三峽水庫香溪河庫灣夏季藍藻水華成因研究
2012-12-25 02:19:58田澤斌劉德富楊正健方小鳳姚緒姣方麗娟三峽大學水利與環境學院三峽庫區生態環境教育部工程研究中心湖北宜昌443002湖北工業大學資源與環境學院湖北武漢430072武漢大學水利水電學院湖北武漢430072
中國環境科學 2012年11期
田澤斌,劉德富,2*,楊正健,3,方小鳳,姚緒姣,方麗娟 (.三峽大學水利與環境學院,三峽庫區生態環境教育部工程研究中心,湖北 宜昌 443002;2.湖北工業大學資源與環境學院,湖北 武漢 430072;3.武漢大學水利水電學院,湖北 武漢 430072)
三峽水庫香溪河庫灣夏季藍藻水華成因研究
田澤斌1,劉德富1,2*,楊正健1,3,方小鳳1,姚緒姣1,方麗娟1(1.三峽大學水利與環境學院,三峽庫區生態環境教育部工程研究中心,湖北 宜昌 443002;2.湖北工業大學資源與環境學院,湖北 武漢 430072;3.武漢大學水利水電學院,湖北 武漢 430072)
為探討香溪河夏季藍藻水華發生過程及主要影響因素,于2010年7月19日~8月30日在藍藻水華暴發區域開展持續監測,并對水華過程進行分析.結果表明,水華持續 25d,自 7月 23日暴發,8月 16日逐漸消退,藻密度最高達到 108.03×106cells/L,優勢藻種為魚腥藻(Anabaena);自身懸浮機制、固氮機制、能夠產生藻毒素抑制其他藻類生長,是魚腥藻成為優勢藻種的主要原因;充足的營養物質、顯著的水體分層是水華暴發的必要條件,水華期間藍藻對硝氮利用顯著;在具備充足的營養鹽的穩定水體中,水溫持續升高、混合層與真光層比值的降低是誘發藍藻水華的關鍵因子,并且在混合層與真光層比值為0.5時對藍藻增殖影響最大.
三峽水庫;香溪河庫灣;藍藻水華;混合層
三峽水庫成庫后,藻類水華發生的頻率和規模開始增加,優勢藻種從河流型向湖泊型轉變[1],尤其是2008年開始部分支流庫灣夏季暴發的藍藻水華不僅破壞了生態環境,而且威脅著人類健康及水源安全.目前,有關三峽水庫支流有害藍藻水華頻繁出現現象的研究較多[1-3],主要依托野外觀測,研究水華期間藻類對特定環境因子的響應關系,以及種群結構和演替特征等.李哲等[4]認為,溫度升高、真光層深度下降是誘發夏季藍藻水華的重要因子,但沒能建立復雜物理背景下藍藻水華與水溫的具體響應機制.張敏等[2]指出,庫灣相對穩定的水體環境是藍藻水華暴發的必要條件,但沒有給出香溪河庫灣具體的水體分層狀態.目前對三峽水庫支流藍藻水華的研究成果尚難以應用于闡釋深水河道型水庫藍藻水華形成機理.因此,本文以三峽水庫典型支流香溪河為例,對2010年7~8月香溪河庫灣水華過程進行持續監測,以該區域特殊水體分層背景下藍藻水華的生態響應機制為研究重點,圍繞水溫、水體穩定系數、特殊的水體層化模式、營養鹽等因子對藍藻水華的影響機理展開探討,探究藍藻形成優勢并暴發水華的誘導因子,以期為探討三峽水庫支流水華的機理提供參考,為有效防治支流水華提供理論依據.
