TA29-Barnase轉基因雄性不育芥菜的獲得
2013-02-24 09:01:32楊海鵬宋洪元
中國蔬菜 2013年4期
王 倩 楊海鵬 鄒 敏 宋洪元
(西南大學園藝園林學院,南方山地園藝學教育部重點實驗室,重慶 400715)
芥菜〔Brassica juncea(L.)Czern.et Coss.〕是我國特產蔬菜之一,營養豐富,經濟價值較高,在我國南、北方均有種植,是重要的加工類蔬菜。在芥菜生產實踐中廣泛存在著諸如病毒病、先期抽薹、加工品質等問題(楊景華 等,2006),植物雜種優勢的利用可以在一定程度上解決這些問題。由于芥菜類作物自交親和指數非常高,雜種優勢育種難以通過自交不親和途徑得以實現,導致長期以來芥菜類蔬菜品種選育都是以常規品種為主。雖然近年來利用細胞質雄性不育在芥菜優勢育種上取得了一定的進展,但如何發掘、創造優良的芥菜雄性不育材料仍是當前芥菜雄性不育研究的重要內容之一。隨著基因工程雄性不育系成功在油菜、玉米、菊苣等農作物上得到商業化應用(Kempken,2010),圍繞花粉的發育過程,已通過多種途徑建立了植物工程雄性不育系。利用具有特異時空表達特性的花藥絨氈層特異啟動子與解淀粉芽孢桿菌的Barnase核糖核酸酶基因融合,導入植物體內調控花粉發育,從而獲得雄性不育材料成為其中應用最為普遍的方法。該方法分別在花椰菜(Reynaerts et al.,1993)、煙草(李勝國 等,1995)、油菜(鐘蓉 等,1996)、甘藍(沈革志 等,2001)、番茄(白玲 等,2002)、菜薹(曹必好和孟成民,2008)等作物上成功獲得了雄性不育材料。基于對芥菜細胞質雄性不育機制的研究,Yang 等(2010)將來自線粒體的編碼基因orf220導入莖用芥菜中獲得雄性不育材料。相對于其他農作物,有關芥菜植物基因工程雄性不育的研究報道較少,為拓寬芥菜雄性不育資源,本試驗將煙草絨氈層TA29啟動子驅動下的Barnase基因通過農桿菌介導法轉入莖用芥菜,獲得了轉基因雄性不育芥菜植株,從分子水平、細胞學水平和結籽情況等方面調查分析了轉基因植株的育性,結果顯示Barnase基因在芥菜絨氈層特異表達后能導致徹底的雄性不育,但伴隨花朵變小,雌蕊、種莢變短,結籽數量下降的現象。
1 材料與方法
1.1 質粒和菌株
含雄性不育基因的表達載體pCABarTA29-Bn的農桿菌(Agrobacterium tumefaciens)EHA105 由南方山地園藝學教育部重點實驗室保存(圖1)。
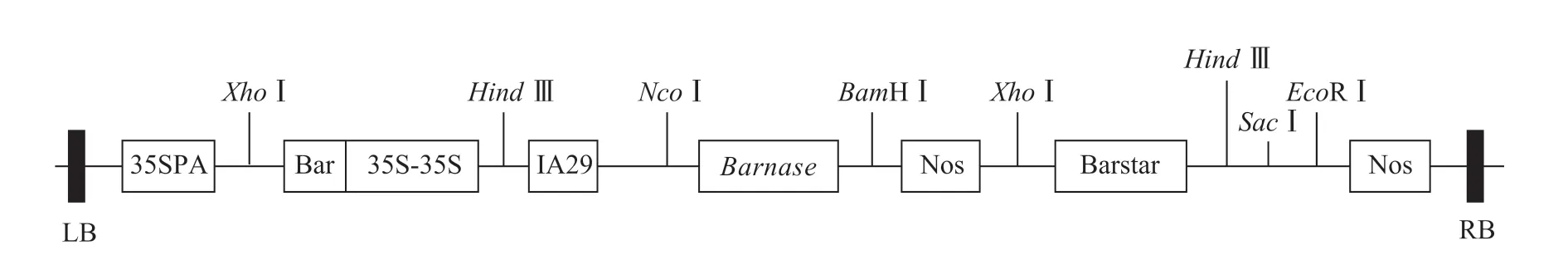
圖1 pCABarTA29-Bn 植物表達載體
1.2 試驗材料
試驗于2012年2月7日~7月15日在西南大學南方山地園藝學教育部重點實驗室進行。供試芥菜品種為渝豐榨菜,由重慶科光種苗有限公司提供。
TaqDNA 聚合酶和DNA marker 購自全式金生物工程有限公司;羧芐青霉素(Carbenicilin)、卡那霉素(Kanamycine)、鏈霉素(Streptomycin)、吲哚乙酸(IAA)、6 芐基腺嘌呤(6-BA)以及細菌培養基均購自北京鼎國生物技術公司;植物DNA 提取、植物組織培養以及顯微制片的各種化學藥品均為國產分析純。除草劑Basta(草丁膦,主要成分為PPT)購自日產化學工業株氏會社。
