基因植株的方法[J].生物工程學報,2003,19(6):668-673.
[10]Tenllado F,Diaz-Ruiz J R.Double-stranded RNA-mediated interference with plant virus infection[J].Journal of Virology,2001,75(24):12288-12297.
[11]Tenllado F,Llave C,Diaz-Ruiz J R.RNA interference as a new biotechnological tool for the control of virus diseases in plants[J].Virus Research,2004,102(1):85-96.
[12]Wang M B A D,Waterhouse P M.A single copy of a virus-derived transgene encoding hairpin RNA gives immunity to barley yellow dwarf virus[J].Molecular Plant Pathology,2000,1(6):347-356.
[13]劉峰,趙伊英,蘇永昌,等.雙農桿菌共轉化獲得無標記轉pepc基因的水稻植株(英文)[J].應用與環境生物學報,2005,11(4):393-398.
[14]Doyle J J,Doyle J L.A rapid DNA isolation procedure for small quantities of fresh leaf tissue[J].Phytochemical Bulletin,1987,19(4):11-15.
[15]張世賢,李莉,王錫鋒,等.我國北方稻區水稻條紋病毒分子變異和水稻品種抗病性分析[J].植物保護,2007,33(5):45-50.
[16]盧百關,方兆偉,劉漢清,等.不同粳稻品種抗水稻條紋葉枯病情況的調查研究[J].中國稻米,2006(1):22-24.
[17]Zhou Y,Yuan Y,Yuan F,et al.RNAi-directed down-regulation of RSV results in increased resistance in rice(Oryza sativa L.)[J].Biotechnology Letters,2012,34(5):965-972.
[18]Sijen T,Wellink J,Hiriart J B,et al.RNA-Mediated virus resistance:role of repeated transgenes and delineation of targeted regions[J].The Plant Cell,1996,8(4):2277-2294.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
