草原與荒漠一年生植物性狀對降水變化的響應
2013-04-10 06:13:30閆建成梁存柱付曉玥王煒王立新賈成朕
草業學報 2013年1期
閆建成,梁存柱,付曉玥,王煒,王立新,賈成朕
(1.內蒙古大學生命科學學院,內蒙古 呼和浩特010021;2.中美生態、能源及可持續性科學內蒙古研究中心,內蒙古 呼和浩特010021;3.內蒙古大學環境與資源學院,內蒙古 呼和浩特010021;4.煤炭科學研究總院唐山研究院,河北 唐山063012)
植物性狀(plant trait)或稱植物屬性(plant attribute),包括植物形態、生理或物候等性狀[1,2],是植物對外部環境適應的客觀表達[3]。由于能反映物種所在生態系統的功能特征而常常被稱為植物功能性狀(plant functional trait)[4,5]。植物性狀常沿資源和環境梯度發生變化[6],但因為資源的有限性,植物生長過程中植物性質無時無刻不在進行著權衡[7]。干旱環境條件下,水在限制和調節生命活動中占支配地位[8],植物若要完整的表達性狀并完成生活史,就必須權衡這些功能間的水資源分配。
位于干旱和半干旱地區的內蒙古草原與荒漠生態系統中有許多一年生植物在不同群落中組成恒有性層片(synusia),這些一年生植物層片主要由一年生小禾草、一年生豬毛菜(Salsolacollina)類、一年生蒿類等類群組成,不僅對水分脅迫和水分供應高度敏感,而且對水資源利用效率很高,即使一生中只有十幾毫米的降水,也能迅速完成生活史[9-11],因此常被稱作短命植物。
一年生植物層片這種對環境的高度適應性,一直備受生態學及其相關學科研究者的廣泛重視。前蘇聯著名植物生態學家拉甫連科[12]特別強調了一年生植物的類群及其生態作用,尤納托夫[13]在《蒙古人民共和國植被的基本特點》一書中也多次提到一年生植物層片。近年來國外關于一年生植物的研究也有報道,如Madon和Medail[14]報道了地中海草地一年生植物的生態意義和生存對策;Gonzalez-Astorga和 Nunez-Farfan[15]研究了個別一年生植物的萌發;Brown[16]報道了科威特荒漠群落中一年生植物的物種豐富度、多樣性和生物量;Krieger等[17]報道了象牙海岸島狀殘山一年生植物的季節動態;Robinson[18]研究了阿曼干旱林地荒漠一年生植物的生長和繁殖。我國干旱與半干旱地區草原與荒漠群落一年生層片的調查研究始于20世紀60年代初期,中國科學院內蒙古寧夏綜合考察隊在內蒙古及其毗鄰地區的植被與草場的綜合考察中,對這一層片進行了調查,在其出版的專著中首次提出了“夏雨型一年生植物層片”的概念,并對該層片的物種組成、類型及功能進行了概述[19,20]。近年來,隨著荒漠化的加劇,人們開始更加關注這一類群的植物。梁存柱等[9,10]、張德魁等[21]以及陶冶和張元明[22]對阿拉善地區、河西走廊地區以及古爾班通古特沙漠的一年生植物組成、分布及區系特征進行了研究,也有學者關注一年生植物種子萌發及種子庫的研究[23-27],還有一部分學者關注個別一年生植物的生理生態特征以及種群動態[28-31]。但上述研究均缺乏對該類群的水分生態適應性的系統研究,特別是水分控制實驗研究。生長于干旱半干旱地區的一年生植物類群,由于氣候的不穩定性和降水的波動性,在自然狀態下,如果沒有長期的定位研究,很難確切掌握一年生植物的生長與水分的相關性,而水分控制實驗研究,將是行之有效的方法,不僅可以準確了解一年生植物生長對水分需求,而且能更好地掌握不同水分梯度下一年生植物類群性狀間的權衡。