1.3 轉基因芥菜植株的獲得
采用農桿菌介導的葉盤轉化法(王關林和方宏筠,2002;曹必好 等,2003)進行轉化,并略有改動:取苗齡7 d 的渝豐榨菜幼苗的下胚軸,切成1 cm 長的莖段,在分化培養基(MS+2 mg·L-16-BA+0.2 mg·L-1NAA+3%蔗糖+0.6%瓊脂)上預培養2 d,用濃度OD600=0.5~0.6的農桿菌菌液侵染10 min,轉到分化培養基上共培養2 d,隨后在篩選培養基(MS+2 mg·L-16-BA+0.2 mg·L-1NAA+8 mg·L-1PPT+400 mg·L-1Carb+3%蔗糖+0.6%瓊脂)上進行多次篩選,每14 d 換1次培養基,最后將再生植株轉到生根培養基(MS+0.2 mg·L-1NAA+8 mg·L-1PPT+400 mg·L-1Carb+3%蔗糖+0.6%瓊脂)上進行生根培養。
1.4 轉基因芥菜植株的分子檢測
采用改進的CTAB 法提取芥菜總DNA(宋洪元 等,2010)。以提取的總DNA 為模板,用TA29啟動子上游引物(TA29-1:5′-CGCGGTACCCCAAGATTGCATAAG-3′)和Barnase基因下游引物(BN-2:5′-CCCCTCGGATCCGTTATCTGATCTTTGTA-3′)進行TA29啟動子及下游Barnase基因的PCR檢測。反應體系:模板DNA 1μL(30 ng·μL-1),上下游引物各1μL (10 ng·μL-1),dNTP Mixture 0.2μL (2.5 mmol·L-1),MgCl22.0 μL(25 mmol·L-1),TaqDNA 聚合酶0.3 μL(5 U·μL-1),10×Buffer 2.5 μL,ddH2O 補至總體積為25 μL。PCR 擴增反應程序:94℃預變性5 min;94℃變性40 s,56℃退火40 s,72℃延伸2 min,30個循環;72℃延伸5 min。擴增產物用1.0%瓊脂糖凝膠電泳檢測。
1.5 轉基因芥菜植株花器官的觀察和授粉試驗
取轉基 因以及野生型芥菜植株當天開放的花,觀察比較雄蕊和花藥的形態結 構。分別調查雄性不育轉基因芥菜植株自交以及用野生型芥菜植株的花粉進行人工授粉后的結籽情況。
1.6 轉基因芥菜植株雄性不育花藥、花粉發育的細胞學觀察
取芥菜轉基因雄性不育株和野生型可育株的不同大小花蕾,投入FAA 固定液固定至少24 h,之后按照常規石蠟法制作切片(鄒瑞昌 等,2012):各級酒精脫水→透明→浸蠟→包埋→切片→展片→脫蠟→染色,最后在LEICACTR5000 顯微鏡下鏡檢并照相。
2 結果與分析
2.1 轉基因芥菜植株的獲得
芥菜幼苗下胚軸經農桿菌菌液浸染后,在含除草劑有效成分8 mg·L-1PPT 的培養基上進行多輪篩選,篩選過程中非轉化個體逐漸黃化死亡,抗性個體成苗并正常生根,生根后移栽到土壤中,獲得5株成活轉基因芥菜植株(圖2)。
