短花針茅葉片解剖結構及與氣候因子的關系
2013-04-10 06:13:30康薩如拉牛建明張慶陳麗萍
草業學報 2013年1期
康薩如拉,牛建明,2*,張慶,陳麗萍,3
(1.內蒙古大學生命科學學院,內蒙古 呼和浩特010021;2.中美生態、能源及可持續性科學研究中心,內蒙古 呼和浩特010021;3.南開大學生命科學學院,天津300071)
植物個體生長于自然環境中,自然受到干旱、鹽堿、低溫(冷害、凍害)、高溫等氣候因子的影響,然而植物對生存的不良環境具有特定的適應性和抵抗力[1-3]。植物對逆境適應與響應過程中有形態結構、解剖結構以及生理生化上的變化[4,5]。生長在不同生境中的植物,常表現出不同的形態結構;同種植物因生境的顯著差異,其形態和解剖結構也會表現出明顯的趨異現象[6,7],這與陳慶誠等[8]的多途徑適應觀點一致。植物形態結構能反映氣候因子對植物的影響以及植物自身對環境的適應,這通常被認為是植物對特殊環境的適應反應[9,10]。
植物葉片是反映植物對環境適應的最敏感器官,也是植物生命活動進行的主要部位[9]。因此在不同生境條件下,植物葉片的各個結構對其所處的環境有各自的響應特征[11]。暴露在空氣中的植物葉片大小、形狀及方向性在一定程度上決定著植物和環境之間能量相互轉換的效率[12-16]。Hegazy和Elamry[17]認為植物葉片形態差異會共同表現在植物葉片大小和形狀上,而恰恰這種特征保證了植物和環境之間物質和能量的相互轉換,是植物在干旱、半干旱環境中生存的重要條件。還有些研究指出,對于不同地段的植物群落,其葉片的類型、形狀、結構是所處環境最獨特的標志,并且認為物種的分化不僅表現在其葉片的外部形態結構,在葉片解剖結構及化學成分上也有極大的不同[18-20]。因此,研究植物葉片形態及解剖結構的分化特征,對植物適應性和抗性生理研究具有重要意義。
短花針茅(Stipabreviflora)系禾本科(Gramineae)針茅族(Stipeae)針茅屬(Stipa)須芒組(Sect.Barbatae)多年生叢生型旱生草本植物,是亞洲中部暖溫型荒漠草原的主要建群種,也常在某些典型草原群落及草原化荒漠群落中成為伴生成分[21]。短花針茅草原是亞洲中部草原亞區荒漠草原帶氣候偏暖地區的主要草原類型,它的分布從我國黃土丘陵區西北部起,向北越過陰山山地到內蒙古高原的南部地區,其南界大體上達到甘肅的蘭州、會寧、寧夏的固原、隴東的環縣,陜北的靖邊、榆林、綏德以及晉西北的河曲、偏關等地。短花針茅草原分布區的濕潤度在0.23~0.47,年降水量為279(化德)~434(準格爾)mm,年均溫為2.1(化德)~7.4℃(準格爾),蘭州可達9.1℃[22]。因此,短花針茅適于研究植物適應性及抗性生理[23,24]。本研究通過短花針茅葉片解剖結構對氣候因子的適應性研究,試圖闡明導致短花針茅葉片結構差異的主導環境因子,探究短花針茅適應環境的策略,為植物適應性和抗性生理研究及相關領域提供理論參考。