本研究通過室內降水控制實驗,人工模擬草原區至荒漠區逐漸減少的降水量,測量4種一年生植物在4個降水梯度下的物候與形態性狀特征。擬從植物對資源分配的角度探討一年生植物的生活史特征和不同水分梯度下一年生植物類群性狀間的權衡,探討一年生植物對水分的適應模式與適應策略。
1 材料與方法
1.1 研究材料與前處理
供試種子為2007年9月在內蒙古阿拉善左旗采集的豬毛菜和霧冰藜(Bassiadasyphylla)2種一年生豬毛菜類植物的種子,以及虎尾草(Chlorisvirgata)和冠芒草(Enneapogonbrachystachyum)2種一年生小禾草類植物的種子。于2008年3月30日將4種一年生植物種子播種于內徑20cm的塑料花盆中,并采用隨機區組設計將花盆擺放于內蒙古大學玻璃溫室培養。待植物長出第1對/片真葉時,間苗至每盆30株。
1.2 控制降水方法
由于一年生小禾草類比一年生豬毛菜類晚1~2d開始大量發芽[24],定于2008年4月12日、14日分別開始控制豬毛菜類、小禾草類降水。設置5,20,70和150mm的4個降水梯度,降水梯度的選擇依據荒漠區至草原區遞增的有效降水,150mm完全被植物利用的有效降水已達到草甸草原的有效降水量。降水量計算方法:在直徑為20cm的雨量器內,每降水31.4mm相當于自然降水1mm,即:3.14×(10cm)2/31.4cm3=S/A,式中,A代表降水量,S代表降水面積,即在面積為S的地面上降水AmL相當于1mm降水[32]。降水時間設在19:30,降水量及降水次數因梯度而異,具體設置見表1。
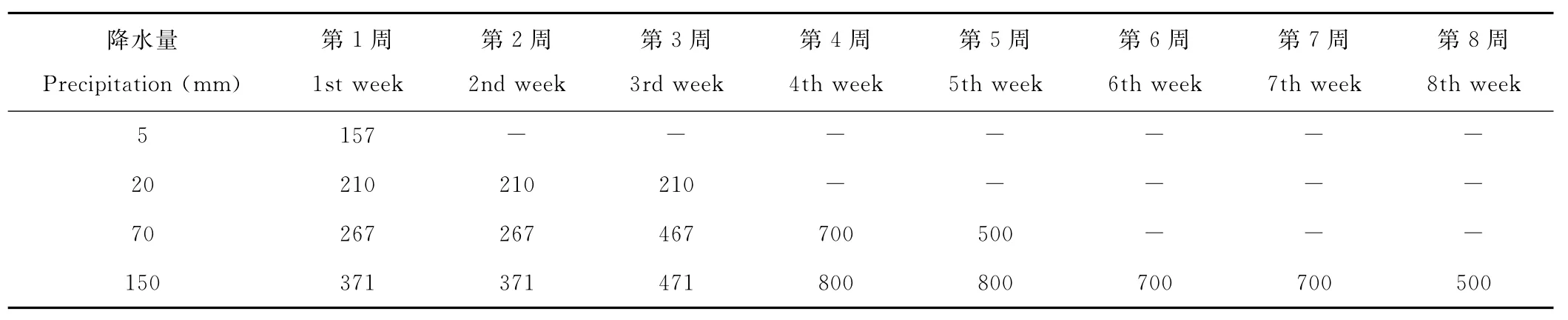
表1 不同降水梯度的降水時間及降水量Table 1 Irrigation times and amount in different rainfall grads mL/盆 Pot
1.3 物候期劃分方法
針對一年生植物物候期變化節奏較快并以種子越冬的特點,自種子開始萌發至幼苗出土記為萌動期,自開始展葉至展葉盛期記為展葉期,自花蕾或花絮開始出現經開花始期、開花盛期至開花末期記為開花期,自果實始熟經果實全熟期至果實脫落(種子散布)期記為果期[33]。
1.4 植物性狀測量方法
于2008年4月18日、20日分別開始測量一年生豬毛菜類和一年生小禾草類的形態學指標。在上述4個降水梯度處理的每種植物中隨機選3盆作為實驗重復進行測量,每盆隨機抽取15株為一個樣本。每7d用直尺測量1次植株自然高度(株高)、葉片長度(葉長),并稱量樣本地上部分鮮重(生物量)。每14d從4種降水條件下的4個物種中隨機選取3盆,采用將整盆植物地上部分全部剪下,同時將花盆中的根全部用水洗出,并分別包好、編號、標記,65℃烘干24h后稱干物質重量,并依“根冠比=地下部分干重/地上部分干重”計算根冠比[34]。于停水后植物約50%枯黃時統計結實量,其中一年生豬毛菜類依結實個數統計,一年生小禾草類依生殖枝數統計。