提取成活植株DNA,利用引物TA29-1 和BN-2 進行PCR 擴增,在5株轉基因植株中均獲得了約630 bp 的DNA片段(圖3),表明外源目的基因已整合到芥菜植物基因組中。

圖2 轉化過程中芥菜植株生根和移栽成苗
2.2 轉基因芥菜植株的花器官形態
盡管獲得的轉基因芥菜植株與野生型植株形態間無明顯差異,但開花時轉基因植株花朵偏小,并伴隨花瓣較小、雌蕊短且粗、雄蕊短縮的現象;而野生型植株則表現為花瓣大、雌蕊細長、雄蕊粗壯(圖4)。同時,轉基因植株花藥瘦小、干癟、無花粉囊及花粉;野生型植株則花藥飽滿,花粉囊充滿大量花粉(圖5)。
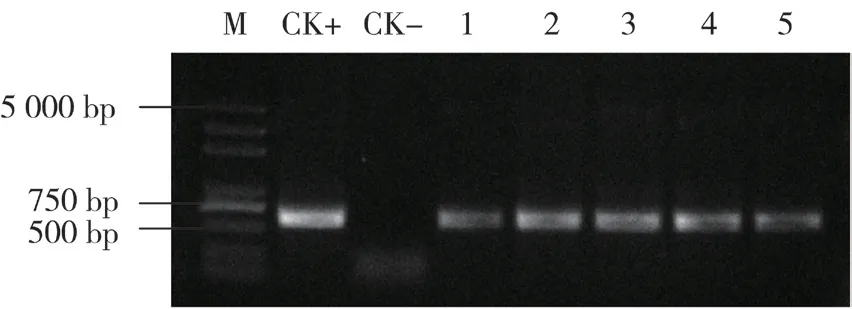
圖3 轉基因芥菜植株的PCR檢測結果

圖4 轉基因芥菜植株花器官的形態結構

圖5 轉基因芥菜植株花藥的形態結構
2.3 轉基因芥菜植株的花藥發育過程
在TA29啟動子的作用下,Barnase基因在花藥絨氈層組織細胞內表達,產生水解絨氈層組織細胞內的RNA 的核糖核酸水解酶,阻礙花藥絨氈層的發育,從而導致雄性不育的產生。顯微切片結果顯示(圖6),花藥發育早期,轉基因芥菜植株與野生型植株均在花藥的4個角隅處形成正常的藥室內壁、中層、絨氈層和造孢細胞,呈蝴蝶形排列(圖6-1,6-4)。野生型植株進入減數分裂期,花粉母細胞變為橢圓形,絨氈層細胞體積增大;進入單核初期,剛釋放出來的游離小孢子形狀不規則,細胞壁很薄;單核晚期,細胞壁逐漸加厚且趨于圓球形,絨氈層開始降解(圖6-2);單核小孢子發生有絲分裂后,進入二核期,絨氈層只余殘跡,生殖核再次有絲分裂形成兩個精核,花粉粒趨于成熟,絨氈層最后在花藥即將開裂釋放花粉粒時已完全消失(圖6-3)。而轉基因植株的花藥中,當花粉母細胞開始進入減數分裂時,藥室內壁的絨氈層細胞已經開始降解,并伴隨花粉母細胞的退化(圖6-5);減數分裂后絨氈層細胞和花粉母細胞進一步退化解體,無小孢子母細胞釋放,藥室呈現空洞狀(圖6-6)。
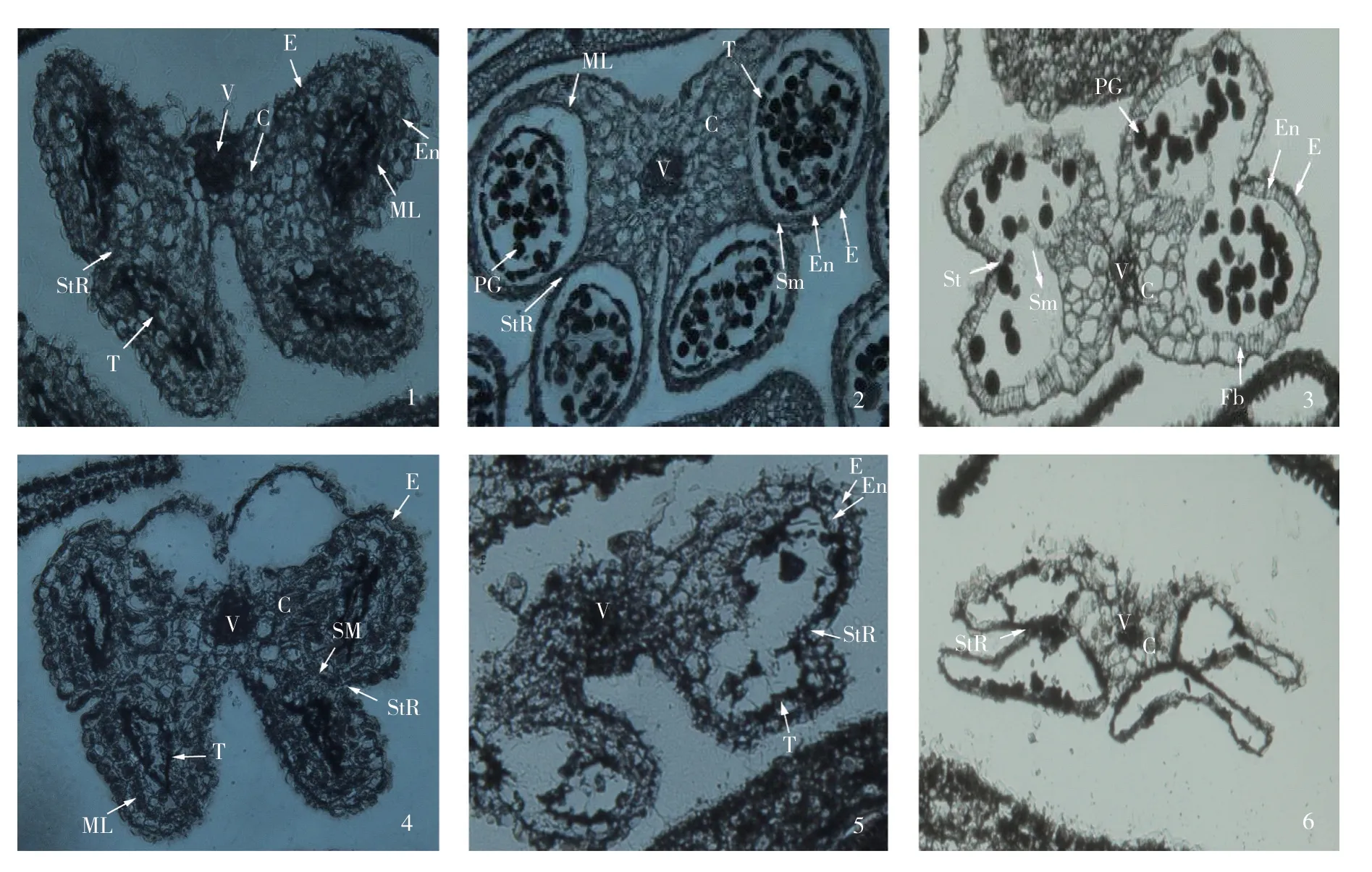
圖6 轉基因芥菜植株的花藥發育過程
2.4 轉基因芥菜植株的結實性
轉基因雄性不育芥菜植株開花后,自交花朵的果莢基本不能膨大,部分脫落,無種子形成(圖7-A1),而野生型植株則結籽正常(圖7-B)。轉基因植株用野生型植株的花粉授粉后,基本均能結實(圖7-A2),但與野生型植株(圖7-C2)相比果莢較短,不飽滿,且種子數少(圖7-C1)。該結果顯示煙草TA29啟動子驅動Barnase基因在芥菜植株的花藥絨氈層表達后,除了影響絨氈層細胞的發育從而獲得雄性不育外,對雌蕊以及其他花器官的發育也有一定的影響。

圖7 轉基因芥菜植株的結籽情況
3 結論與討論