1 研究區域及研究方法
1.1 研究區域及樣點設置
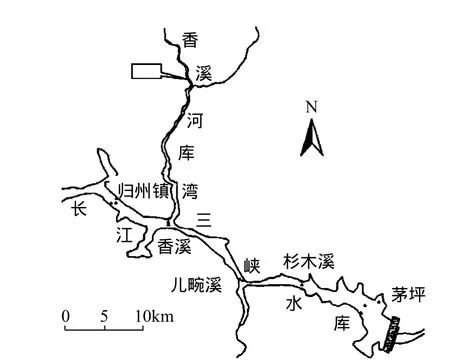
圖1 香溪河采樣點位置示意Fig.1 Sampling site in Xiangxi Bay
香溪河系長江三峽庫區湖北省境內最大的支流,發源于神農架林區,河流由北向南,在秭歸香溪匯入長江,其干流長度 94km,流域范圍(110°25′~111°06′)E,(30°57′~31°34′)N,流域面積3099km2,擁有南陽河、古夫河、高嵐河3條主要支流.地處亞熱帶大陸性季風氣候區,由于地勢高差大,地形復雜,氣溫垂直變化明顯.年平均氣溫為16.6℃,流域多年平均流量為40.18m3/s,年降雨量 1015.6mm.香溪河庫灣屬深水型回水庫灣,由于干支流的水溫差異及顆粒物濃度含量不同導致分層異重流現象長期存在,同時庫灣存在顯著的水溫分層現象[5].兩種密度相差不大、可以相混的流體,因為密度的差異而發生的相對運動稱為異重流[6].三峽水庫調度時,長江干流水體與香溪河庫灣水體通過異重流方式進行水體交換.依托三峽大學水生態與環境野外觀測站,2010年7月-8月在香溪河庫灣峽口附近選取香溪河水華暴發嚴重的區域設置監測點(圖1)進行監測.
1.2 監測頻率與指標
監測時間為2010年7~8月,每天上午10:00監測1次,主要監測指標有:葉綠素a濃度、藻種及其密度、水溫、透明度、流速、水下光強、營養鹽(N-N、PO43--P、SiO2-Si)等.在采樣點每次取350mL表層水,用GF/C濾膜過濾并10℃保存帶回實驗室分析,用90%的丙酮提取 24h,3500r/min離心 10min,收集上清液,用分光光度法[7]測定表層葉綠素a濃度.根據《水和廢水監測分析方法》[7]測定水中 NO3--N、PO43--P濃度,用硅鉬黃分光光度法[7]測定溶解性硅酸鹽濃度SiO2-Si;水溫由Hydrolab DS5多參儀現場測定;流速采用Vector三維點式流速儀(挪威)測定;現場同步采用手持式風速風向儀現場記錄觀測點風速風向;用PVC塑料瓶取1200mL水樣,加魯哥固定,帶回實驗室沉降濃縮成75 mL,在顯微鏡(10×40倍)下對浮游植物細胞進行計數,藻類根據《中國淡水藻類》[8]和《淡水微型生物圖譜》[9]進行鑒定.
1.3 數據處理
本文忽略水體中泥沙對水體密度的影響,水溫對應的水體密度按Lawson等[10]的公式(1)計算:
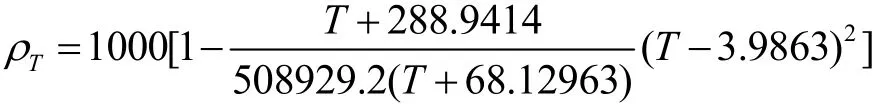
式中: ρT為水體密度,單位 kg/m3;T為水體溫度,單位℃.
水體穩定系數(RWCS)是一個無量綱的參數,該參數通過比較底部(Db)和表面(Ds)水的密度差異和純水在 4℃(D4)和 5℃(D5)時的密度差異,根據公式(2)計算[11]:
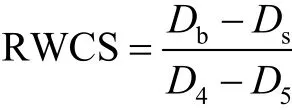
自然條件下的水體真光層深度的計算采用透明度的 2.7倍[12],混合層深度采用與表層水溫相差不大于0.2℃的深度范圍[13].
2 結果與分析
2.1 葉綠素a濃度及藻密度
研究表明[13],香溪河庫灣藍藻水華暴發時葉綠素a濃度的閥值為40mg/m3.由圖2可知,監測期間,香溪河庫灣葉綠素a濃度先增大后減小,變幅為0.47~87.48mg/m3.香溪河庫灣在7月23日~8月16日暴發了一次藍藻水華,持續25d.7月19日葉綠素a濃度為31.70mg/m3,隨后葉綠素a濃度波動明顯,總體呈增大趨勢,至8月5日達峰值87.48mg/m3,之后葉綠素 a濃度維持在較高水平波動,8月17日回落至7.23mg/m3,水華消退.此后葉綠素 a濃度出現短期小幅升降過程,并未形成藍藻水華,8月 25日以后葉綠素 a濃度降至2mg/m3以下.
