miRNA-17對靶基因整合素受體β8的調控表達作用
2013-09-05 05:07:00王福軍蔡軍劉巖顧松張希濤安向光顏均高杰蘇丕雄
山東醫藥 2013年13期
王福軍,蔡軍,劉巖,顧松,張希濤,安向光,顏均,高杰,蘇丕雄
(首都醫科大學附屬朝陽醫院,北京 100020)
miRNA是一類存在于真核生物中內源性非編碼小分子RNA,參與基因轉錄后水平的調控;它通過與靶基因序列的3'非翻譯區(3'UTR)以堿基互補配對的方式來執行對靶基因轉錄翻譯抑制功能,從而調控基因的表達[1]。miRNA-17~92家族與心血管疾病的發生密切相關[2],能促進肺臟細胞起源和心臟分隔作用的發生。整合素(ITG)是細胞表面的異二聚體受體,包含α和β兩種亞型,它們通過與細胞外基質配體結合,使細胞內效應器發生變化,最終引起病理生理變化。ITG表達與心血管疾病的發生、發展有著緊密關系[3],ITG的上調會導致心律失常和心肌收縮功能障礙[4]。有文獻指出,ITG受體β8(ITG-β8)和配體蛋白4.1B結合共同調控胚胎心臟的發育[5]。但是,有關ITG-β8與心血管疾病的發生機制尚不清楚。2011年3月~2012年12月,我們就miRNA-17與ITG-β8的調控抑制關系進行了研究。
1 材料與方法
1.1 材料 ①主要試劑:HEK293T細胞,Ultrapure Agarose(GIBCOTM美國);1 ×TAE:0.040 mol/L Tris-Acetate和0.001 mol/L EDTA,pH 7.5;DNA 電泳用6 ×loading buffer:40%Ficoll,0.06%溴酚蘭,培養液(GIBCOTMnvitrogen,美國),胎牛血清(中國醫學科學院血液病研究所);消化液:含有0.25%胰蛋白酶的0.02%的 EDTA,無血清培養基(GIBCOTMInvitrogen,美國),LipofectamineTM2000 Reagent(Invitrogen公司,美國),引物合成(華大基因,北京),RNAsin(Promega,美國);細胞裂解液:25 mmol/L Tris-HCl、pH 7.5,150 mmol/L NaCl,0.5%NP-40,0.5 mmol/L PMSF,0.5%protease inhibitor cocktail。②主要儀器:水浴恒溫箱(DK-8)(上海精宏實驗設備有限公司),水平電泳槽(北京市六一儀器廠),超凈工作臺(BCM-8)(蘇州凈化設備公司),電熱恒溫孵育箱(DNP-9082)、干燥消毒烤箱(DHG-9246A)(上海精宏實驗設備有限公司),紫外分光光度計(美國Bio-tek儀器公司),臺式離心機、低溫高速離心機(5417C)(德國Eppendorf公司),搖床(TS-1)(海門市其林貝爾儀器公司),CO2孵箱(Thermo Forma美國),熒光定量PCR儀(Bio-Rad iQ5美國),凝膠成像系統(UVP美國),F-4500 fluorescence spectrophotometer(HITACHI日本)。
1.2 方法
1.2.1 細胞培養 取HEK293T細胞置于含10%胎牛血清的RPMIDMEM培養液,在37℃、5%CO2孵箱中培養,2~3 d換液,待細胞融合70%~80%時,用0.25%胰酶消化傳代,取對數生長期細胞用于后續實驗。
1.2.2 靶基因載體的構建 應用生物信息學軟件Target Scan、Miranda和 PicTar對 miRNA-17進行靶基因預測。增強型綠色熒光蛋白cDNA用限制性內切酶消化并亞克隆成pcDNA3.1,用人腦的cDNA數據庫克隆成含有miRNA-17預測靶點的ITG-β8的3'UTR,即 pcDNA3.1/EGFP,進一步應用 BamHI、EcoRI設計成合適的核苷酸長度。
1.2.3 miRNA-17的真核表達載體的構建 生物學軟件預測靶基因序列和已知miRNA-17基因序列見圖1、圖2。目的片段經過T-A克隆后篩選正確的序列,將其亞克隆到pcDNA3.1(+)上,然后進行酶切鑒定和測序分析其正確性。將構建成功的重組質粒pcDNA3.1(+)-miRNA-17轉染HEK293T細胞 ,然后采用實時定量PCR法檢測成熟miRNA-17的表達水平。

圖1 生物學軟件預測靶基因序列
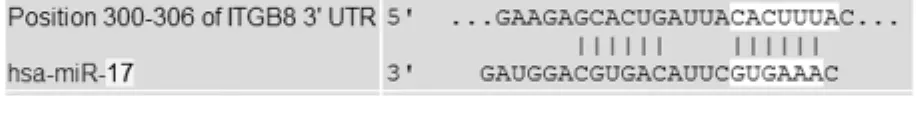
