利用EROD和GST酶評估竹子制漿造紙廢水的生物毒性
2013-01-05 02:10:22侯麗萍應光國趙建亮
中國造紙學報 2013年1期
關鍵詞:生物
侯麗萍 應光國 舒 琥 趙建亮
(1.廣州大學生命科學學院,廣東廣州,510006;2.中國科學院廣州地球化學研究所有機地球化學國家重點實驗室,廣東廣州,510640;3.廣州大學華南生物多樣性研究所,廣東廣州,510006)
造紙廢水含有大量有毒的有機污染物,對造紙廢水進行生物毒性測試可補充化學分析方法的不足,有效評價造紙廢水的安全性。目前,一般采用急性毒性測試方法[1]及基于細胞培養(yǎng)的生物標記法來評估造紙廢水的綜合毒性[2]。
食蚊魚(Gambusia affinis)屬鳉形目(Cyprinodontiformes)、胎鳉科(Poeciliidae),具有廣泛的生境適應性、繁殖周期短、生殖率高、明顯的兩性性狀等特點,且易于在實驗室飼養(yǎng),被廣泛用作造紙廢水毒性的指示生物物種[3]。采用酶活性指標監(jiān)測造紙廢水污染程度尚處起步階段,且尚未有利用野生食蚊魚活體 7-乙氧基-3-異吩嗆哇酮-脫乙基酶(7-ethoxyresorufino-deethylase,EROD)和谷胱甘肽硫轉移酶(Glutathione-S-transferase,GST)來評估造紙廢水毒性的報道。EROD是混合功能氧化酶系統(tǒng)(MFO)的主要組分,屬細胞色素氧化酶p450系中同工酶CYP1A1的一族,為第一階段解毒酶,主要通過催化氧化反應增加底物的極性。在正常水體環(huán)境中,魚體內的EROD活性相對較低,但在某些特定的外來化學物質(特別是各種多環(huán)芳香烴類化合物)的誘導下,魚體內的EROD活性異常升高。因此,可將魚體內EROD的活性作為水體環(huán)境中特定污染物的監(jiān)測指標。GST廣泛存在于動物肝臟微粒體中,是外源性物質在體內生物轉化的關鍵酶類,也是內源性物質代謝的重要酶。GST屬于第二階段解毒酶,主要催化內源性谷朧甘肽與底物進行結合反應,從而提高底物的水溶性,有利于底物從生物體內排出。
侯麗萍等[4]研究發(fā)現,廣東省四會市受竹子制漿造紙廢水污染的鄧村河道中食蚊魚的種群、個體和組織都發(fā)生了改變,并出現了雄性化的雌性食蚊魚,表明該區(qū)域河流可能受到竹子制漿造紙廢水中有毒有機物的污染,對當地生態(tài)環(huán)境和人們的健康造成嚴重的潛在影響。因此,本實驗測定了鄧村河道中野生食蚊魚活體EROD和GST的活性,以研究竹子制漿造紙廢水對魚類生物轉化酶系的損害,并深入評估該區(qū)域造紙廢水的生物毒性。