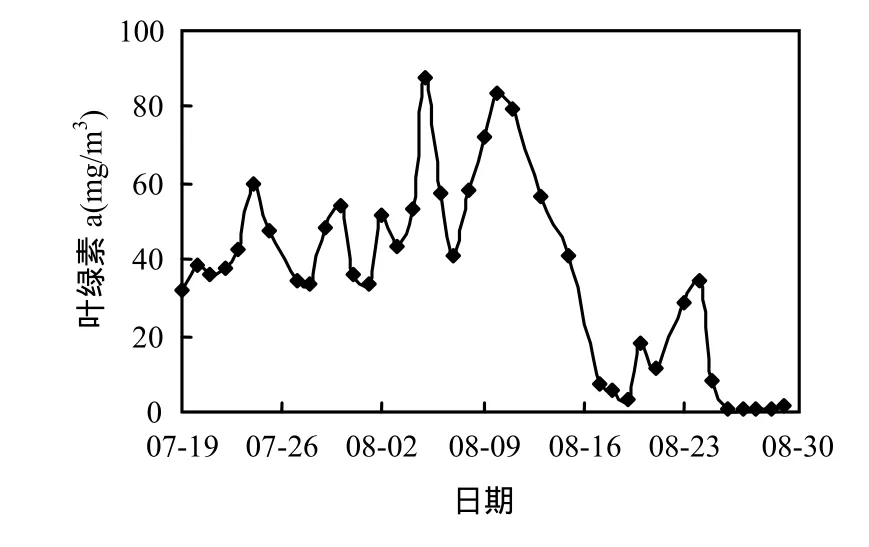
圖2 葉綠素a濃度隨時間的變化Fig.2 Variation of chlorophyll a concentration in Xiangxi Bay during the summer
藻細胞密度能很直觀的表明水體中藻類的多少和水華暴發強度[13],富營養化水體發生水華期間,藻類大量繁殖,藻密度不斷增加,當水體發生重度水華時,浮游植物的細胞密度一般大于20×106cells/L[13-14].從圖3可知,香溪河庫灣藻類總密度整體先增大后減小,與葉綠素 a濃度變化趨勢大體一致,鏡檢得出優勢藻種為魚腥藻(Anabaena).7月19日藻密度為29.46×106cells/L,之后呈上升趨勢,7月 28日達峰值 138.54×106cells/L后,呈現先下降后上升的過程,8月5日藻密度增至127.84×106cells/L,此后逐漸回落,8月25日降至0.48×106cells/L后維持較低水平.監測期間,藍藻密度占總密度的百分比最高達94%,與總密度變化趨勢基本一致.