1.5 統計分析方法
用 Microsoft Excel 2010軟件完成全部數據處理,使用SigmaPlot 10.0軟件作圖,采用SPSS 16.0統計分析軟件進行獨立樣本T檢驗和單因素方差分析,并用LSD顯著性檢驗方法比較差異性。
夕照輕陰郭,回車訪棗花。 山門分野色,經閣帶林霞。 香篆穿簾細,旛風曳柳斜。 摩娑紅杏卷,觴詠想王查。[5]
2 結果與分析
2.1 一年生植物的物候期特征
在溫室培養條件下一年生植物的物候期經歷了萌動期、展葉期、開花期和果期4個階段。但一年生植物在不同降水條件下各物候期的持續時間各不相同。隨降水量增加,植物生長期或壽命逐漸延長,5mm最低降水量與150mm最高降水量相差約1倍。表現為展葉期、開花期、果期均增長,萌動期長短因種子萌發特性而異(表2)。在不同降水條件下,除虎尾草在5mm降水條件下未結實外,其余全部結實,表明在極少的降水條件下,這些一年生植物大部分能完成其全部生活史。
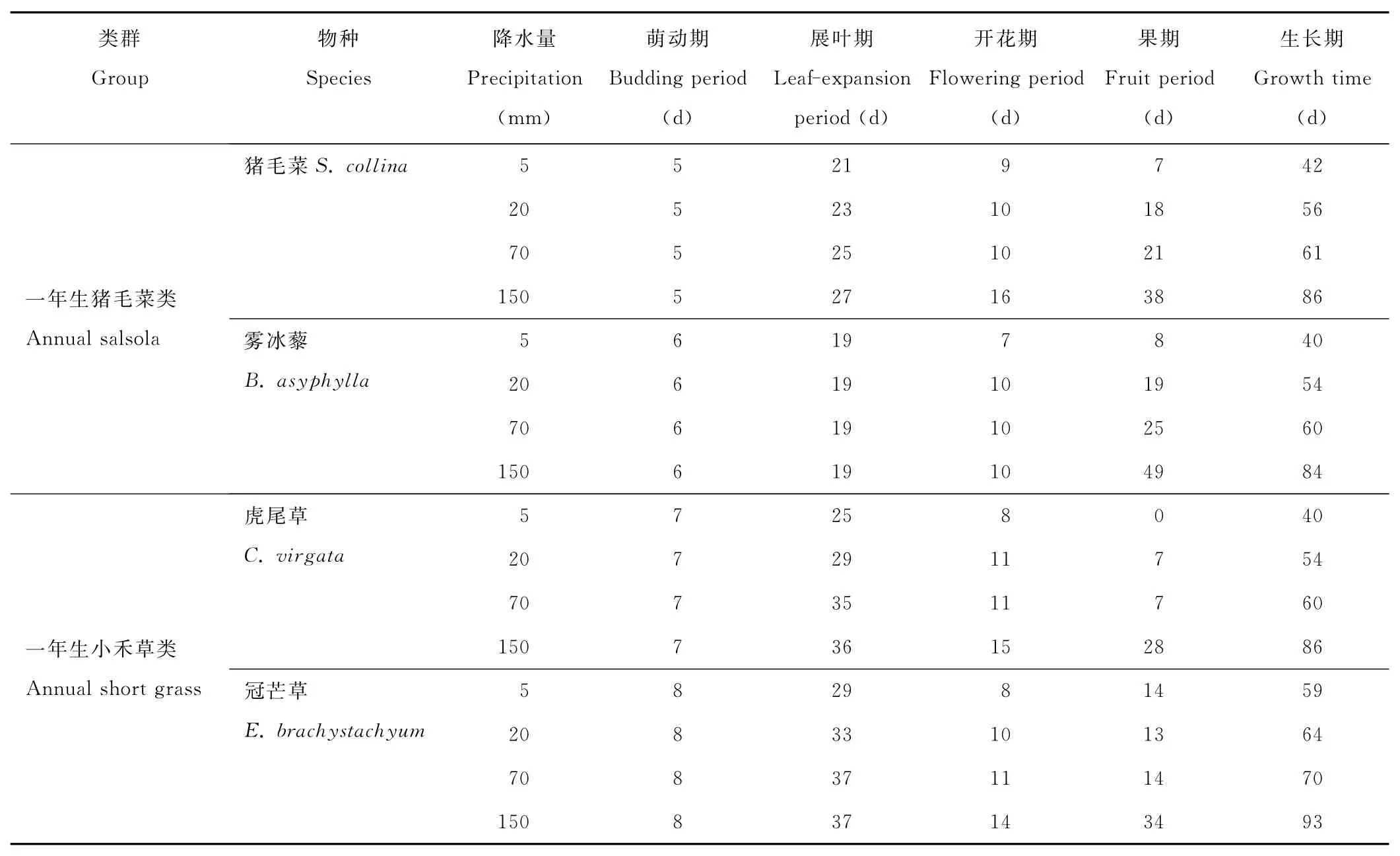
表2 4個降水梯度下一年生植物的物候期Table 2 Phenological phases of annual plants in four rainfall grads
2.2 株高變化
一年生植物各類群植株高度均隨降水量增加而增加,其中一年生豬毛菜類各梯度之間的差異均達到顯著水平,一年生小禾草類除5和20mm降水條件下差異不顯著外(P>0.05),其余均差異顯著(P<0.05)(圖1a)。但20mm以下降水梯度其株高變幅較小,株高生長主要表現在70mm以上較充足的降水條件下,在枯死前的最后1周即第8周達到最大值(圖2a)。
4個降水梯度中不同類群一年生植物株高在苗期并無顯著差異,但隨著營養生長的加快,一年生豬毛菜類高于一年生小禾草類,并且差異顯著(P<0.05)。到生殖生長后期,一年生小禾草類由于生殖枝的存在,株高高于一年生豬毛菜類,并且差異顯著(P<0.05)(圖2a)。
2.3 葉長變化
4個降水梯度一年生植物的葉長除一年生豬毛菜類表現為5mm降水條件下最長外,其余均表現為隨降水量增加而增加。但一年生豬毛菜類5和150mm降水條件下差異不顯著,其余均差異顯著(P>0.05);一年生小禾草類5和20mm降水條件下差異不顯著(P>0.05)外,其余均差異顯著(P<0.05)(圖1b)。
苗期一年生豬毛菜類和一年生小禾草類的葉長并無顯著差異。但此后一年生豬毛菜類的葉長幾乎不再增長;而一年生小禾草類的葉長不斷增長,苗期以后均顯著長于一年生豬毛菜類(P<0.05)(圖2b)。
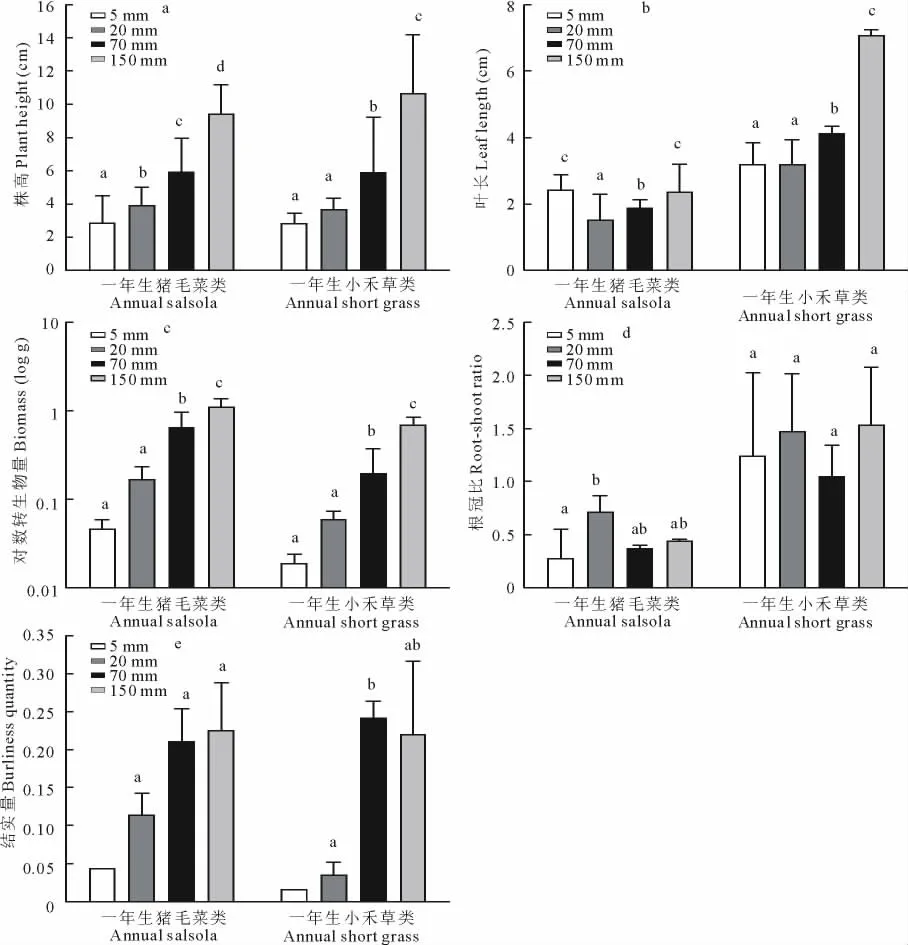
圖1 不同降水梯度下一年生植物性狀特征(單因素方差分析)Fig.1 The characters of annual plant traits between in four rainfall grads(One-way ANOVA)