1 實驗
1.1 造紙作坊與采樣點
調查區(qū)域為受竹子制漿造紙廢水污染的廣東省四會市鄧村河(見圖1)。調查區(qū)域段的鄧村河沿岸有7家造紙作坊(造紙作坊M1與M2、M3與M4、M4與M5均相距約400 m,M2與M3、M5與M6、M6與M7分別相距約800、500、300 m),均以竹子為原料,采用“小蘇打浸泡法”制漿,每個作坊的紙產品產量約為5 t/a。造紙廢水主要為碎竹和洗漿過程中產生的廢水,廢水只經過濾處理后直接排入鄧村河,無其他處理措施,排放量約為40 m3/d。
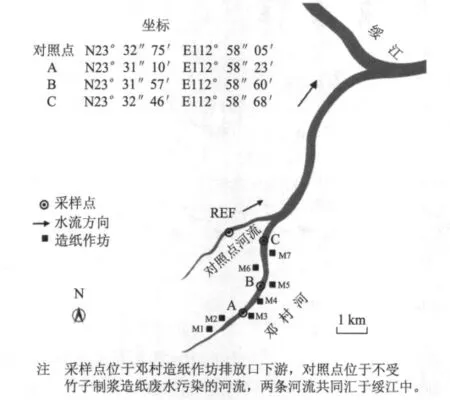
圖1 采樣點與對照點示意圖
1.2 食蚊魚的采集及預處理
食蚊魚采集時間為2009年8月和2010年3月。在各采樣點,用大型漁網捕獲足夠數量的食蚊魚,放入已編號的大型塑料桶中,每個塑料桶盛裝10 L采樣點的原水,用氧氣泵充氧,并在24 h內運回實驗室,48 h內進行雌雄分類、解剖及性體指標測定。EROD活性受魚體內激素的影響較大,不同生長發(fā)育階段的雌性食蚊魚的EROD活性具有顯著差異。因此,在測定EROD活性時,把食蚊魚分為成熟雌性食蚊魚、未成熟雌性食蚊魚及成熟雄性食蚊魚,而測定GST活性時只分為雌性食蚊魚和雄性食蚊魚。根據Noggle[5]的方法,定義具有生殖突起的為雌性食蚊魚,無生殖突起的為雄性食蚊魚;交接器具有鉤狀結構的雄性食蚊魚為成熟的雄性食蚊魚,不帶鉤狀結構的為未成熟的雄性食蚊魚;體長>20 mm的雌性食蚊魚為成熟的雌性食蚊魚,體長<20 mm的雌性食蚊魚為未成熟的雌性食蚊魚;具有明顯胎斑的為懷孕的食蚊魚。分類后,挑選大小基本一致的食蚊魚進行解剖,保證每個采樣點制備酶源的食蚊魚的數量不少于30條。采集食蚊魚的同時,測定各采樣點的水質參數,如pH值、溫度、硬度、溶氧量、TSS及BOD7。
1.3 酶源的制備
將食蚊魚于冰浴條件下解剖后,迅速取出魚肝組織,將用濾紙吸去血漬的魚肝置于組織勻漿器中。按以下步驟配置緩沖液 PBS1:加入11.184 g KCl和5.206 g HEPES(4-羥乙基哌嗪乙磺酸、N-(2-羥乙基)哌嗪-N'-2-乙烷磺酸)于1 L水中,調節(jié)pH值至7.5。加入1 mL該緩沖液于組織勻漿器中勻漿1 min。然后取適量勻漿液于高速冷凍離心機中(Beckman,4℃,10000 r/min)離心20 min。取上清液并分為3份,每份200 μL,在4℃下保存。其中,2份分別用于EROD和GST活性測定,1份用于酶源中蛋白質含量的測定。