1 材料與方法
1.1 研究區概況
本研究設置野外調查樣地共8個,分布于內蒙古準格爾旗皇甫川、鄂托克旗三北羊場、鄂托克旗東深井、達茂旗巴音敖包蘇木、達茂旗百靈廟、達茂旗石寶鄉、蘇尼特右旗都仁烏力吉蘇木(圖1)。其中,在蘇尼特右旗都仁烏力吉蘇木選取2個不同生境下的樣地,其地理位置介于北緯39.24°~42.48°、東經107.34°~113.33°。樣地的植被類型包括溫性典型草原、溫性荒漠草原、暖溫性典型草原、暖溫性荒漠草原和草原化荒漠,詳情見表1[25]。
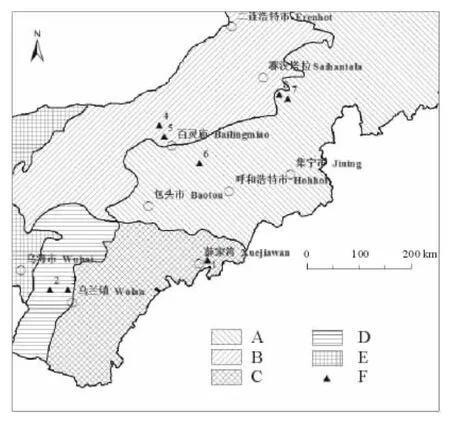
圖1 樣地分布Fig.1 The distribution of observation sites
1.2 研究方法
1.2.1 取樣 2007年8月,在研究區各樣地隨機剪取短花針茅(依據內蒙古植物志[22]及內蒙古大學植物標本館中短花針茅標本對所取樣品進行鑒定)植株相同高度的成熟、健康、向陽、無蟲害的基生葉3.0~3.5 cm,重復取樣25~30株,迅速放入福爾馬林-醋酸-酒精固定液(formalin-acetic acid-alcohol stationary liquid,FAA)中固定,帶回實驗室4℃保存備用。
1.2.2 制作石蠟切片及成像 將FAA固定液中固定的短花針茅葉片(每個樣地隨機選擇5~6株),通過脫水、透明、滲蠟、石蠟包埋、修蠟、切片、粘片(每片粘10~20橫切面,如圖2,每株重復3次)、烘片、脫蠟、雙重染色及封片等步驟,制作短花針茅葉片的石蠟切片[26,27]。
在OLYMPUS CX41顯微鏡下分別對切片進行4×10倍和10×10倍觀察,篩選出染色效果較佳的切片,使用CANON DS16275數碼相機以照片與切片100∶1的比例進行拍照與存儲,顯微照片示例圖像見圖3。并按JPEG格式存儲圖像文件。根據測微尺的刻度確定圖像的長和寬,分別為1 550.24和1 033.49 μm。
1.2.3 圖像分析及數據獲取 首先將JPEG圖像轉化為PIX圖像文件,在PCI Geomatica V 8.2軟件下,確定每張圖片左下角及右上角的坐標,定義單位為微米(μm)。
在 ArcMap version 9.3下每個樣地選取10~15幅染色及成像效果較佳的切片,對其實際結構進行矢量化。該處主要對葉片表皮、角質層、柵欄組織、維管束、主脈導管腔進行了矢量化與拓撲,形成Coverage文件[28]。

圖2 短花針茅葉片石蠟切片Fig.2 Paraffin section of S.breviflora
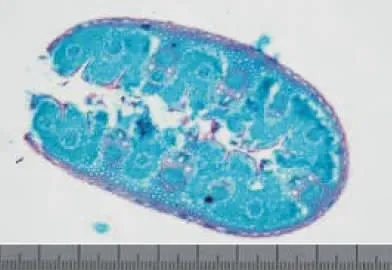
圖3 短花針茅顯微照片示例(1mm,100∶1)Fig.3 Micro-image of S.breviflora
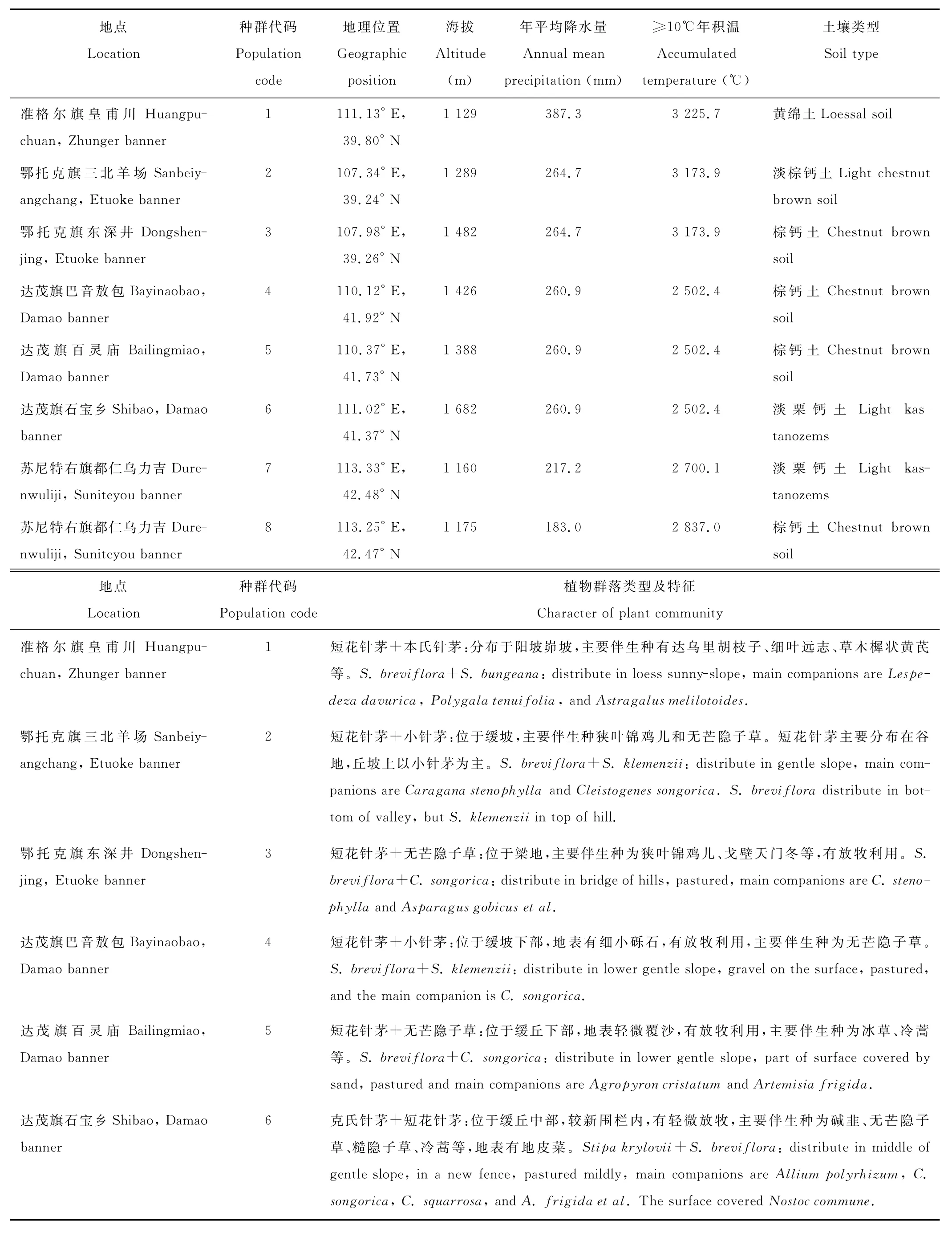