2.4 生物量變化
一年生植物生物量均表現為隨降水量增加而增加,其中一年生豬毛菜類除5和20mm降水條件下差異不顯著(P>0.05)外,其余差異均顯著(P<0.05);而一年生小禾草類除150mm降水條件下的生物量顯著(P<0.05)大于其他3個降水梯度外,其余各梯度之間的差異均不顯著(P>0.05)。最小降水量(5mm)單株生物量非常低,在0.1g以下,僅為0.065g(圖1c)。
4個降水梯度,一年生豬毛菜類的生物量始終大于一年生小禾草類的生物量,差距隨生長時間的增長而增大,并且在5和20mm降水條件下差異顯著(P<0.05)(圖2c)。
2.5 根冠比變化
2.6 結實量
4個降水梯度下的一年生豬毛菜類和一年生小禾草類均能結實,能夠正常完成生活史。一年生植物結實量大致表現為隨降水量增加而增加(圖1e),特別是一年生豬毛菜類結實量表現為150mm>70mm>20mm>5 mm,但各梯度結實量之間差異不顯著(P>0.05)。一年生小禾草類的結實量表現為70mm降水條件下最大,150mm降水條件下次之,5mm降水條件下最少,除20和70mm降水條件下的結實量差異顯著外(P<0.05),其余梯度間差異均不顯著(P>0.05),在5mm的降水條件下,因為植物個體存活較少,不符合統計學原理,所以沒有顯著性分析。
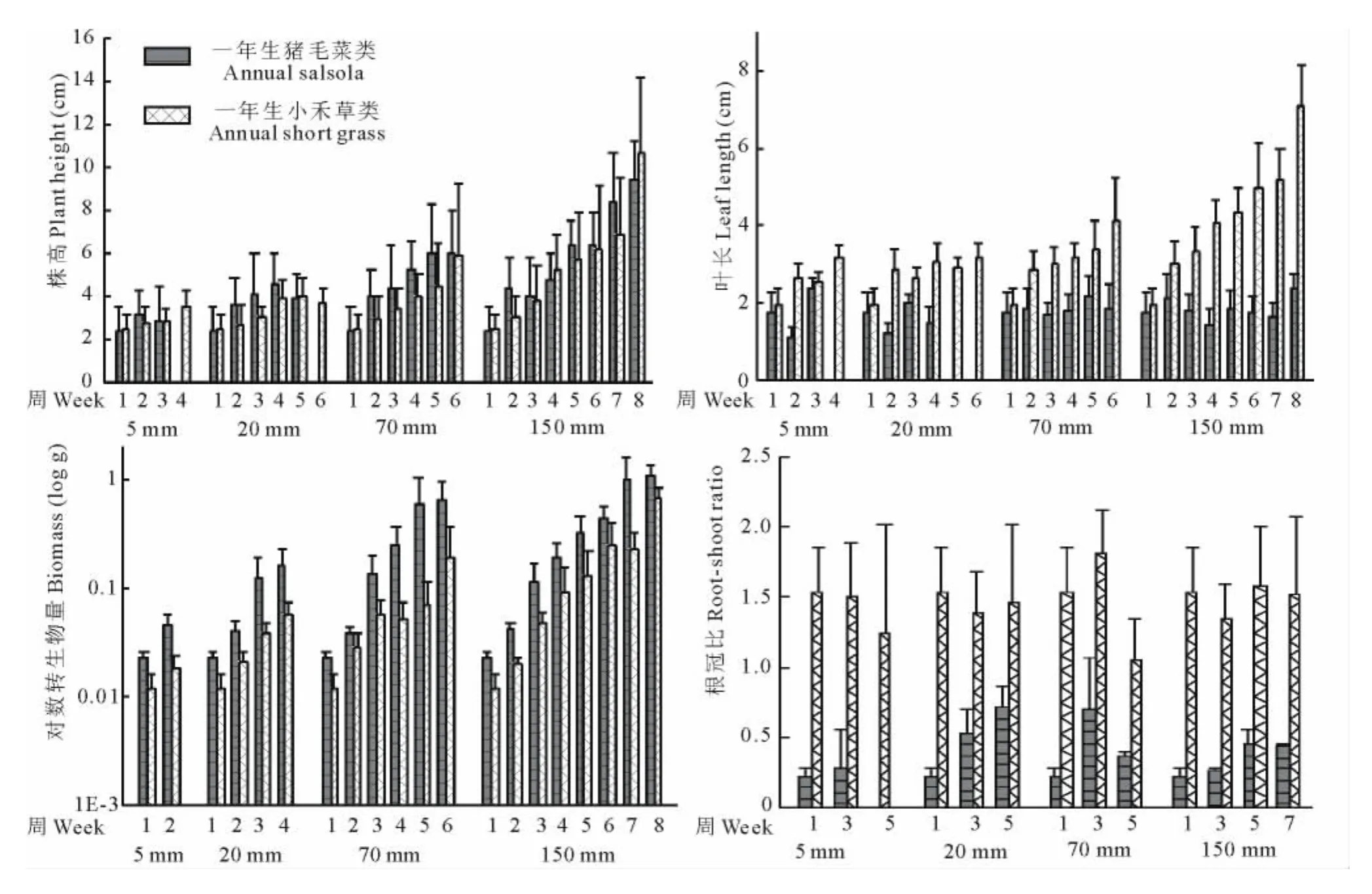
圖2 不同降水條件下一年生豬毛菜類和一年生小禾草類植物性狀差異(獨立樣本T檢驗)Fig.2 Plant trait differences between annual salsolas and annual short grasses in four rainfall grads(Independent sample Ttest)
3 討論
3.1 一年生植物生長期特征
溫室栽培降水控制實驗表明草原與荒漠一年生植物生長期一般為40~90d,通常不超過100d。5mm最低降水量植物生長期大部分為40d左右,150mm最高降水量大部分為85d左右。這一結果與草原和荒漠一年生植物生長期通常不超過90d,最短時僅40~50d的野外經驗觀察基本一致。在自然條件下,由于降水的不確定性,通常很難準確觀察到一年生植物層片的確切生長期或壽命。本實驗中的150mm降水量,在草原區是較高的有效降水量,通常在全年降水量約350mm左右的典型草原區亦很難達到完全被一年生植物吸收利用的150mm有效降水,這一降水條件已達到草甸草原降水條件。因此本實驗較準確地確定了我國北方溫帶草原與荒漠一年生植物在較高降水和較低降水條件下的生長期。
3.2 一年生植物對水分的高效利用
梁存柱等[10]認為一年生植物有很高的水資源利用效率,對水分脅迫和水分供應高度敏感,能根據水分資源量的多少,調節自己的生育周期和生長勢,以適應干旱與半干旱地區降水波動環境特點,但這一認識一直沒有可靠的實驗證據。本實驗結果表明,從5mm到150mm的4個降水梯度一年生植物大多能夠開花結實,正常完成生活史,并在最少降水條件下利用最短時間完成全部生活史,雖然保證了基本的后代延續,卻犧牲了營養生長和結實量;而在較高降水條件下不僅有很好的營養生長,并能夠大量繁殖后代,由此驗證了上述觀點。