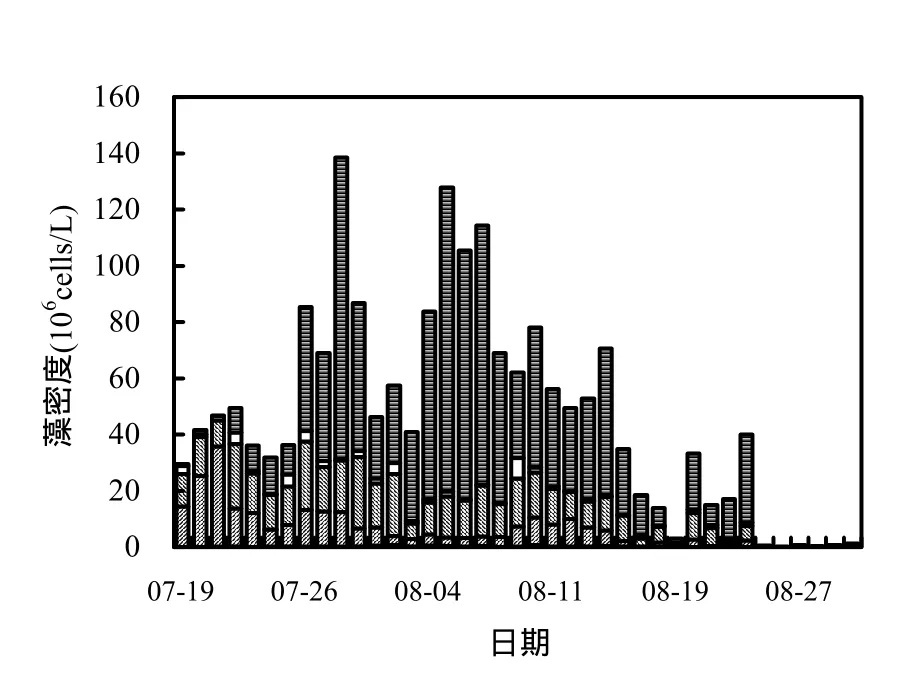
圖3 藻密度及藻種組成隨時間的變化Fig.3 Variation of cell density and composition in Xiangxi Bay during the summer
2.2 水溫
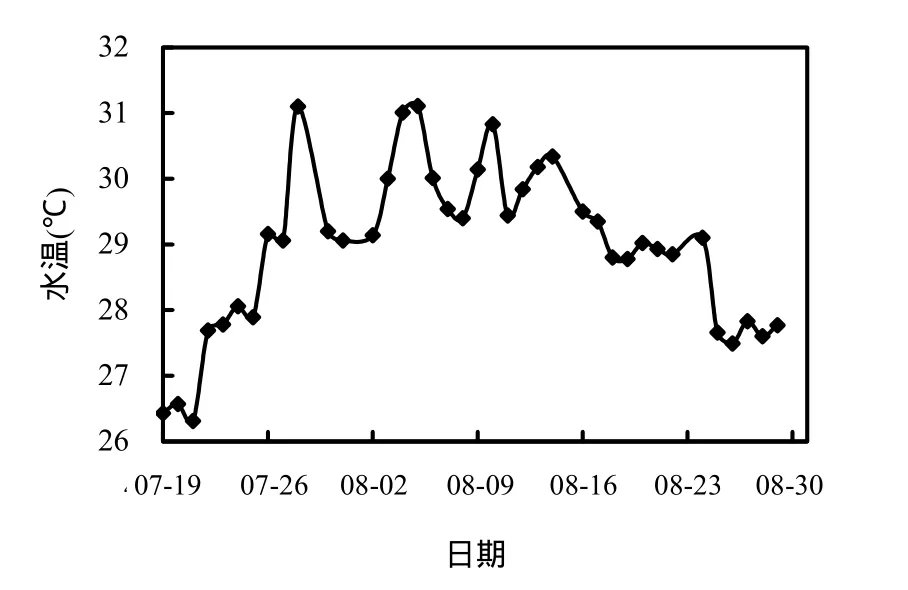
圖4 表層水溫隨時間的變化Fig.4 Variation of surface temperature in Xiangxi Bay during the summer
水溫是藻類生長的重要影響因子.它通過控制光合作用、酶促反應或呼吸作用強度,直接影響藻類細胞生長增殖相應的一系列過程[4,15],同時適宜的水溫是藍藻暴發的必要條件[16].由圖 4知,香溪河庫灣表層水溫先增大后減小,在26.31~31.11℃之間,符合多數藍藻生長增殖最適溫度(25~35℃[17]).監測期間,由于受氣溫降低及降雨的影響,7月19日庫灣表層水溫較低,隨著太陽輻射及氣溫的顯著升高,表層水溫升溫較快,7月28日達峰值31.11℃,此后在 30℃上下波動,維持藍藻生長所需的較高溫度,8月16日后氣溫逐漸降低,降雨較為頻繁,加之來流量明顯增大,加快了水體交換,表層水溫呈降低的趨勢,至 8月 25日表層水溫降至27.66℃,此后呈變幅較小的波動趨勢.