表1 樣地描述[25]Table 1 The description of observation sites

續表1 Continued
在ArcMap version 9.3下對葉片結構的長度、厚度、面積等參數進行測量統計后,確定以下6個指標,并獲得每個樣地指標的均值。
A:角質層占葉片厚度的比例(%)=角質層厚度/葉片厚度×100
B:維管束占葉面積比例(%)=維管束總面積/葉面積×100
C:柵欄組織占葉片厚度的比例(%)=柵欄組織厚度/葉片厚度×100
D:主導管占主維管束比例(%)=主維管束導管面積/主維管束面積×100
E:表皮細胞密度(%)=表皮細胞個數/統計長度×100
F:表皮毛密度(%)=表皮毛個數/統計長度×100
1.3 樣地與氣候指標的獲得
選取10個氣候因子,即≥0℃年積溫(AMT0)、≥10℃年積溫(AMT10)、最冷月平均氣溫(CMT)、最熱月平均氣溫(WMT)、年平均氣溫(MT)、年蒸發量(EVP)、年降水量(P)、夏秋降水量(P_SF)、冬春降水量(P_WS)及無霜期(FRZ_FD)。依據內蒙古氣候要素空間分布模型[29],基于調查地點的經度、緯度與海拔,分別獲取8個樣地的上述10個氣候因子指標。
1.4 統計分析
利用Canoco For Windows 4.5軟件,對葉片結構數據矩陣(6個指標)與氣候數據矩陣(10個指標)進行典范對應分析(canonical correspondence analysis,CCA),闡明葉片結構與氣候因子之間的關系,探究影響短花針茅分布及葉片結構分化的主導氣候因子。
結合上述CCA分析結果,選取2個主導氣候因子年平均氣溫和年降水量,利用SPSS 16.0,對葉片結構數據矩陣(6個指標)與氣候因子(2個指標)之間進行Pearson相關顯著性檢驗,闡明主導氣候因子與各個葉片結構的關系,探討短花針茅適應環境時所采取的策略。
結合8個樣地短花針茅的遺傳分化[25],對短花針茅葉片結構和遺傳結構數據矩陣之間進行Mantel檢驗,闡明遺傳分化在短花針茅葉片結構分化中所起的作用。
2 結果與分析
2.1 短花針茅葉片的結構特征
短花針茅葉片有以下典型的旱生特征(圖4)。如葉片橫切面完全卷曲成“V”型或橢圓型,表皮細胞排列整齊密集,上表皮有致密的表皮毛,有發達緊密的維管組織及維管束有發達的導管腔,葉片上表皮彎曲的每個凹陷處有數個運動細胞,厚壁組織排列緊密,具有加厚的角質層,有較大的氣孔密度等。
統計表明,短花針茅葉片厚度為(122.42±5.28)μm,角質層加厚,為(5.23±0.63)μm,表皮細胞密度為(4.90±0.36)個/μm,維管束總面積占葉面積的比例為(10.52±0.87)%,靠近下表皮的柵欄組織厚度為(38.16±3.29)μm,柵欄組織厚度占葉片厚度的比例為(31.44±1.64)%(表2)。