TA29是從煙草中克隆的一個絨氈層特異表達基因,在煙草減數分裂期到小孢子有絲分裂期的絨氈層中特異表達,在其他器官和花的其他組織中均沒有表達 (Koltunow et al.,1990)。該啟動子驅動Barnase基因在絨氈層中表達后獲得了煙草和油菜雄性不育材料(Mariani et al.,1990)。隨后基于TA29-Barnase雄性不育的轉基因油菜獲得商業化應用。然而,后來有研究顯示TA29-Barnase基因導致的雄性不育對溫度敏感,在溫度升高后轉基因植株出現不同程度的育性恢復現象。如在轉基因煙草和油菜的研究中發現,當溫度低于20℃時不育性穩定,溫度升高到25℃時開始轉育,30℃時大部分可育(鐘蓉 等,1996)。另外,TA29-Barnase基因轉化甘藍后發現雄性不育植株的園藝學性狀與未轉化植株相同,不育植株的花朵表現為雄蕊完全退化,但蜜腺和雌蕊健全,能接受外來花粉,雜交結實率較高,但是轉基因植株中存在全不育植株和半不育植株,半不育植株在開花中后期出現育性恢復現象(沈革志 等,2001)。該基因在 菜薹中表達后產生的雄性不育植株在30~35℃/25~28℃的高溫條件下自交全部不能結籽,沒有育性恢復現象發生(曹必好和孟成民,2008)。本試驗利用該基因獲得的芥菜雄性不育植株的生長發育全程處于控溫條件下的人工氣候室內,其雄性不育性是否受田間溫度變化的影響有待進一步研究。通過原位雜交和啟動子報告基因的轉基因分析顯示,TA29基因專一性地在絨氈層細胞中表達,表達時期為減數分裂期到花粉壁增厚期(-1 到+6 期)(Koltunow et al.,1990)。但在棉花中的研究結果顯示,該啟動子除了在棉花花藥中優勢表達外,其控制的基因也會在棉花的葉片表皮毛和花粉中表達(尹夢回 等,2008)。同時,TA29-Barnase轉基因棉花除花藥干癟、花粉無活力外,也表現出花朵變小、花絲變短等現象(Zhang et al.,2007),這與本試驗所獲得的芥菜雄性不育轉基因植株的花器官類似。但本試驗獲得的雄性不育芥菜植株除了上述特點外,還出現了雌蕊偏短、種莢變短的現象,說明TA29-Barnase基因在芥菜中表達后,對非靶標組織的影響相對其他已報道的植物更大。因此,在利用Barnase基因創建芥菜雄性不育系時,有必要尋找更為嚴謹的絨氈層啟動子表達Barnase基因,減少對花器官中非靶標細胞、組織發育的影響,獲得結籽能力正常的雄性不育系,從而實現一代雜種的經濟有效地繁育。
白玲,劉凡,李鎖平,曹鳴慶.2002.農桿菌介導Barnase基因轉化番茄.河南大學學報:自然科學版,32(3):16-19.
曹必好,雷建軍,宋洪元,秦耀國.2003.芥菜農桿菌高效遺傳轉化體系初步建立.華南農業大學學報:自然科學版,4(4):48-51.
曹必好,孟成民.2008.TA29-Barnase基因轉化菜心.生物工程學報,25(5):881-886.
李勝國,劉玉樂,朱鋒,羅玉英,康良儀,田波.1995.基因工程雄性不育煙草的獲得.植物學報,37(8):659-660.
沈革志,王新其,朱玉英,楊紅娟,陸桂華,王江,宛新衫,張景六.2001.TA29-Barnase基因轉化甘藍產生雄性不育植株.植物生理學報,27(1):43-48.
宋洪元,任雪松,司軍,李成瓊,宋明,雷建軍.2010.利用Cre/lox 重組系統獲得煙草TA29-Barnase基因工程雄性不育恢復系.農業生物技術學報,18(3):468-475.
王關林,方宏筠.2002.植物基因工程.北京:科學出版社:742-744.