2.3 水體穩定系數
水體穩定系數是評價水體垂向混合程度及水體分層狀態的重要參數,水體穩定系數越大,水體垂向越穩定,分層現象越顯著;水體穩定系數越小,越易形成混合水體[18].水體的穩定性是影響藍藻生長及分布的重要物理因素,垂直穩定分層越明顯的水體,越易暴發高強度的藻類水華[19].
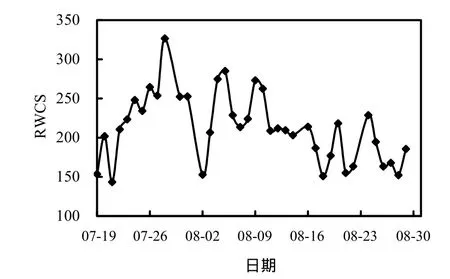
圖5 水體穩定系數隨時間的變化Fig.5 Variation of RWCS in Xiangxi Bay during the summer
由圖5知,監測期間香溪河庫灣水體穩定系數先增大后減小,在 143.40~326.66范圍變化,與表層水溫(圖4)變化趨勢基本一致.由于表層水溫顯著升高,表底溫差增大,水溫分層現象明顯,之后隨著氣溫的降低,加之降雨頻繁,水體垂向摻混劇烈,表底溫差逐漸減小,分層特性減弱,從而使水體穩定系數降低,峰值出現在 7月 28日,達326.66.監測期間水體穩定系數較高,熱分層現象明顯,藍藻可以通過控制偽空胞的數量來調控細胞密度,利用其懸浮機制在獲取更多的營養和光照競爭中取得優勢[20],而水體的分層加速了藍藻這種喜好在穩定水體中生長的種類大量的繁殖和聚集,致使藍藻在水體中更具有優勢[21].
2.4 真光層、混合層及其比值
真光層是浮游植物接受有效光照進行光合作用的最大深度[22],它直接影響水柱中藻類的生長增殖過程.由圖6知,香溪河庫灣真光層深度變幅較小,變化范圍在 0.8~3.4m之間.混合層是藻類可以遷移懸浮的水層范圍,香溪河庫灣混合層深度呈先降低后升高趨勢,在 0.8~6m范圍內變化.8月中下旬開始,混合層深度顯著增加,表層水體呈混合狀態.在研究中,可以采用混合層和真光層的比值(Zmix/Zeu)來反映藻類在真光層內生長的潛勢[23].由圖 6知,監測期間 Zmix/Zeu與藻密度變化(圖3)趨勢相反,在0.3~3.1之間變化,7月19日之后Zmix/Zeu明顯減低,隨著Zmix/Zeu的降低,真光層范圍內的藻類能夠利用充足的光照,提高增長率,表現為藻密度相應增加,8月16以前Zmix/Zeu隨時間變幅較小,相應藻密度維持較高水平,此后Zmix/Zeu總體呈增加趨勢,混合層深度明顯大于真光層深度,表現為混合層內的藻類受到光照的限制,生長率降低,從而對藻類增殖起到抑制作用.