比較發現,不同樣地短花針茅主維管束導管腔占主維管束面積比值的差異最大,其次為表皮毛密度(表2)。而進一步對不同樣地間短花針茅葉片結構分化差異顯著性檢驗表明(表3),葉片結構分化具有極顯著性差異的有樣地8與樣地1,2,3,7之間和樣地2與樣地5,6之間。樣地8與樣地4,5和6之間的葉片結構分化差異顯著。
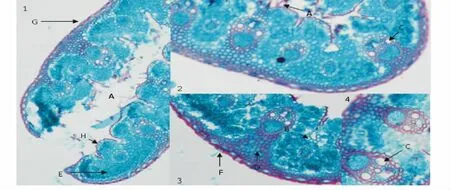
圖4 石蠟切片Fig.4 Parafin section
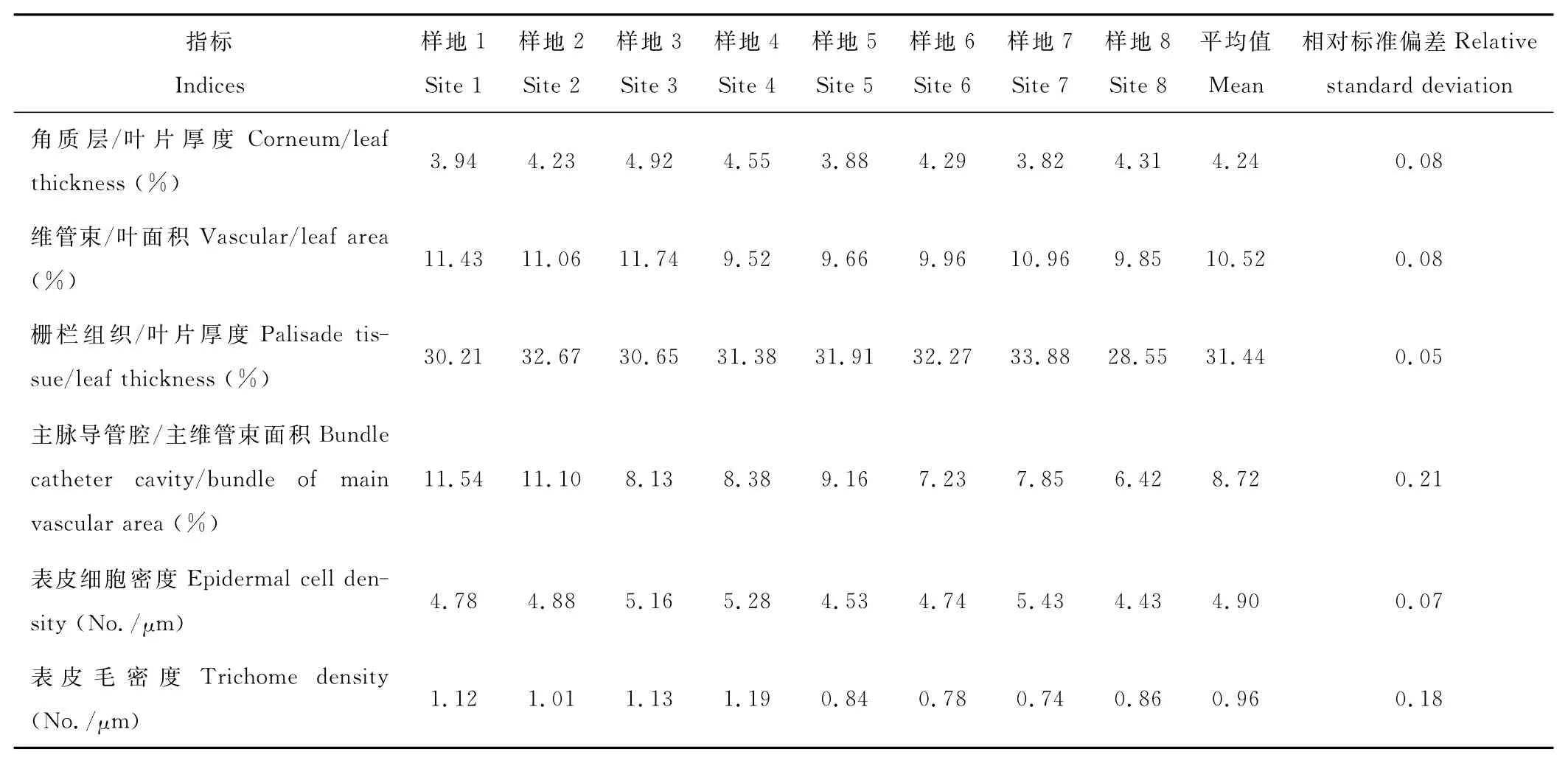
表2 6個指標統計數值表Table 2 Numerical value of six indices
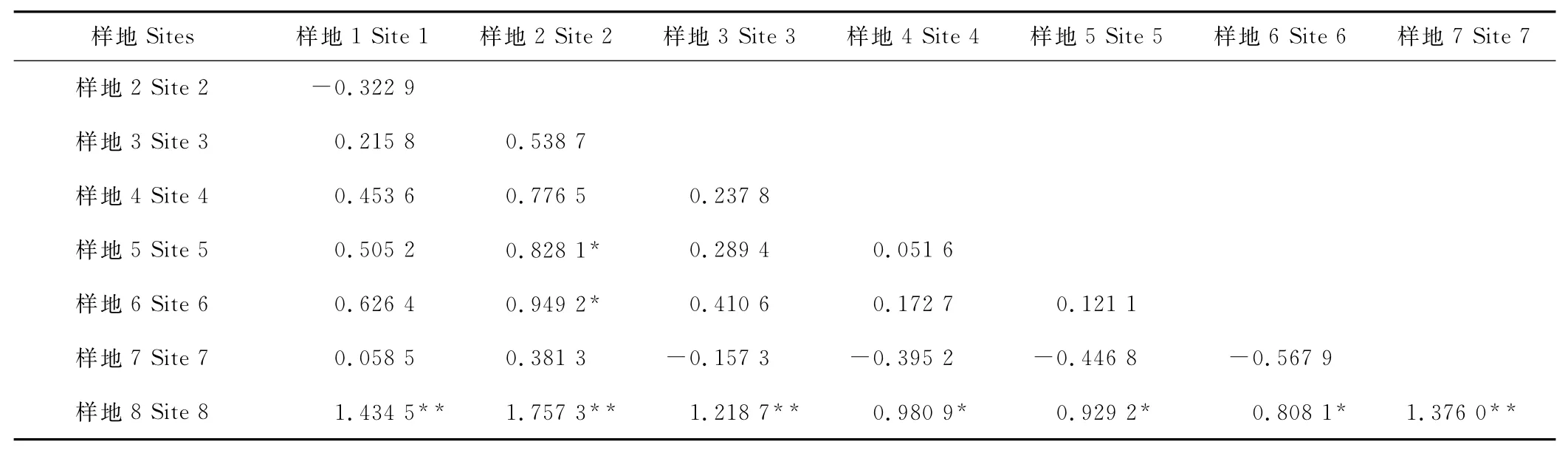
表3 樣地之間葉片結構差異顯著性檢驗Table 3 Variable significant analysis of eight sites
2.2 葉片結構與氣候因子的關系
第1主導因子為熱量因子(83.0%),而水分因子位居第2(13.9%)(圖5)。8個樣地在第1軸上受AMT0℃、AMT10℃、MT、FRZ_FD、CMT、WMT的影響較大,在第2軸上受P_SF、P、P_WS的影響較大。1,2和6,7,8樣地受熱量因子的影響較大;3和4,5樣地受水分因子的影響較大。1,2與熱量因子正相關,而6,7,8三個樣地與熱量因子負相關;第3樣地與水分因子正相關,4,5兩個樣地與水分因子負相關。
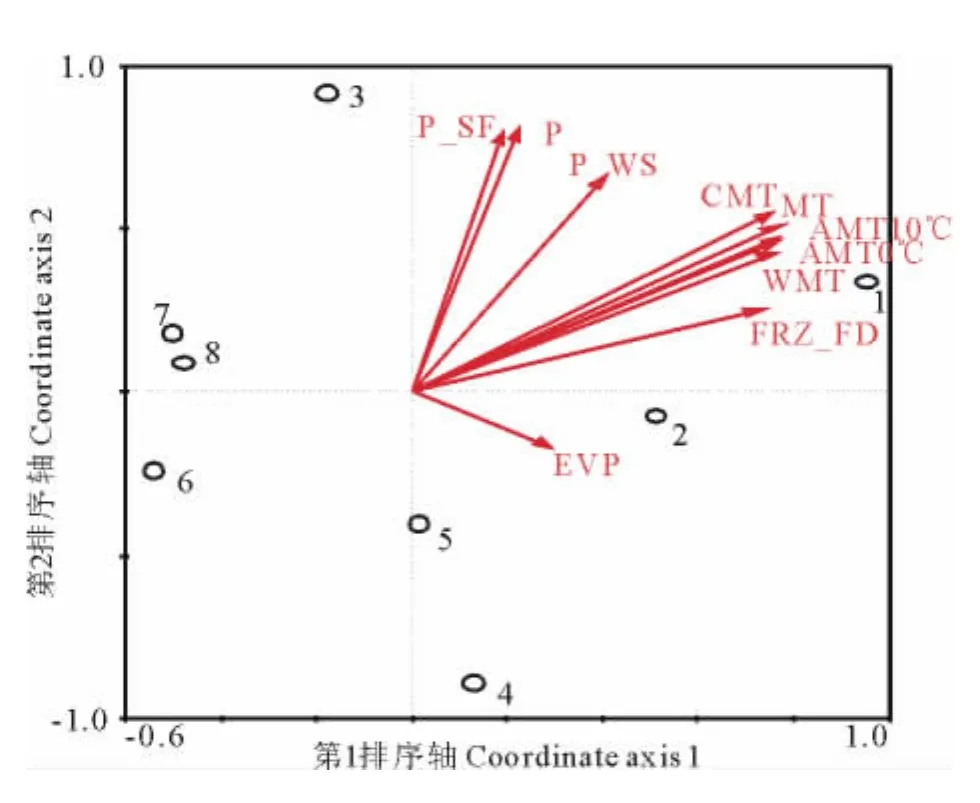
圖5 CCA分析結果Fig.5 The result of CCA
通過對信息量相關性矩陣的分析發現(表4),在熱量因子當中年平均氣溫對8個樣地短花針茅的影響最大(0.763),而≥0℃年積溫與≥10℃年積溫位居第2(0.762 8和0.759 1);結合排序結果,無霜期因子最接近第1軸,說明無霜期與熱量的相關性非常顯著。水分因子當中年平均降水量的影響最大(0.803 3),夏秋降水量居第2位;年降水量與夏秋降水量的相關性比與冬春降水量的相關性大,說明一年中夏秋降水量對8個樣地短花針茅生長過程中的影響比冬春降水量的影響大。
Pearson顯著性檢驗結果表明(表5),維管束占葉面積的比值和主維管束導管占主維管束面積的比值與熱量因子呈顯著正相關,而角質層厚度占葉片厚度的比值、柵欄組織厚度占葉片厚度的比值、表皮細胞密度、表皮毛密度與熱量因子的相關性不顯著。
與水分因子具有顯著相關性的指標為維管束占葉面積的比值,其他指標均不顯著。不過,盡管角質層厚度/葉片厚度、柵欄組織厚度/葉片厚度、表皮細胞密度與水分因子之間的相關性不顯著,但具有負相關特征,即隨著年降水量減小,角質層厚度增大,表皮細胞密度增加,柵欄組織厚度增大,符合植物抗旱性的特點。
2.3 葉片結構與遺傳特征的關系
對短花針茅葉片解剖結構與遺傳結構之間的Mantel檢驗結果表明(圖6),短花針茅葉片解剖結構與遺傳特征之間存在相似性,但不顯著。