實驗結果表明,在5mm有效降水條件下,大部分物種在6周左右完成其從萌發到開花結果的全部生活史,而較高降水條件下也僅有3個月的生長期,表現出干旱區短命植物的特點。在亞洲中部荒漠區,即使只有苗期得到有限的降水,生殖生長期后不再降水,一年生植物也可以利用土壤中蓄積的水分完成生活史。研究結果同時也表明干旱脅迫將使一年生植物的花果期提前,在水分嚴重虧缺來臨之前及時完成生活史,首先保證其后代的延續,達到植物避旱的目的,這與Franks和Weis[35]的研究結論一致。干旱與半干旱區一年生植物這一高度順應氣候波動的特點,早已被當地牧民掌握并加以利用。內蒙古草原與荒漠一年生植物層片,又稱為“夏雨型”一年生植物層片,在干旱的荒漠區也稱“熱草”或“雨草”,是借用當地蒙古族牧民對生長在蒙古高原中西部一批一年生短命植物的專用名稱“呼熱嘎日吉”而來,即雨后馬上長出的一年生草類。“熱草”一詞形象地把生長在荒漠草原、草原化荒漠和典型荒漠群落的一年生植物層片的生態—生物學特征概括出來,它完全符合當地生態、氣候特征:降水來得較遲,通常只有到夏季最熱的月份(6月下旬到7月上旬),才能得到少量的降水(80~200mm)。不多的降水,幾乎成為當地植物生長發育的唯一水分來源。有效的降水加上適宜的溫度(18~30℃),為夏雨型一年生植物提供了良好的生長發育條件,并能夠最充分、最經濟地利用降水。因此,當地牧民常以一年生植物長勢好壞來衡量、預測當年小畜的膘情和渡過冬春能力。
3.3 不同類群一年生植物對降水變化的響應
干旱與半干旱區水分是影響植物生長與生存的主要限制因子,荒漠植物的適應特征都與水資源的利用有關[36]。有限的降水直接限制一年生植物性狀的表達,所有植物都必然面臨著繁殖、生長和維持三方面的資源分配權衡[37-39]。故資源分配模式在很大程度上反映了植物生活史特征,而生活史對策就是這種不同功能間權衡資源分配的綜合結果[40,41]。一年生植物性狀對水分變化的響應,正是這一類群植物長期適應干旱的結果。
一年生豬毛菜類在5mm降水條件下株高、生物量以及結實量最小,葉長與根冠比最大(圖1和2),通過減少葉片數來提高水分利用效率,不斷增加對吸收器官的投入,以從土壤中吸收盡可能多的水,來完成生活史。20 mm降水條件下,隨生長時間的增加根冠比持續增加,說明植物生長仍受水的限制,仍然將較多能量投入到水分吸收上。70mm降水條件下,隨生長時間的增加根冠比先增后減,說明一年生豬毛菜類在生長期先發展吸收器官;后期,在充足的水分條件下吸收器官已能滿足營養器官生長和結實的需要,且結實量相對較高。150mm降水條件下,生長期內根冠比幾乎無變化,說明在水分豐富的條件下,一年生豬毛菜類分配給吸收器官和營養生殖器官的能量比例幾乎不變,其結實量也與70mm降水條件下的結實量相當,表明其過多的水分已對其生長無太大的作用。