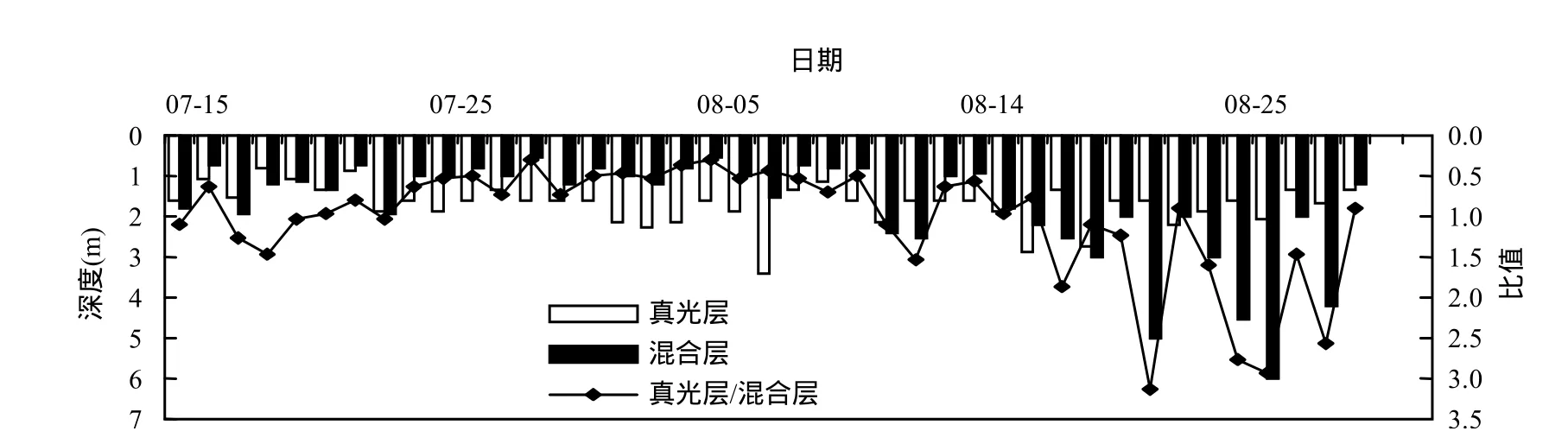
圖6 真光層、混合層深度及其比值隨時間變化Fig.6 Variations of Zeu, Zmix and Zmix/Zeu in Xiangxi Bay during the summer
2.5 營養鹽
溶解性營養鹽可以被浮游植物直接吸收利用進行自身生長增殖.香溪河庫灣氮磷營養含量豐富[24],對藍藻水華的暴發不存在絕對的營養鹽限制.
由圖7可知,監測期間香溪河庫灣NO3--N濃度大致呈先降低后增加的趨勢,結合圖2可知,藍藻水華期間NO3--N濃度顯著降低,水華消退后NO3--N濃度恢復到較高水平,說明藍藻的增殖需要消耗大量的無機氮營養,同時由于反硝化作用也會消耗大量的氮營養.監測期間正磷酸鹽PO43--P濃度整體呈波動降低的趨勢,變幅較大.SiO2-Si濃度在監測期間變化幅度不大,說明藍藻增殖期間對硅營養消耗較少.
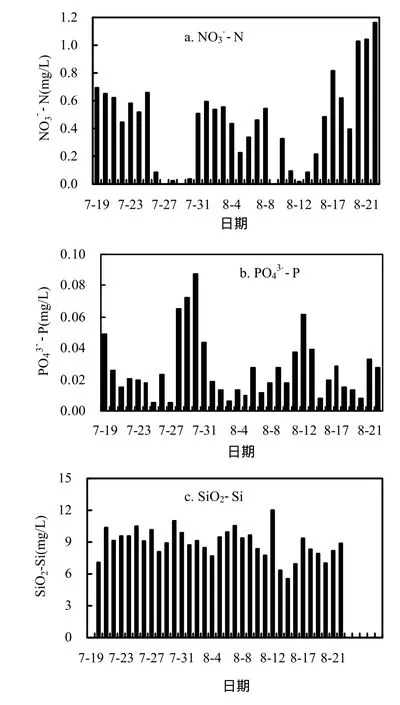
圖7 表層NO3--N, PO43--P、SiO2-Si濃度隨時間的變化Fig.7 NO3--N, PO43--P and SiO2-Si concentrations of surface water in Xiangxi Bay
3 討論
3.1 藍藻水華暴發相關分析
影響藍藻水華暴發的環境因子很多,且不同因子在水華暴發過程中所起的作用大不相同,其中溫度、水體光學特性、水動力特性、營養鹽濃度被大多數研究認為是直接促使藍藻形成水華的重要條件[25].水華的發生不是瞬時的,而是浮游植物生物量在水體中不斷增加的過程[26].一般認為水華的暴發是適宜的水溫、光照、水動力條件和充足的營養共同作用的結果.監測過程中香溪河水流流速緩慢,營養鹽含量豐富[24],總氮、總磷含量均大于富營養化閥值,因此可以推測水溫、Zmix/Zeu是誘發藍藻水華的主要控制因子.
相關分析表明,整個監測期間藍藻密度與水溫、水體穩定系數呈顯著正相關關系,與混合層/真光層(Zmix/Zeu)、混合層、硝酸鹽濃度、TN/TP呈顯著負相關關系.藍藻生長的主要控制因子是水溫、Zmix/Zeu、水體穩定系數.