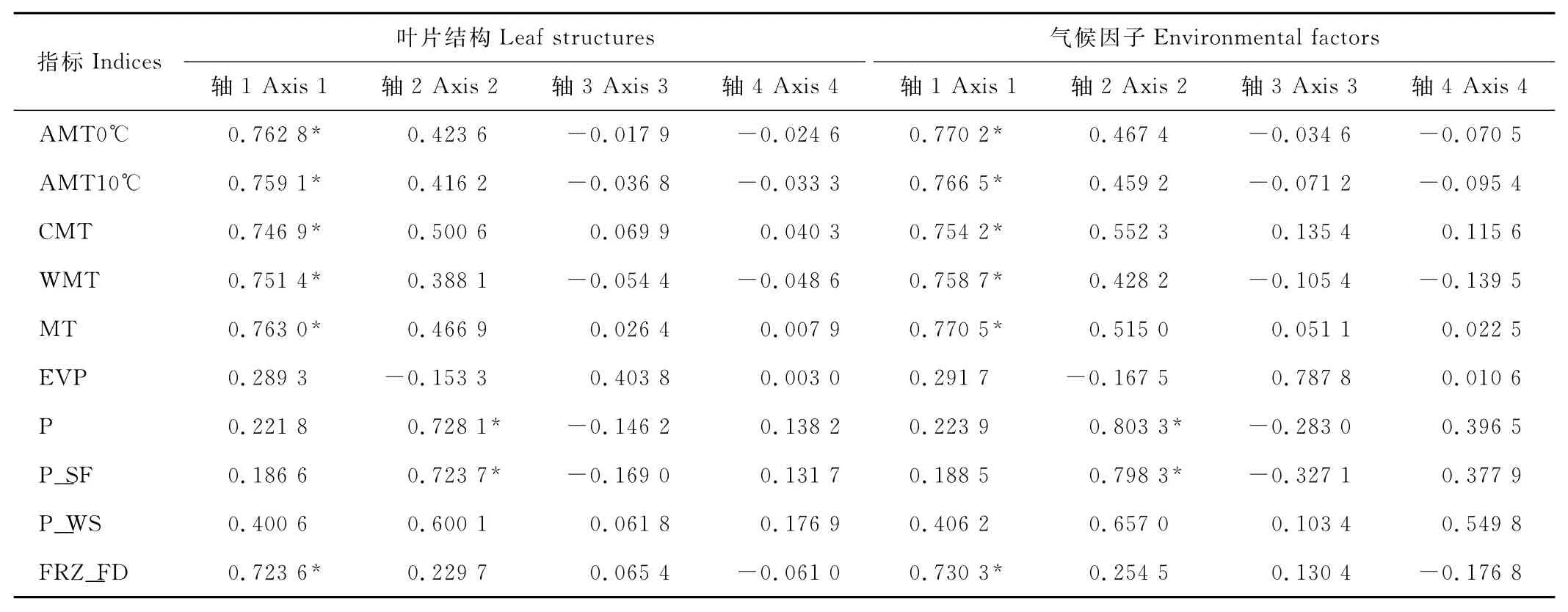
表4 CCA信息量相關性矩陣Table 4 Information table of CCA
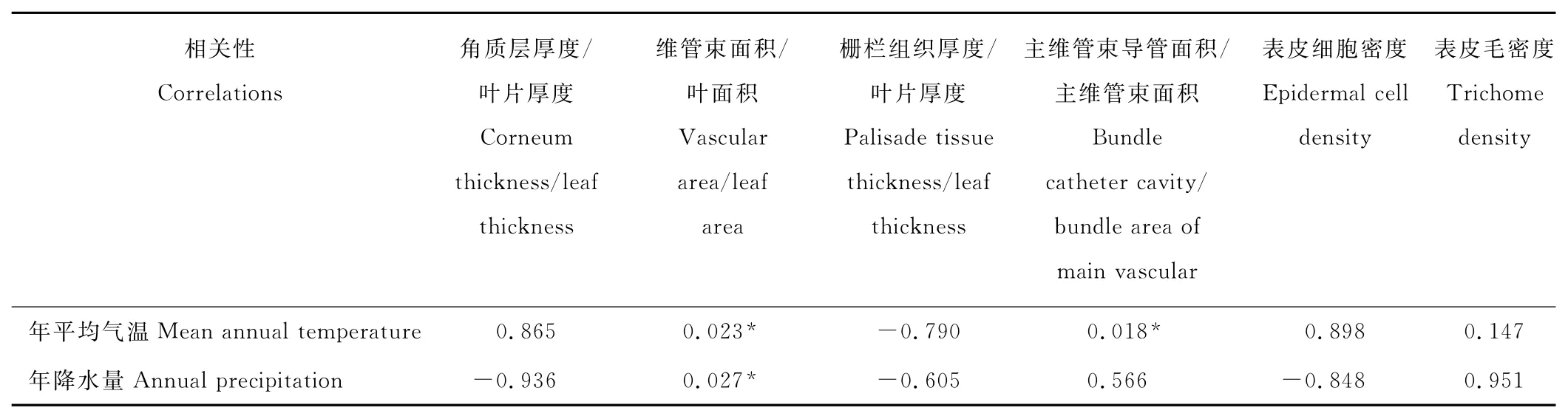
表5 Pearson相關顯著性檢驗結果Table 5 Result of Pearson test
3 結論與討論
3.1 短花針茅葉片具有典型的旱生結構
為了更好地適應高溫、缺水等干旱環境,植物葉片解剖結構均具有相應的變化特征[30-32]。不同干旱條件下的植物,其葉片本身、葉表皮附屬結構、機械組織、輸導組織及通氣組織等都較發達[33]。植物適應干旱環境的第一個特點是盡量縮小葉面積以減少蒸騰量,但不同植物適應干旱環境所采取的策略都有所不同[1]。有些植物隨環境干旱程度的加劇,不僅葉面積減小,有時退化形成鱗片狀甚至無葉[1,34,35],有些植物葉片較多肉質或薄葉植物的葉片卷曲[6,36,37];或葉片上下表皮角質層加厚[34,37-42],表皮較多白色絨毛,并且密度增加[6],葉表皮蠟質結構增多[35];或葉肉細胞較小,柵欄組織厚度增加,柵海組織比增大以及后壁組織發達[35];或葉脈致密,中脈厚度增加,葉脈維管束及導管腔發達[43];或氣孔下陷,單位面積氣孔數增多[33]等。而以上適應策略在同種植物中并不是單獨存在的,有些植物在極度干旱環境下同時采取以上少數幾種或多種適應策略[12]。黃振英等[36]把高溫、干旱環境下不同適應策略的植物歸納為薄葉植物、多漿植物、肉莖植物和卷葉植物4類,其葉片可歸為正常型、全柵型、環柵型、不規則型、禾草型、退化型6類。
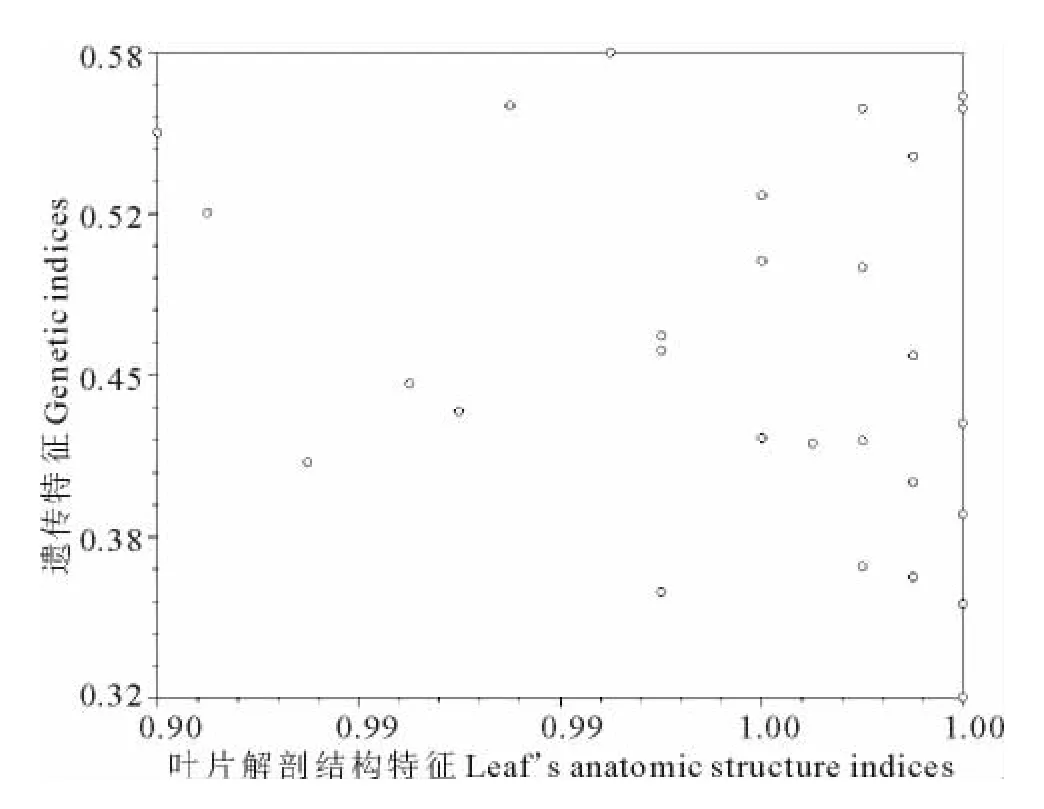
圖6 結構相似性系數與遺傳相似性系數之間的Mantal檢驗結果Fig.6 Result of Mantel test between structure indices and genetic indices
短花針茅葉片也具有典型的旱生特征,其適應策略為:1)葉片橫切面完全卷曲成“V”形或橢圓形,它屬于薄葉卷葉植物,以縮小暴露在空氣中的葉面積;2)表皮細胞排列整齊密集,減少蒸散量,并且上表皮有致密的表皮毛,可反射陽光,降低葉表面的溫度;3)有發達緊密的維管組織,增強了輸導水分和有機物的能力,而維管束導管腔也較發達,可增加對水分的輸導能力;4)葉片上表皮彎曲的每個凹陷處有數個運動細胞,可根據外部環境的變化調整葉片的卷曲程度;5)厚壁組織排列緊密,可減小水分蒸騰損失;6)角質層較厚,增加水分的擴散阻力,使葉片葉肉細胞更有效的利用水分,發達的角質層也可保護葉肉組織以免被強光灼傷;7)氣孔密度大,由于影響光合作用的一個主要因子為CO2的濃度[31],因此大的氣孔密度會增加CO2的吸收量,從而提高光合速率。