1.4 EROD活性的測定
采用改進的快速終止熒光光度法測定食蚊魚的EROD活性,即根據鯽魚肝臟EROD活性的測定方法略作修改[6]。反應體系包含190 μL磷酸緩沖液(濃度 0.1 mol/L,pH 值 7.8),30 μL 7-乙氧基-3-異吩噁唑酮(ERF,濃度 1.73 μmol/L),50 μL 酶源上清液。向測試管中加入30 μL四鈉鹽(NADPH,濃度0.4 mol/L),并在 20℃水浴中反應 10 min,加入2 mL預冷的甲醇終止反應。空白管用重蒸水代替酶源上清液。用熒光分光光度計測定產物7-羥基-3-異吩噁唑酮(RF)的熒光強度,激發(fā)波長為550 nm,發(fā)射波長為585 nm。參照標準曲線得到RF的含量,并計算EROD活性。
1.5 GST活性的測定
參照Habig等[7]的方法測定食蚊魚的GST活性。用購自南京建成生物工程研究所的試劑盒進行測定,酶促反應為1200 μL的反應體系。向1.5 mL的離心管中加入150 μL的基質液,50 μL酶源上清液,對照管不加酶液,在37℃下水浴10 min,再加入500 μL試劑(取試劑盒中的粉末加入170 mL 100℃的蒸餾水,充分溶解,取出試劑盒中50 mL的溶液,充分混合這兩種液體,即成過飽和溶液,該飽和溶液的上清液即為實驗用的試劑),500 μL無水乙醇,對照管另加入50 μL酶源上清液,大力振蕩搖勻后,在4000 r/min下常溫離心10 min,取上清液進行顯色反應。顯色反應在1.5 mL的EPA管中進行,反應總體系為1125 μL。顯色反應液混勻后在室溫下放置15 min,取300 μL顯色后的反應液于96孔板中進行412 nm吸光值的測試。
1.6 酶源中蛋白質含量的測定
采用酶標儀按Bradford的方法[8]測定酶源中蛋白質的含量,蛋白標準液為牛血清蛋白(BSA)。取適量離心后的酶源上清液或其稀釋液,加入顯色劑考馬斯亮藍(Coomassie brilliant blue G250),當顯色劑與蛋白質結合后,會由紅色變?yōu)樗{色。測定前先按V(考馬斯亮藍)∶V(水)=1∶4稀釋考馬斯亮藍,在96孔板中加入300 μL稀釋后的考馬斯亮藍,然后加入10 μL酶源上清液,并設3個孔加入蛋白標準液,另外3個孔為空白樣。待反應結束后,通過測定595 nm下的吸光度來確定酶源中蛋白質的含量。
1.7 統(tǒng)計分析
實驗所得數據均用SPSS(ver.11.5)進行分析;用單因素方差分析(ANOVA)檢驗不同采樣點食蚊魚的EROD和GST活性,并用Dunnett's test對結果進行驗后多重比較。如果P<0.05,認為有顯著性統(tǒng)計意義。