楊景華,張明方,喻景權,陳竹君.2006.莖用芥菜細胞質雄性不育系及其保持系SPOROCYTELESS基因研究.園藝學進展,(7):416-419.
尹夢回,董靜,李先碧,侯磊,羅明,李德謀,裴炎,肖月華.2008.煙草絨氈層特異啟動子pTA29在棉花中的表達特性.作物學報,34(12):2092-2098.
鐘蓉,劉玉樂,朱鋒,李勝國,康良儀,羅鵬.1996.TA29-Barnase基因導致油菜雄性不育的研究.植物學報,38(7):582-585.
鄒瑞昌,萬正杰,徐躍進,楊文杰,傅延棟.2012.新型葉用芥菜細胞質雄性不育系0912A 的花藥發育特征.華中農業大學學報,31(1):44-49.
Kempken F.2010.Engineered male sterility.Genetic Modification of Plants-Biotechnology in Agriculture and Forestry,64(2):253-265.
Koltunow A M,Truettner J,Cox K H,Wallroth M,Goldberg R B.1990.Different temporal and spetial gene expression patterns occur during anther development.Plant Cell,2:1201-1224.
Mariani C,de Beuckeleer M,Truettner J,Leemans J,Goldberg R B.1990.Induction of male sterility in plants by a chimaeric ribonuclease gene.Nature,347:737-741.
Reynaerts A,van de Wiele H,de Sutter G,Janssens J.1993.Engineered genes for fertility control and their application in hybrid seed production.Scientia Horticulturae,55:125-139.
Yang J H,Liu X Y,Yang X D,Zhang M F.2010.Mitochondrially-targeted expression of a cytoplasmic male sterility-associatedorf220gene causes male sterility inBrassica juncea.BMC Plant Biology,10:231.
Zhang H J,Wang H Y,Shi Y J,Zhang Y M,Yue J X,Wu S J,Zhu Y H,Liu Y L,Yang H Y.2007.Cotton genetic tranformation ofBarnasemale sterility gene.Cotton Sci,19(4):261-266.
