三種豆科植物光合特征初步研究
2013-02-24 10:00:06瞿思民吳夏華瞿巧文尹瑞安
中國林副特產 2013年1期
關鍵詞:植物
瞿思民,吳夏華,瞿巧文,尹瑞安
(漸江省慶元縣林業局,浙江慶元 323800)
山核桃(CaryacathayensisSarg.為我國特有的高檔干果和木本油料植物,20世紀80年代隨著人們生活水平的提高及對保健休閑產品需求的增加,農民種植山核桃積極性高漲,每年新增面積約3萬畝。山核桃栽培中,農民為采收方便,大量使用除草劑去除林地雜草,從而造成山核桃林生物多樣性下降、病蟲害加劇、水土流失等生態問題。
隨著人們生態意識的增強和山核桃產業的的可持續發展,如何在保持山核桃經濟效益的基礎上,實現生態效益和社會效益的最大化,是擺在當地政府和林農面前的一個重大問題。林草復合栽培是農業結構調整、改變單一種植結構、提高單位面積產出量及生態經濟效益的有效途徑[7-9]。近幾年山核桃產區林業技術部門對林下生草栽培進行了一系列的探索,如不同生草的選擇、栽培模式及技術等,選擇的草種包括白三葉(TrifoliumrepensLinn.)、紫云英(AstragalussinicusL.)和紫花苜蓿(MedicagosativaL.),黑麥草(Loliumperenne)、油菜(Brassica chinensisL.)等。本文對栽植于山核桃林下的三種豆科植物的光合生理進行初步研究,以期為山核桃林下生草栽培提供理論基礎。
1 材料與方法
1.1 研究地概況
實驗地設在浙江省臨安市浙江農林大學東湖校區苗圃,地理位置為E 118°51′,N 29°56′,該區屬亞熱帶季風氣候,年平均氣溫16℃,極端最高氣溫41.7℃,極端最低氣溫-13.3℃,年有效積溫5774℃,年降水量1350~1500 mm,年日照時數1774 h,無霜期235 d。海拔高度在90 m,土壤為發育于凝灰巖的紅壤土類。
1.2 試驗材料
3種豆科植物分別是白三葉、紫云英和紫花苜蓿,均為種子繁殖,于2010年9月種植于山核桃林下,常規管理。
1.3 氣體交換參數測定
于2011年5月25日上午9:30~11:00時,對3種豆科植物進行光合特性測定。測為生長完好、無病蟲害、葉面積相近的完全展開葉。光響應曲線利用Li-6400(美國,LI-Cor公司)紅藍光源6400-02測定,測量時的葉室溫度設置為25℃,CO2濃度370 μmol/L,光照強度梯度設置1500、1000、800、500、300、100、50、0μmol/m2·s,測定的的指標有光合速率(Pn 蒸騰速率(Tr 胞間CO2濃度(Gs 氣孔導度(Gs)等。每種植物測定6片葉片。
1.4 數據分析
采用Excel、DPS等數據分析軟件進行數據分析。
2 結果與分析
2.1 三種豆科植物光合速率的差異
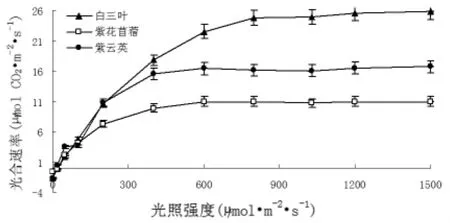
圖1 不同豆科植物的光飽和曲線