3.2 熱量是引起短花針茅葉片結構差異的主導氣候因子,水分居次要地位
在長期的進化過程中,每一種植物存在于一個適于生長和發育的地理區域內,形成了與氣候因素(例如溫度、降水、光照等)相適應的分布格局[44]。因此氣候是在大尺度上決定物種分布的主要因素[45]。通常認為,決定大尺度植被分布格局及植物形態結構差異的因素主要為溫度和降水等氣候因子[30,37]。而在小尺度上,影響植物形態結構差異主要由微環境導致,如地形條件(海拔)、土壤母質性質、土壤鹽分含量、土壤水分條件及其他生物因素[35,46]。植物的分布與氣候因子的關系,尤其是水熱因子的關系逐漸從定性描述轉變為定量描述[47,48]。本項關于短花針茅葉片解剖結構與氣候因子關系的研究與之有相同的結果。在較為廣闊的研究區域內(即大的空間尺度),熱量因子的分異對短花針茅葉片結構的變化產生了決定性的作用,水分因子處于次要地位,盡管其作用也是不能夠被忽視的。
既然熱量和水分因子是引起短花針茅葉片解剖結構分化的主要氣候因子,那么必定有一種或多種葉片結構與熱量和水分因子之間呈現相關關系。研究結果顯示,短花針茅葉片維管束/葉面積與熱量因子(年平均氣溫)和水分因子(年降水量)均顯著正相關,主脈導管腔/主維管束面積與熱量因子(年平均氣溫)顯著正相關,因此認為短花針茅極有可能在適應生長環境的水熱變化時,首先采取的策略是改變維管束結構,使得在高溫環境下能夠吸收更多的水分來調節葉片自身表面的溫度[44,49]。
樣地8雖然與樣地7的地理位置非常相近,但是由于樣地8的年平均降水量在8個樣地中處于最低水平。因此,樣地8的短花針茅葉片結構分化出能夠適應該地區極度干旱環境的獨特的葉片結構特征。
3.3 葉片解剖結構和分子結構之間有差異
每個生物都有其獨特的遺傳特性和形態特征,其形態特征是生物遺傳特性的外部表象,而遺傳多樣性決定了外部形態結構千姿百態的特征,因此形態結構的多樣化必然會受到遺傳分化的制約[50-53]。
通過與張慶等[25]關于短花針茅遺傳多樣性研究結果的比較發現,8個樣地短花針茅的葉片解剖結構與遺傳特征之間存在相似性,但不顯著。其原因極有可能是遺傳特征受到地理位置隔離的影響,受遺傳因子基因流的影響比較大[54],而屬于外部形態特征的葉片結構,不僅受到大尺度水熱條件的影響,還要受到微環境的影響;另一方面,形態特征是遺傳特征的外部表象,葉片只是植物器官的一部分,因此植物的遺傳特征不只表現在植物葉片上,而是綜合表現在根、莖、葉和其他繁殖器官上[55]。
[1] 周云龍.植物生物學[M].北京:高等教育出版社,1999.
[2] 李長復.葉子表面的適應性能[J].生物學通報,1985,(9):3-5.
[3] 李芳蘭,包維楷.植物葉片形態解剖結構對環境變化的響應與適應[J].植物學通報,2005,22(B08):118-127.
[4] 馮金朝,周宜君,周海燕,等.沙冬青對土壤水分變化的生理響應[J].中國沙漠,2001,21(3):9-12.