一年生小禾草類在5,20,70和150mm降水條件下,其根冠比均大于1,說明對吸收器官的投入均高于對營養器官和生殖器官的投入。出現這一現象的原因可能與一年生小禾草長期適應干旱環境有關。在內蒙古草原與荒漠區,一年生小禾草的分布通常比一年生豬毛菜類分布更靠西,一年生小禾草主要分布于荒漠區及草原化荒漠區,而一年生豬毛菜類主要分布于典型草原區。由于長期適應干旱環境,增加根系的生長已保證其吸收更多的水分。因此即使栽培條件下有充足的水分,遺傳因素導致其維持植物固有屬性。這一推測有待于進一步實驗驗證。
自然條件下不同類群一年生植物根據降水量多少產生的性狀各不相同,即不同植物類群對降水響應的敏感程度各異。本研究發現,一年生豬毛菜類在不同降水條件下物候期長短的變化比一年生小禾草類小(表2)。一年生小禾草類能根據降水量變化及時調整各物候期長短以適應生長和繁殖;而一年生豬毛菜類,傾向于以調整生殖生長期長短的方式,即以減少結實量為代價加速種子成熟的方式響應降水量不足。并且,相同降水條件下小禾草類植物的生長期相對較長并且種子能夠發育成熟。因此,豬毛菜類植物表現出更強的避旱能力,而小禾草類植物則表現出相對更強的耐旱能力。
3.4 一年生植物性狀對降水梯度變化的響應
有研究表明,水分的異質性能顯著影響植物的形態特征,主要表現在植物的生物量和生物量分配格局以及植物的高度等方面[42]。在沒有脅迫和干擾生境中的植物具有較大的根冠比;而在脅迫環境下植物將采取降低生物量,形體變小,生長速率減緩等生長策略來應對脅迫[43,44]。2類一年生植物在降水較少時(5和20mm)都采用增大根冠比,降低株高、減少葉片數等減小營養器官的生長的策略來適應干旱環境。在70mm降水條件下,資源分配達到最佳化,此時具有最小的根冠比,并獲得了最大的結實量。減少的水分供應迫使植物調整資源配比,將更多的能量投入給吸收器官,并且這種投入隨水分的減少而增加。相反,增加水分供給(150mm),并沒有獲得更高的結實量,反而不利于資源的合理分配,雖獲得較高的產量,但不利于物種的延續。
4 結論
綜上所述,通過在溫室進行的不同梯度降水控制實驗發現:
1)草原與荒漠一年生植物類群生長期通常在40~90(<100)d。
2)隨降水梯度的增加,一年生植物的生長期、植物高度、葉長、生物量等均增加,5mm有效降水雖然能基本完成生活史,但生產能力極低,在70mm有效降水條件下資源分配達到最佳化,具有最小的根冠比,并獲得了最大的結實量,而150mm有效降水生物量與結實量等無顯著增加,表現出一定資源浪費現象。
3)不同類群一年生植物對降水變化及適應方式不同,一年生豬毛菜類在不同降水條件下的物候期長短的變化比一年生小禾草類小,傾向于以調整生殖生長期長短,以減少結實量為代價,加速種子成熟的方式響應降水量的不足;一年生小禾草類能根據降水量變化及時調整各物候期長短以適應生長和繁殖;相同降水條件下小禾草類植物的生長期相對較長。在適應方式上,豬毛菜類植物表現出更強的避旱能力,而小禾草類植物則表現出相對更強的耐旱能力。
[1] Violle C,Navas M L,Vile D,etal.Let the concept of trait be functional![J].Oikos,2007,116(5):882-892.
[2] 周道瑋.植物功能生態學研究進展[J].生態學報,2009,29(10):5644-5655.