2 結果與討論
2.1 各采樣點的水質參數
各采樣點的水質參數如表1所示。由表1可知,采樣點A、B、C處的水溫、pH值與對照點差異不大。與對照點的水質相比,采樣點水質的電導率稍高,溶氧量低,BOD7和TSS含量高。這表明,鄧村竹子制漿造紙廢水的排放對該區(qū)域水體環(huán)境造成了污染。
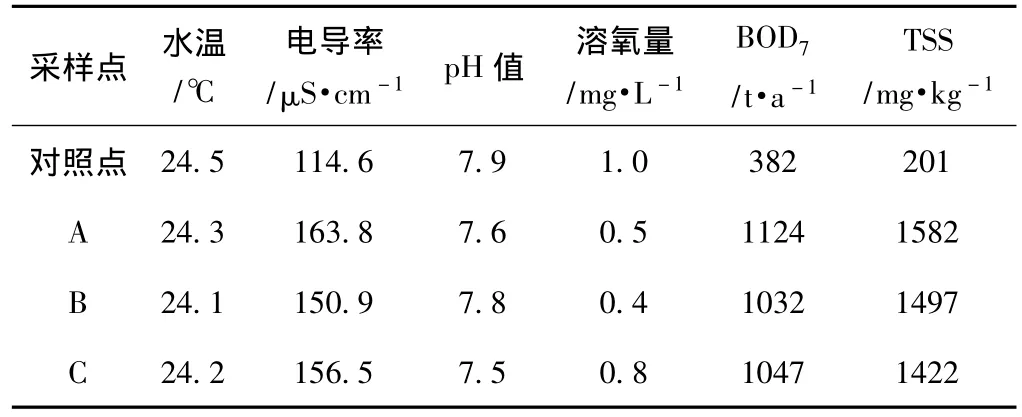
表1 采樣點的水質參數
2.2 造紙廢水對食蚊魚EROD活性的影響
圖2所示的是2009年夏季(8月)各個采樣點和對照點的食蚊魚EROD活性。由圖2可知,與對照點食蚊魚的EROD活性相比,各采樣點食蚊魚的EROD活性顯著高(P<0.01)。對照點未成熟雌性食蚊魚、成熟雌性食蚊魚、成熟雄性食蚊魚的EROD 活 性 分 別 為 2.62 ×10-12、1.97 ×10-12、2.08×10-12mol/(min·mg)。其中,采樣點 A處的未成熟雌性食蚊魚、成熟雌性食蚊魚、成熟雄性食蚊魚的EROD活性分別為對照點的4.1、4.0和3.5倍;采樣點B處的未成熟雌性食蚊魚、成熟雌性食蚊魚、成熟雄性食蚊魚的EROD活性分別為對照點的3.2、3.4、3.5倍;采樣點C處的未成熟雌性食蚊魚、成熟雌性食蚊魚、成熟雄性食蚊魚的EROD活性分別為對照點的3.2、3.4、3.1倍。各采樣點的未成熟雌性食蚊魚的EROD活性比成熟雌性食蚊魚和成熟雄性食蚊魚的高。
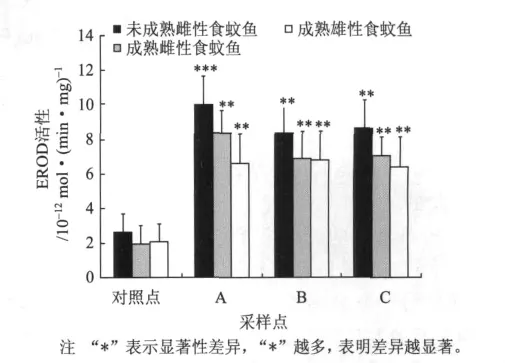
圖2 2009年8月份鄧村造紙廢水對食蚊魚EROD活性的影響
圖3所示的是2010年春季(3月)各采樣點和對照點食蚊魚EROD活性。由圖3可知,對照點未成熟雌性食蚊魚、成熟雌性食蚊魚、成熟雄性食蚊魚的EROD 活 性 分 別 為 1.91 × 10-12、1.34 × 10-12、1.08×10-12mol/(min·mg)。與對照點相比,各采樣點食蚊魚的EROD活性顯著高(P<0.01)。其中,采樣點A處的未成熟雌性食蚊魚、成熟雌性食蚊魚、成熟雄性食蚊魚的EROD活性分別為對照點的4.6、4.4、2.2倍;B點未成熟雌性食蚊魚、成熟雌性食蚊魚、成熟雄性食蚊魚的EROD活性分別為對照點的3.8、4.0、2.3倍;C點未成熟雌性食蚊魚、成熟雌性食蚊魚、成熟雄性食蚊魚的EROD活性分別為對照點的3.2、3.8、2.3倍。各采樣點的未成熟雌性食蚊魚的EROD活性比成熟雌性食蚊魚和成熟雄性食蚊魚高。