光響應曲線反映了植物光合速率隨光照強度的變化而變化的規律[8]。從圖l可以看出,隨著光量子通量的增大,凈光合速率幾乎呈直線上升;當光量通量達到一定值后,凈光合速率增加的幅度就逐漸減慢,最后達到一定限度,不再隨光量子通量的增加而增加,即達到光飽和。圖1表明,在相同條件下,白三葉的光合速率最大。通過光響應曲線可以計算出光補償點、光飽和點及最大凈光合速率,計算結果見表1。由于植物在光補償點時不能積累干物質,因此,光補償點的高低可以作為判斷植物在低光照強度條件下能否生長的標志。光補償點越小表明植物利用弱光的能力越強,紫云英、紫花苜蓿、白三葉光補償點分別為12.20、15.14、24.16μmol/m2·s。植物在光飽和點時光合速率最大,因此,光飽和點反映了植物利用強光的能力,越高說明植物在受到強光時不易發生抑制,植物的耐陽性越強,紫云英、紫花苜蓿、白三葉樹光飽和點分別為510.67、512.35、758.00μmol/m2·s,最大凈光合速率為15.55、10.70、23.58μmol CO2/m2·s。
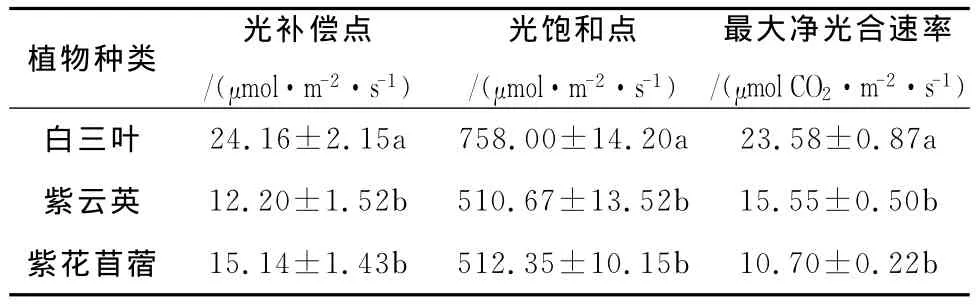
表1 三種豆科植物光補償點、光飽和點和光飽和點時凈光合速率
2.2 三種豆科植物氣孔導度的差異
從圖2中可知,在光合過程初期,氣孔導度隨光強的增大而增大,光強為200μmol/m2·s時,三種植物的氣孔導度均達最大值,隨著光強的繼續增大,氣孔導度則逐漸減小。從圖中還可看出。在整個光合過程中白三葉的氣孔導度明顯大于紫云英和紫花苜蓿。
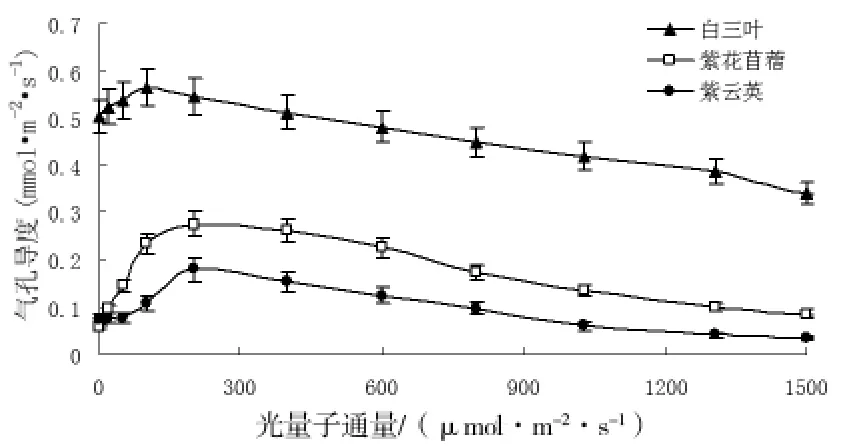
圖2 不同豆科植物的氣孔導度對光強的響應
2.3 三種豆科植物胞間二氧化碳濃度的差異
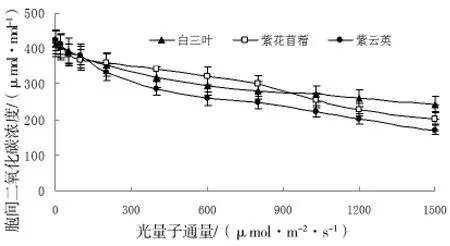
圖3 胞間二氧化碳濃度對光強的響應
圖3結果顯示,三種植物胞間二氧化碳濃度均著隨光強的增大而減小,植物間的差異并不明顯。
2.4 三種豆科植物蒸騰速率的差異
三種豆科植物蒸騰速率的變化見圖4。從圖中可知,光量子通量密度為0~50μmol/m2·s時,白三葉、紫云英、紫花苜蓿的蒸騰速率均隨光強的增大而增大,而后分別保持5.4、1.5、2.4mmol/m2·s左右。在整個光合過程中,白三葉的蒸騰速率明顯大于其它兩種植物。
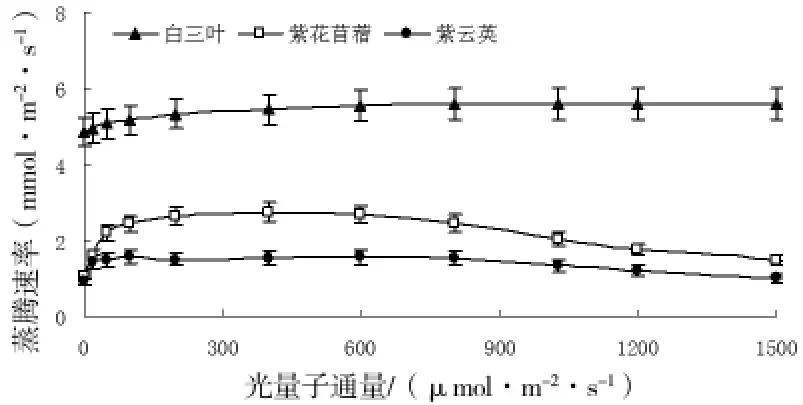
圖4 蒸騰速率對光強的響應
3 結論
通過對三種豆科植物氣體交換參數的測定,結果表明,白三葉、紫云英、紫花苜蓿的光補償分別為24.16、12.20、15.14μmol/m2·s,光飽和點分別為758.00,510.67,512.35μmol/m2·s,最大凈光合速率為23.58 15.55 10.70μmol CO2m s在整個光合過程中白三葉的氣孔導度、騰速率明顯大于紫云英和紫花苜蓿,而胞間二氧化碳濃度在植物間的差異并不明顯。白三葉光合能力明顯高于其它兩種植物,僅從光合特征進行分析,山核桃林下以栽培白三葉為好,從而可以產生較大的生物量。