[3] McIntyre S,Lavorel S,Landsberg J,etal.Disturbance response in vegetation - towards a global perspective on functional traits[J].Journal of Vegetation Science,1999,10(5):621-630.
[4] Cornelissen J H C,Lavorel S,Garnier E,etal.A handbook of protocols for standardised and easy measurement of plant functional traits worldwide[J].Australian Journal of Botany,2003,51(4):335-380.
[5] 孟婷婷,倪健,王國宏.植物功能性狀與環境和生態系統功能[J].植物生態學報,2007,31(1):150-165.
[6] Reich P B,Wright I J,Cavender-Bares J,etal.The evolution of plant functional variation:Traits,spectra,and strategies[J].International Journal of Plant Sciences,2003,164(3Supplement):143-164.
[7] Cheplick G P.Life history trade-offs in amphibromus scabrivalvis(Poaceae):allocation to clonal growth,storage,and cleistogamous reproduction[J].American Journal of Botany,1995,82(5):621-629.
[8] Arndt S K.Integrated research of plant functional traits is important for the understanding of ecosystem processes[J].Plant and Soil,2006,285(1-2):1-3.
[9] 梁存柱,王煒,朱宗元,等.荒漠區一年生植物層片的組織格局與生態適應模式[J].干旱區資源與環境,2002,16(1):77-83.
[10] 梁存柱,劉鐘齡,朱宗元,等.阿拉善荒漠區一年生植物層片物種多樣性及其分布特征[J].應用生態學報,2003,14(6):897-903.
[11] 李雪華,李曉蘭,蔣德明,等.干旱半干旱荒漠地區一年生植物研究綜述[J].生態學雜志,2006,25(7):851-856.
[12] 拉甫連科.關于亞洲中部草原植物群落的某些突出特征[J].蘇聯植物學雜志,1973,58(11):1603-1607.
[13] 尤納托夫.蒙古人民共和國植被的基本特點[M].李繼侗,譯.北京:科學出版社,1959.