3.2 藍藻水華影響因子
水溫與藍藻密度具有顯著的正相關關系,研究表明[27],在營養物質充足、水體較為穩定等條件具備時,適宜的高水溫是藍藻生長旺盛的外界誘發因子.監測發現,藍藻密度對水溫有很好的響應,隨著溫度的升高,藍藻光合作用速率加快,豐度增加趨勢明顯,在高水溫的誘發下藍藻大量聚集在表層形成明顯的水華現象.綠藻在溫度較高時雖然可以形成水華[2],但當溫度維持較高水平時,魚腥藻可以產生藻毒素來抑制綠藻生長增殖[28].同時其他藻類在 25℃以上時生長速率緩慢[29],耐受不住高水溫,因此高溫條件下藍藻更具有競爭優勢暴發水華.可以推測,藍藻增殖成為優勢種群和藍藻水華發生是高水溫的誘發結果.
香溪河庫灣夏季的持續高溫,營養物質充足,藻類的生長更多的取決于接受有效光照的幾率,這與混合層與真光層之比密切相關;由于香溪河庫灣光照強度較高,加之水體上下混合作用微弱,減小了混合層(藻類可以懸浮分布的水層范圍)和真光層(藻類維持宏觀凈增長的水層范圍)的比值,有助于具有懸浮機制的魚腥藻在真光層中自由遷移,以便接受充足光照、營養鹽迅速繁殖而形成水華[30].隨著水體摻混劇烈,混合層厚度逐漸增大,分層不明顯,表層混合水體易將藻類攜帶至真光層以下,降低藻類接受充足光照的幾率,增大藻類在真光層下的死亡率[31],則浮游植物生長增殖受到限制[32].對比圖3、圖6可知,Zmix/Zeu與藍藻密度變化呈相反趨勢,水華期間Zmix/Zeu小于 1,水柱垂向混合的范圍在真光層深度以上,藻類進行充分光合作用并超過呼吸作用消耗,有利于水華的暴發,同時在Zmix/Zeu為0.5左右時,水華強度最大;水華后期Zmix/Zeu大于1,水體紊動劇烈,水柱混合均勻,混合層深度顯著增加,在混合過程中,聚集在表層的藻類易被攜帶至下部水體,在混合層內的藻類會受到光照和營養鹽的限制,降低水體初級生產力,水華消退.

表1 環境因子與藍藻密度相關系數Table 1 Correlation coefficients between some environmental factors and cell density in Xiangxi Bay during the summer
魚腥藻屬固氮藻類,當周圍氮源濃度降低時,魚腥藻的部分營養細胞會轉變為含固氮酶的異形胞,起到固氮作用,是較強的氮競爭者[28].監測期間,魚腥藻對硝氮利用明顯,而氮濃度的降低更加顯現了魚腥藻的競爭優勢,從而更好的進行生長增殖.
研究表明,風對藻類的遷移有一定的影響,過大的風浪作用不利于藻類的富集.風的剪切、拖拽作用引起表層水體的擾動摻混,會一定程度加深Zmix深度.本研究中最大風速為4.2m/s,平均風速為1.1m/s,且風速大小變化不顯著,風向以偏南風為主.香溪河庫灣地形狹窄且曲折,吹程通常不超過2km[24],較之水溫分層的變化,風對香溪河庫灣水體混合層深度的影響有限.對于深水型水庫,藻密度的分布受控于水動力擾動強度,水體垂向混合程度比風速對藻類垂向混合影響更為顯著.監測期間,Zeu深度隨時間變化較小, 因此,Zmix深度的變化直接決定著支流庫灣水華的生消,極小的水體混合層深度導致了夏季藍藻水華占優.異重流現象是導致 Zmix深度變化的主要影響因素,根據紀道斌等[33]的研究結果,可以通過三峽水庫潮汐調度來改變水位波動方式,從而改變支流分層異重流的潛入方式,打破支流水溫分層狀態,進而迅速擴大支流庫灣的Zmix深度,使Zmix深度遠大于 Zeu深度,影響水下垂向光熱條件,可促使藍藻水華消退.