圖2 已知miRNA-17基因序列
1.2.4 細胞轉染與反義寡核苷酸技術 細胞常規培養于含10%胎牛血清培養基中,37℃、5%CO2,每2~3 d用0.25%胰酶消化傳代培養。將處于對數生長期的細胞消化,以2×105/mL接種至6孔板,繼續培養24 h待細胞長至培養板70%融合時轉染。轉染分為二組,即 ITG-β8+miRNA-17組和 ITG-β8+pcDNA3.1組。為了進一步證實miRNA-17調節抑制ITG-β8的特性,我們又設計合成miRNA-17的反義寡核苷酸序列(ITG-β8+ASO-miRNA-17)組和無義寡核苷酸序列(ITG-β8+ASO-Ctl)組,按照上述基因構建方法分別轉染HEK293T細胞。轉染后48 h,裂解細胞并提取蛋白。然后利用F-4500 fluorescence spectrophotometer檢測各組綠色熒光蛋白的光密度值作為綠色熒光蛋白的相對表達水平。
1.2.5 統計學方法 采用SPSS13.0統計學軟件,先對數據進行正態性檢驗,數據以表示,組間比較采用方差分析及t檢驗。P≤0.05為有統計學差異。
2 結果
綠色熒光蛋白表達水平:ITG-β8組為12.03±0.02,ITG-β8+miRNA-17 組為 9.33 ±0.04,ITG-β8+pcDNA3.1 組 為 11.04 ±0.09,ITG-β8+ASO-miR-17 組為 13.76 ±0.07,ITG-β8+ASO-Ctl組為11.34 ±0.09。ITG-β8+miRNA-17 組綠色熒光蛋白表達水平較ITG-β8+pcDNA3.1組明顯下降(P<0.05),ITG-β8+ASO-miR-17 組綠色熒光蛋白水平與 ITG-β8+pcDNA3.1 組、ITG-β8+ASO-Ctl組比較無統計學差異(P均>0.05)。見圖3。雙熒光素酶報告基因分析提示,miRNR-17能夠特異結合基因的ITG-β83'UTR區域使靶基因的mRNA表達量下降。HEK293T細胞未轉染、轉染miRNA-17、轉染ASO-miNA-17綠色熒光蛋白染色見圖4。
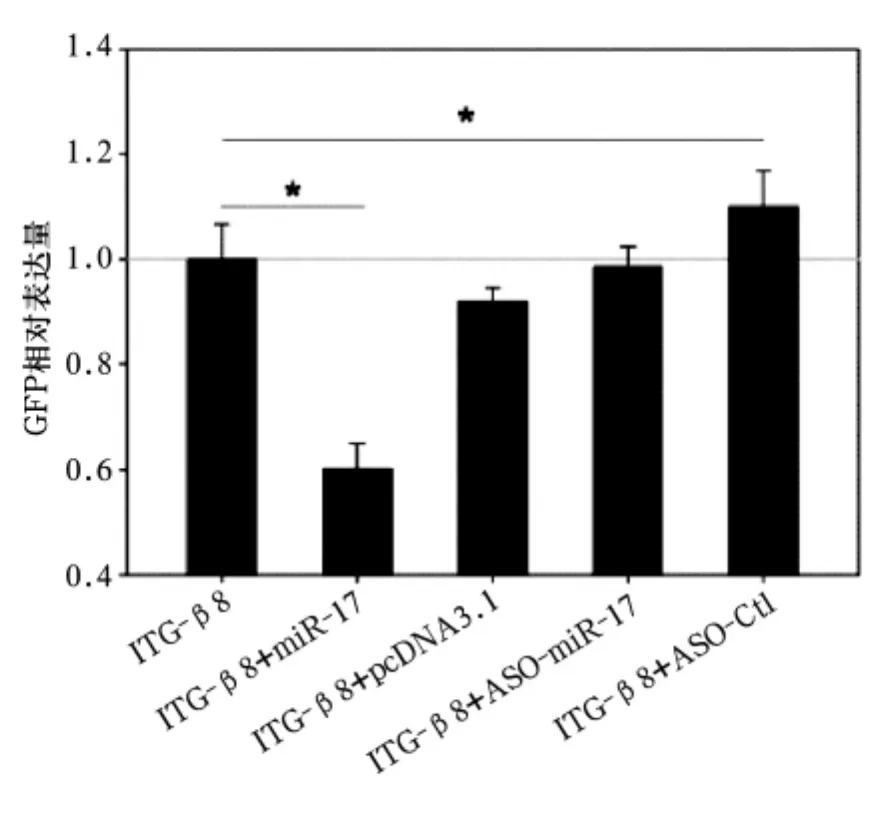
圖3 各組綠色熒光蛋白表達水平比較

圖4 HEK293T細胞轉染前后綠色熒光蛋白染色
3 討論
越來越多的研究發現,miRNA參與多種心血管疾病的發病過程,如心肌肥厚、心衰[6,7]、心律失常、心肌細胞凋亡等[8]。miRNA一般通過與靶基因miRNA 3'UTR的相互作用來調控靶基因的表達水平,尤其是miRNA5'端種子序列與靶基因的匹配程度尤為重要。我們通過網站預測到了miRNA-17可能的靶基因ITG-β8。BLAST數據庫比對表明,ITG-β8的3'UTR區確實存在與miRNA-17的5'端相互作用的序列。在熒光報告載體實驗中,我們發現miR-17能降低ITG-β83'UTR報告質粒的綠色熒光蛋白的表達水平,但對于ITG-β83'UTR報告空質粒pcDNA3.1-Egfp-ITG-β88-3'UTR 表達綠色熒光蛋白卻沒有明顯影響。說明miRNA-17能夠作用于ITG-β83'UTR的特異性序列,是miRNA-17的靶基因。利用反義寡核苷酸技術進一步證實,通過無義序列抑制miRNA-17的方法,可使綠色熒光蛋白恢復原來水平。
ITG是由α和β亞基通過非共價鍵組成的異二聚體,是細胞表面一類重要的兼具黏附和信號轉導功能的受體。α亞單位與配體結合發揮作用,而β亞單位主要介導黏附、連接、信號傳導等功能。近年來研究發現,β亞單位的胞質尾通過蛋白質之間的相互連接起到信號傳導的樞紐作用[9,10]。ITG是心肌細胞和成纖維細胞與細胞外基質之間相互作用的主要部分,在心臟發育過程中,ITG的模式表達變化表明細胞外基質與特定的ITG之間有協同表達的特點,不同亞單位組合形成的ITG在心肌發育和疾病等生理病理過程中發揮不同的作用。
本研究結果證明,ITG-β8是miR-17的靶基因,反義核酸技術進一步證實miRNA-17可以負性調控靶基因ITG-β8的表達。利用反義寡核苷酸技術開展針對miR-17的靶向治療,將可能減少先天性心臟病的發生。但是,ITG-β8影響心血管疾病的具體機制尚不清楚,還有待進一步研究闡明。