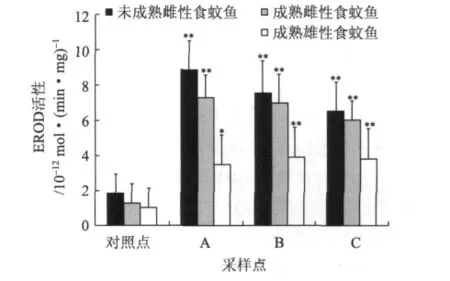
圖3 2010年3月份鄧村造紙廢水對食蚊魚EROD活性的影響
Andersson等[9]研究表明,與未受造紙廢水污染的河流中的魚類相比,生活在受含漂白工段造紙廢水污染的河流中魚類的EROD活性顯著升高,其活性與離造紙廠的距離成反比。目前,由于生產工藝的改良,漂白工段廢水中的許多對環(huán)境有害的化合物已被去除,如多氯聯(lián)苯(PCB)和多環(huán)芳烴(PAH)等。但Martel等[10]研究證實,不含漂白工段的造紙廢水仍會誘導魚類的EROD活性升高。他們認為,造紙廢水中誘導魚類EROD活性升高的化合物不是高分子質量的疏水性化合物,而是來源于造紙原材料中的二級PAH形式的疏水化合物——樹脂、緇醇類等物質。本研究結果顯示,無論是夏季還是春季,生活于受鄧村竹子制漿造紙廢水污染的河道中的食蚊魚EROD活性均比對照點的高,說明該造紙廢水中的某些化合物能與細胞內芳烴受體(Ah2R)結合,誘導了EROD活性升高。鄧村造紙作坊采用的原料是竹子,因此,很有可能在生產過程中釋放出來的緇醇類物質在影響魚類生長發(fā)育的同時,誘發(fā)EROD活性升高,引發(fā)毒性效應。因此,除了對常規(guī)的PCB和PAH化合物進行監(jiān)測外,由二級PAH形式的疏水化合物引起的環(huán)境污染問題也需引起一定的重視。
Elskus等[11]研究表明,雌魚在繁殖期間生物轉化系統(tǒng)受到抑制,EROD活性也受到抑制。因此,處于繁殖期間的雌魚的EROD活性比其他魚的低。Bankey等[12]研究表明,造紙廢水均能引起未成熟和成熟黑鱸的EROD活性升高,未成熟黑鱸的EROD活性均比成熟并處于繁殖期間黑鱸的EROD活性高。本實驗研究結果顯示,各采樣點未成熟雌性食蚊魚EROD活性比其他發(fā)育階段食蚊魚的EROD活性高,這與Bankey等的研究結果一致。
EROD在內分泌系統(tǒng)中扮演何種角色及其對繁殖造成何種潛在的影響,機理如何,這些都需要進一步研究。EROD的活性與水體環(huán)境的溫度、魚的種類、性別、生長發(fā)育階以及肝臟的健康情況密切相關,用其作為生物標志物的時候一定要注意以上條件的差異性。
2.3 造紙廢水對食蚊魚GST活性的影響
圖4顯示的是2009年夏季(8月)鄧村竹子制漿造紙廢水對食蚊魚GST活性的影響。由圖4可知,與對照組雌性食蚊魚的GST活性相比,采樣點A、B處雌性食蚊魚的GST活性顯著降低(P<0.05)。除了采樣點A外,采樣點B、C處的雄性食蚊魚的GST活性均比對照點的低,但無顯著差異(P>0.05)。