[5] 周宜君,劉春蘭,賈曉紅,等.沙冬青抗旱、抗寒機理的研究進展[J].中國沙漠,2001,21(3):98-102.
[6] 楊武,郭水良,方芳.不同生境下十七種蘚類植物葉的比較解剖學[J].云南植物研究,2007,29(5):409-417.
[7] Parida A K,Das A B,Mittra B.Effects of salt on growth,ion accumulation,photosynthesis and leaf anatomy of the mangrove,B.parviflora[J].Trees-Structure and Function,2004,18(2):167-174.
[8] 陳慶誠,趙松嶺,楊鳳翔.針茅草原放牧演替中種群消長的數學模型[J].植物學報(英文版),1981,23(4):323-328.
[9] 王勛陵,王靜.植物形態結構與環境[M].蘭州:蘭州大學出版社,1989:1-90.
[10] 周玲玲,宋曉麗.新疆四種補血草屬植物葉片的解剖學研究[J].廣西植物,2007,27(4):537-559.
[11] 蔣志榮.沙冬青抗旱機理的探討[J].中國沙漠,2000,20(1):72-75.
[12] Gates D M.Energy,plants,and ecology[J].Ecology,1965,46:1-13.
[13] Gates D M.Energy exchange between organisms and environment[J].Australian Journal of Science,1968,31:67-74.
[14] Gates D M.Transpiration and leaf temperature[J].Annual Review of Plant Physiology,1968,19(1):211-238.
[15] Parkhurst D F,Loucks O L.Optimal leaf size in relation to environment[J].The Journal of Ecology,1972,60:505-537.
[16] Smith W K.Temperatures of desert plants:another perspective on the adaptability of leaf size[J].Science,1978,201:614.
[17] Hegazy A,Elamry M.Leaf temperature of desert sand dune plants:perspectives on the adaptability of leaf morphology[J].African Journal of Ecology,1998,36(1):34-43.