[1]Ambros V,Lee RC,Lavanway A,et al.MicroRNAs and other tiny endogenous RNAs in C.elegans[J].Curr Biol,2003,13(10):807-818.
[2]Wang J,Greene SB,Bonilla-Claudio M,et al.Bmp signaling regulates myocardial differentiation from cardiac progenitors through a MicroRNA-mediated mechanism[J].Dev Cell,2010,19(6):903-912.
[3]Sun M,Opavsky MA,Stewart DJ,et al.Temporal response and localization of integrins beta1 and beta3 in the heart after myocardial infarction:regulation by cytokines[J].Circulation,2003,107(7):1046-1052.
[4]Valencik ML,Zhang D,Punske B,et al.Integrin activation in the heart:a link between electrical and contractile dysfunction[J].Circ Res,2006,99(12):1403-1410.
[5]Jung Y,Kissil JL,McCarty JH.beta8 integrin and band 4.1B cooperatively regulate morphogenesis of the embryonic heart[J].Dev Dyn,2011,240(1):271-277.
[6]Care A,Catalucci D,Felicetti F,et al.MicroRNA-133 controls cardiac hypertrophy[J].Nat Med,2007,13(5):613-618.
[7]Thum T,Gross C,Fiedler J,et al.MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts[J].Nature,2008,456(7224):980-984.
[8]Cheng Y,Liu X,Zhang S,et al.MicroRNA-21 protects against the H(2)O(2)-induced injury on cardiac myocytes via its target gene PDCD4[J].J Mol Cell Cardiol,2009,47(1):5-14.
[9]Humphries JD,Byron A,Bass MD,et al.Proteomic analysis of integrin-associated complexes identifies RCC2 as a dual regulator of Rac1 and Arf6[J].Sci Signal,2009,2(87):ra51.
[10]Shattil SJ,Kim C,Ginsberg MH:The final steps of integrin activation:the end game[J].Nat Rev Mol Cell Biol,2010,11(4):288-300.
猜你喜歡
品牌研究(2022年29期)2022-10-15 08:01:00
品牌研究(2022年26期)2022-09-19 05:54:46
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
品牌研究(2021年35期)2022-01-18 08:45:18
火花(2019年12期)2019-12-26 01:00:28
海峽姐妹(2019年3期)2019-06-18 10:37:10
人大建設(2019年12期)2019-05-21 02:55:32
學苑創造·A版(2015年11期)2016-01-14 09:03:27
中國火炬(2010年8期)2010-07-25 11:34:30
百科知識(2008年8期)2008-05-15 09:53:30