圖5顯示的是2010年春季(3月)鄧村竹子制漿造紙廢水對食蚊魚GST活性的影響。由圖5可知,與對照點食蚊魚GST活性相比,采樣點A、B、C處食蚊魚的GST活性總體低,采樣點B處的雌性食蚊魚及C處的雌、雄食蚊魚的GST活性較對照點的有顯著差異(P<0.05)。
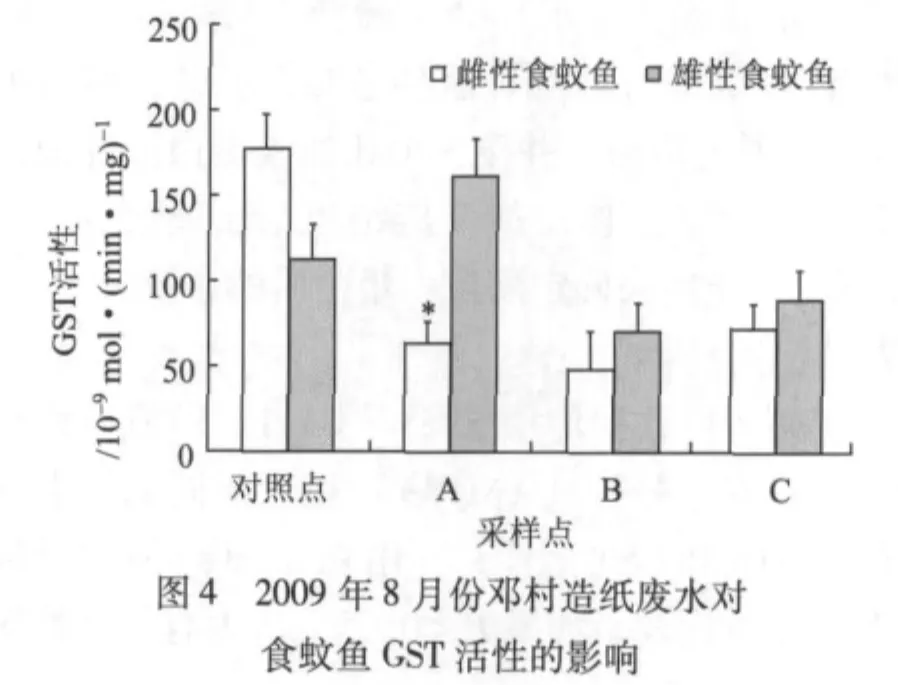
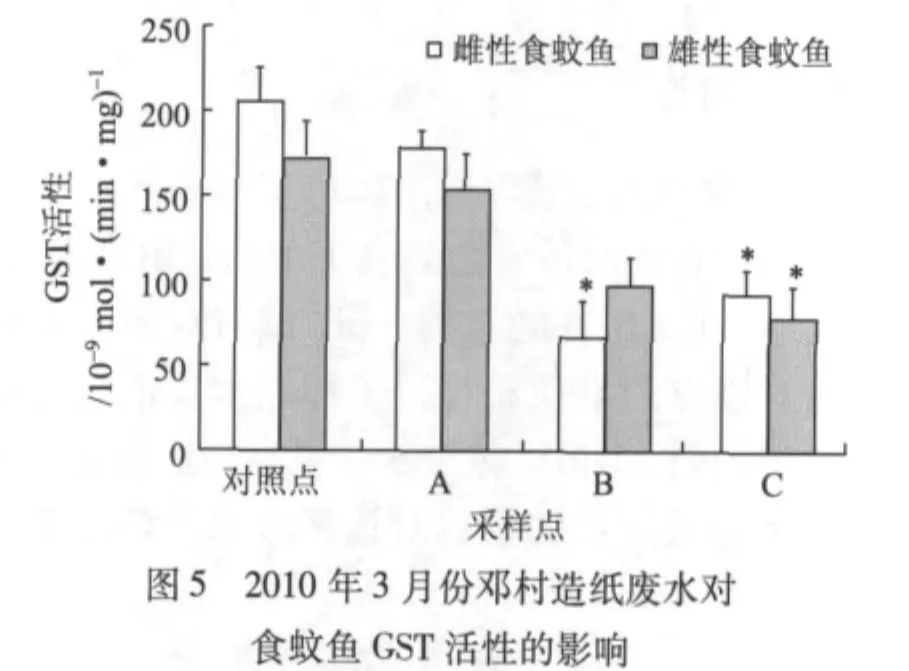
GST是生物機體內重要的代謝酶系之一。該酶存在于生物機體的各種組織中,具有消除體內自由基和解毒的雙重功能,其活性的大小可反映生物機體抗氧化能力的高低,并對生物機體肝臟的早期損傷診斷具有一定的價值[13]。尹大強[14]研究證實,含漂白工段的造紙廢水會顯著誘導鯽魚肝臟微粒體GST的活性。Holth T F等[15]用造紙廢水暴露斑馬魚,研究結果表明,短時間暴露對斑馬魚GST活性起誘導作用,長時間暴露則起抑制作用。王重剛[16]的研究也發(fā)現,用2,4-二氯苯酚、苯并芘(BaP)、重金屬等污染物暴露魚類后,在短時間和低濃度劑量暴露時,其肝臟和脾臟的谷胱甘肽過氧化物酶(GPx)和GST活性顯著升高;在長時間和高濃度劑量暴露后,由于魚體內谷胱甘肽(GSH)含量降低和過氧化氫酶(CAT)活性增大,則污染物對GPx和GST活性的誘導作用消失,甚至抑制它們的活性。本研究結果顯示,各采樣點食蚊魚的GST活性總體比對照點的低,但只有個別采樣點出現顯著差異。GST活性低的原因可能是,受竹子制漿造紙廢水的污染程度比較嚴重,食蚊魚體內GSH消耗過大,造成GST活性下降。以上結果表明,GST用作生物標志物指示竹子制漿造紙廢水生物毒性時沒有EROD敏感。
3 結論
測定了生活于受竹子制漿造紙廢水(采用“小蘇打浸泡法”制漿)污染的河道中食蚊魚活體肝臟7-乙氧基-3-異吩嗆哇酮-脫乙基酶(7-ethoxyresorufinodeethylase,EROD)和谷胱甘肽硫轉移酶(Glutathione-S-transferase,GST)的活性。結果表明,食蚊魚可作為檢測竹子制漿造紙廢水污染的敏感生物標志物,其EROD活性可有效真實地評價竹子制漿造紙廢水的安全性,GST在用作竹子制漿造紙廢水生物毒性的標志物時,響應沒有EROD敏感。EROD的活性與水溫、魚的性別、成熟階段及肝臟的健康情況密切相關,用其作為生物標志物的時候一定要注意以上條件的差異性。
[1]沈盎綠,馬勝偉,平仙隱,等.制漿造紙廢水對不同魚類急性毒性研究[J].海洋漁業(yè),2006,28(4):332.