[18] Cunningham S A,Summerhayes B,Westoby M.Evolutionary divergences in leaf structure and chemistry,comparing rainfall and soil nutrient gradients[J].Ecological Monographs,1999,69(4):569-588.
[19] Grubb P J.Plant populations and vegetation in relation to habitat,disturbance and competition:problem of generalization[J].Ecology,1985,63:921-931.
[20] Turner I.Sclerophylly:primarily protective[J].Functional Ecology,1994,8(6):669-675.
[21] 內蒙古植物志編輯委員會.內蒙古植物志(第二版,第五卷)[M].呼和浩特:內蒙古人民出版社,1994:200-202.
[22] 中國科學院內蒙古寧夏綜合考察隊.內蒙古植被[M].北京:科學出版社,1985.
[23] 盧生蓮,吳珍蘭.中國針茅屬植物的地理分布[J].植物分類學報,1996,34(3):242-253.
[24] 陳世鐄,李銀鵬,孟君,等.內蒙古幾種針茅特性和生態地理分布的研究[J].內蒙古農牧學院學報,1997,18(1):40-46.
[25] 張慶,牛建明,董建軍.內蒙古地區短花針茅(S.breviflora)種群遺傳多樣性[J].生態學報,2008,28(7):3447-3455.
[26] 李正理.植物制片技術[M].北京:科學出版社,1987.
[27] 李和平.植物顯微技術[M].北京:科學出版社,2009.
[28] 康薩如拉,牛建明,張慶.一種植物葉片的解剖結構顯微測量新方法——基于GIS技術[J].生物學雜志,2012,29(1):95-96,94.
[29] 牛建明.基于氣候的植被空間分布的數字模擬——以內蒙古為例[J].生態學報,2001,21(7):1064-1071.
[30] 胡建瑩,郭柯,董鳴.高寒草原優勢種葉片結構變化與生態因子的關系[J].植物生態學報,2008,32(2):370-378.
[31] 李翠,程明,唐宇丹,等.青藏高原2種柳屬植物葉片解剖結構和光合特征的比較[J].西北植物學報,2009,29(2):275-282.
[32] 孟林,楊宏新,毛培春,等.偃麥草屬植物種間苗期抗旱性評價[J].草業學報,2011,20(5):34-41.
[33] 強科斌,郭敏,張曉慶,等.圓囊苔草葉片的形態解剖學研究[J].草業學報,2007,16(2):76-83.
[34] 布仁圖雅.內蒙古典型草原退化演替過程中植物個體小型化解剖學機制研究[D].呼和浩特:內蒙古大學,2007.
[35] 鄧彥斌,姜彥成,劉健.新疆10種藜科植物葉片和同化枝的旱生和鹽生結構的研究[J].植物生態學報,1998,22(2):69-75.
[36] 黃振英,吳鴻,胡正海.30種新疆沙生植物的結構及其對沙漠環境的適應[J].植物生態學報,1997,21(6):34-43.
[37] 蔡永立,王希華,宋永昌.中國東部亞熱帶青岡種群葉片的生態解剖[J].生態學報,1999,19(6):844-849.
[38] He J S,Wang X L.Morphological and anatomical features of Quercus section suber and its adaptation to the ecological environment[J].Acta Phytoecologica Sinica,1994,18(3):219-227.
[39] 孫憲芝,鄭成淑,王秀峰.木本植物抗旱機理研究進展[J].西北植物學報,2007,27(3):629-634.
[40] 楊九艷,楊劫,楊明博,等.鄂爾多斯高原錦雞兒屬植物葉表皮特征及生態適應性[J].植物生態學報,2005,29(6):961-967.
[41] 史剛榮,趙金麗,馬成倉.淮北相山不同群落中3種禾草葉片的生態解剖[J].草業學報,2007,16(3):62-68.
[42] 萬宏偉,楊陽,白世勤,等.羊草草原群落6種植物葉片功能特性對氮素添加的響應[J].植物生態學報,2008,32(3):611-621.
[43] 李亞男,羅麗娟.不同居群狗牙根的葉片解剖結構比較研究[J].草業學報,2010,19(4):149-158.
[44] Cox C B,Moore P D.Biogeography:an Ecological and Evolutionary Aapproach[M].Oxford:Blackwell Publishing,2010.
[45] 余振,孫鵬森,劉世榮.中國東部南北樣帶主要植被類型物候期的變化[J].植物生態學報,2010,(3):316-329.
[46] 邰建輝,王彥榮,李曉霞,等.不同覆蓋物對無芒隱子草建植的影響[J].草業學報,2011,20(3):287-291.
[47] 柯文山,鐘章成,楊毅,等.四川大頭茶地理分布與環境因子的關系[J].生態學雜志,1999,18(6):24-27.
[48] 莫億偉,郭振飛,謝江輝.溫度脅迫對柱花草葉綠素熒光參數和光合速率的影響[J].草業學報,2011,20(1):96-101.
[49] 李洪山,張曉嵐,侯彩霞,等.梭梭適應干旱環境的多樣性研究[J].干旱區研究,1995,12(2):15-17.
[50] Helenurm K,Ganders F R.Adaptive radiation and genetic differentiation in Hawaiian Bidens[J].Evolution,1985,39:753-765.
[51] Gottlieb L,Warwick S,Ford V S.Morphological and electrophoretic divergence betweenLayiadiscoideaandL.glandulosa[J].Systematic Botany,1985,10:484-495.
[52] Charlesworth B,Lande R,Slatkin M.A neo-Darwinian commentary on macroevolution[J].Evolution,1982,36(3):474-498.
[53] Land R,Arnold S J.The measurement of selection on correlated characters[J].Evolution,1983,50:1210-1226.
[54] 王中生,安樹青,冷欣,等.島嶼植物舟山新木姜子居群遺傳多樣性的RAPD分析[J].生態學報,2004,24(3):414-422.
[55] 潘瑞熾,李娘輝.植物生理學[M].北京:高等教育出版社,2001.
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