[14] Madon O,Medail F.The ecological significance of annuals on a Mediterranean grassland(Mt Ventoux,France)[J].Plant E-cology,1997,129(2):189-199.
[15] Gonzalez-Astorga J,Nunez-Farfan J.Variable demography in relation to germination time in the annual plant Tagetes micrantha Cav.(Asteraceae)[J].Plant Ecology,2000,151(2):253-259.
[16] Brown G.Species richness,diversity and biomass production of desert annuals in an ungrazedRhanteriumepapposumcommunity over three growth seasons in Kuwait[J].Plant Ecology,2003,165(1):53-68.
[17] Krieger A,Porembski S,Barthlott W.Temporal dynamics of an ephemeral plant community:species turnover in seasonal rock pools on Ivorian inselbergs[J].Plant Ecology,2003,167(2):283-292.
[18] Robinson M D.Growth and abundance of desert annuals in an arid woodland in Oman[J].Plant Ecology,2004,174(1):137-145.
[19] 中國科學院內蒙古寧夏綜合考察隊.內蒙古植被[M].北京:科學出版社,1985.
[20] 中國科學院內蒙古寧夏綜合考察隊.內蒙古自治區及其東西部毗鄰地區天然草場[M].北京:科學出版社,1980.
[21] 張德魁,馬全林,劉有軍,等.河西走廊荒漠區一年生植物組成及其分布特征[J].草業科學,2009,26(12):37-41.
[22] 陶冶,張元明.3種荒漠植物群落物種組成與豐富度的季節變化及地上生物量特征[J].草業學報,2011,20(6):1-11.
[23] 王宗靈,徐雨清,王剛.沙區有限降水制約下一年生植物種子萌發與生存對策研究[J].蘭州大學學報,1998,34(2):102-107.
[24] 張繼恩,梁存柱,付曉玥,等.阿拉善荒漠一年生植物種子萌發特性及生態適應性分析[J].干旱區資源與環境,2009,23(2):175-179.
[25] 楊磊,王彥榮,余進德.干旱荒漠區土壤種子庫研究進展[J].草業學報,2010,19(2):227-234.
[26] 劉有軍,紀永福,馬全林,等.溫度和光照對3種一年生植物種子萌發的影響[J].中國生態農業學報,2010,18(4):810-814.
[27] 劉有軍,劉世增,紀永福,等.碟果蟲實種子萌發對策及生態適應性[J].生態學報,2010,30(24):6910-6918.
[28] 王兮之,張景光,施溯筠,等.人工固沙區一年生植物小畫眉草的競爭調節[J].中國沙漠,2001,21(3):26-29.
[29] 張景光,李新榮,王新平,等.沙坡頭地區固定沙丘一年生植物小畫眉草種群動態研究[J].中國沙漠,2001,21(3):232-235.
[30] 張景光,周海燕,王新平,等.沙坡頭地區一年生植物的生理生態特性研究[J].中國沙漠,2002,22(4):350-353.
[31] 徐彩琳,李自珍.荒漠一年生植物小畫眉草的種群動態調節與模擬[J].西北植物學報,2002,22(6):1415-1420.
[32] 寇祥明,楊利民,姜雷,等.五葉地錦幼苗生長和生理生態特性對模擬降水量變化的響應[J].吉林農業大學學報,2006,28(5):521-524.
[33] 宛敏渭,劉秀珍.中國物候觀測方法[M].北京:科學出版社,1979.
[34] 劉曉軍,洪光宇,袁志誠,等.干熱脅迫下兩種葦狀羊茅對不同水肥處理的響應機理[J].草業學報,2011,20(1):46-54.
[35] Franks S J,Weis A E.A change in climate causes rapid evolution of multiple life-history traits and their interactions in an annual plant[J].Journal of Evolutionary Biology,2008,21(5):1321-1334.
[36] 張景光,王新平,李新榮.荒漠植物生活史對策研究進展與展望[J].中國沙漠,2005,25(3):306-314.
[37] 張大勇.理論生態學研究[M].北京:高等教育出版社,2000.
[38] 張大勇.植物生活史進化與繁殖生態學[M].北京:科學出版社,2004.
[39] 王斌世,張榮.半干旱區農田雜草的生活史對策研究[J].草業學報,2011,20(1):257-260.
[40] Bostock S J,Benton R A.The reproductive strategies of five perennial Copositea[J].Journal of Ecology,1979,67:91-107.
[41] Jacob W.Allocation,plasticity and allometry in plants[J].Perspectives in Plant Ecology,Evolution and Systematics,2004,6(4):207-215.
[42] 何維明.水分因素對沙地柏實生苗水分和生長特征的影響[J].植物生態學報,2001,25(1):11-16.
[43] Dong M.Morphological responses to local light conditions in clonal herbs from contrasting habitats,and their modification due to physiological integration[J].Oecologia,1995,101(3):282-288.
[44] 武高林,杜國禎.植物形態生長對策研究進展[J].世界科技研究與發展,2007,29(4):47-51.
猜你喜歡
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