[2]倪瀟瀟,王子健.造紙廠廢水的生物毒性測試[J].中國造紙,2006,25(12):27.
[3]Cody R P,Bortone S A.Masculinization of mosquitofish as an indicator of exposure to kraft mill effluent[J].Bulletin of Environmental Contamination and Toxicology,1997,58(3):429.
[4]Hou Liping,Xie Yongping,Ying Guangguo,et al.Developmental and reproductive characteristics of western mosquitofish(Gambusia affinis)exposed to paper mill effluent in the Dengcun River,Sihui,South China[J].Aquatic Toxicology,2011,103(3/4):140.
[5]Noggle J J.Eastern Mosquitofish as Bioindicator of Pulp and Paper Effluents[D].Gainesville:University of Florida,2005.
[6]吳 鷹,孟艷玲,周忠良.環(huán)境內分泌干擾物對河川沙塘鱧的雌激素效應檢測[J].環(huán)境檢測管理與技術,2006,8(6):13.
[7]Habig W H,Pabst M J,Jakoby W B.Glutathione stransferases:The first enzymatic step in mercapturic acid formation[J].The Journal of Biological Chemistry,1974,249:7130.
[8]Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding[J].Anal Biochemistry,1976,72(1/2):248.
[9]Andersson T,Hardig J,Larsson A.Physiological disturbances in fish living in coastal water polluted with bleached kraft pulp mill effluent[J].Canadian Journal of Fisheries and Aquatic Sciences,1988,45(9):1525.
[10]Martel P H,Kovacs T G,O’Connor B I,et al.A survey of pulp and paper mill effluents for their potential to induce mixed function oxidase enzyme activity in fish[J].Water Research,1994,28(8):1835.
[11]Elskus A A,Stegeman J J,Susani L C,et al.Polychlorinated biphenyls concentration and cytochrome P-450E expression in winter flounder from contaminated environments[J].Marine Environmental Research,1989,28(1/4):25.
[12]Laura A Bankey,Peter A Van Veld,Dennis L Borton,et al.Responses of cytochrome P4501A in freshwater fish exposed to bleached kraft mill effluent in experimental stream channels[J].Canadian Journal of Fisheries and Aquatic Sciences,1995,52(3):439.
[13]George S G.Enzymology and molecular biology of phase xenobioticⅡ conjugating enzymes in fish[M].Washington D C:Lewis Publishers,1994.
[14]尹大強.漂白紙漿廢水對鯽魚(Carassius auratus)肝臟微粒體代謝酶的影響[J].南京大學學報:自然科學,2003,39(3):392.
[15]Holth T F,Nourizadeh-Lillabadi R,Blaesbjerg M.Differential gene expression and biomarkers in zebrafish(Danio rerio)following exposure to produced water components[J].Aquatic Toxicology,2008,4(11):277.
[16]王重剛.苯并(a)芘和芘的混合物暴露對梭魚脾臟抗氧化防御系統(tǒng)的影響[J].海洋學報,2003,25(2):135.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