[1] 王冀平,李亞南,馬建偉.山核桃仁中主要營養成分的研究[J].食品科學,1998,194:44-46.
[2] 王延平,張海,劉生禹,等.黃土區杏—苜蓿復合系統的土壤水分環境及效益研究[J].中國水土保持科學,2004,2(1):74-78.
[3] 王齊瑞,樊巍,譚曉風.杏—紫花苜蓿生草栽培系統根系分布及生長動態[J].林業科學,2008,44(8):141-144.
[4] 楊英,宋繼學,楊繼民,等.論苜蓿在農牧業和秀美山川中的作用[J].西安聯合大學學報,2001,4(13):97-100.
[5] 樊巍,孔令省,陰三軍,等.干旱丘陵區蘋果—紫花苜蓿復合系統對蘋果生長、產量和品質的影響[J].河南農業大學學報,2004,38(4):423-426,440.
[6] 劉芳,田春麗,劉世亮,等.有機酸—磷配施對紫花苜蓿生長及磷吸收的影響[J].河南農業大學學報,2007,41(4):382-386.
[7] 李文華,賴世登.中國農林復合經營[M].北京:科學出版社,1994.
[8] 陳玉香,周道瑋,張玉芬.玉米、苜蓿間作的產草量及光合作用[J].草地學報,2004,12(2):107-112.
[9] 張久海,安樹青,李國旗,等.林牧復合生態系統研究評述[J].中國草地,1999(4):52-60.
[10] 王建麗,張永亮.雜花苜蓿光合生理生態特性[J].草地學報,2006,14(2):1382142.
[11] 萬素梅,胡建宏,胡守林,等.不同紫花苜蓿品種水分利用效率研究[J].西北農業學報,2004,20(4):133-137.
[12] Vaughn D L,Viands D R.Nut ritive value and forage yield of alfalfa synthetics under three harvestmanagement systems[J].Crop Sci.1990,30:69 9-703.
[13] Olssont,Leverenz JW.Nonuniform stomatal closure and the apparent convexity of the photosynthetic photon flux density response curve[J].Plant,Cell and Environment,1994,17:701-710.
[14] Bhagsari A S,Brown R H.Leaf Photosynthesis and It s Correlation with Leaf Area[J].Crop Sci.1986,26:1272132.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