4 結論
4.1 夏季香溪河庫灣暴發了嚴重的藍藻水華,優勢藻種為魚腥藻,水華期間魚腥藻通過自身懸浮機制、固氮機制增加獲取更多光照、營養鹽的幾率,同時因其能夠產生藻毒素,可以抑制其他藻類生長,增加其競爭優勢,從而更好的進行生長增殖.
4.2 研究期間,香溪河庫灣水體存在顯著分層,氮磷營養含量充足,水華期間藍藻對硝氮利用顯著,在營養物質充足、庫灣水體穩定等條件具備的情況下,適宜的水溫是影響藍藻生長的關鍵因子;水溫的持續升高、Zmix/Zeu減小是藍藻顯著增殖并暴發水華的誘發因子并且在 Zmix/Zeu為 0.5左右時最有利于藍藻的生長.水華后期水體表層紊動劇烈,打破庫灣水溫分層狀態,迅速擴大 Zmix深度,導致 Zmix/Zeu增大從而限制藻類生長增殖,水華消退.
[1] 邱光勝,胡 圣,葉 丹,等.三峽庫區支流富營養化及水華現狀研究 [J]. 長江流域資源與環境, 2011,20(3):311-316.
[2] 張 敏,蔡慶華,王 嵐,等.三峽水庫香溪河庫灣藍藻水華生消過程初步研究 [J]. 濕地科學, 2009,7(3):230-236.
[3] 許秋瑾,鄭丙輝,朱延忠,等.三峽水庫支流營養狀態評價方法[J]. 中國環境科學, 2010,30(4):453-457.
[4] 李 哲,郭勁松,方 芳,等.三峽小江回水區藍藻季節變化及其與主要環境因素的相互關系 [J]. 環境科學, 2010,31(2):301-309
[5] 易仲強,劉德富,楊正健,等.三峽水庫香溪河庫灣水溫結構及其對春季水華的影響 [J]. 水生態學雜志, 2009(005):6-11.
[6] 王光謙,方紅衛.異重流運動基本方程 [J]. 科學通報, 1996, 41(018):1715-1720.
[7] 魏復盛,國家環境保護總局,水和廢水監測分析方法編委會.水和廢水監測分析方法 [M]. 北京:中國環境科學出版社, 2002.
[8] 胡鴻鈞.中國淡水藻類 [M]. 上海:上海科學技術出版社, 1980.
[9] 周鳳霞,陳劍虹.淡水微型生物圖譜 [M]. 北京:化學工業出版社, 2005.
[10] Lawson R, Anderson M A. Stratification and mixing in Lake Elsinore, California: An assessment of axial flow pumps for improving water quality in a shallow eutrophic lake [J]. Water Research, 2007,41(19):4457-4467.
[11] 張書農.環境水力學 [M]. 南京:河海大學出版社, 1988.
[12] Lawrence I, Bormans M, Oliver R, et al. Physical and nutrient factors controlling algal succession and biomass in Burrinjuck Reservoir [Z]. Land and Water Resources Australia, 2000.
[13] 鄭建軍,鐘成華,鄧春光.試論水華的定義 [J]. 水資源保護, 2006,22(5):45-47.
[14] 金相燦,屠清瑛.環境科學. 湖泊富營養化調查規范 [M]. 北京:中國環境科學出版社, 1990.
[15] 王海云,程勝高,黃 磊.三峽水庫 “藻類水華” 成因條件研究[J]. 人民長江, 2007,38(2):16-18.
[16] 陳宇煒,秦伯強,高錫云.太湖梅梁灣藻類及相關環境因子逐步回歸統計和藍藻水華的初步預測 [J]. 湖泊科學, 2001,13(1): 63-71.
[17] 高月香,張永春.水文氣象因子對藻華爆發的影響 [J]. 水科學與工程技術, 2006(2):10-12.
[18] Zhang Y, Prepas E E. Regulation of the dominance of planktonic diatoms and cyanobacteria in four eutrophic hardwater lakes by nutrients, water column stability, and temperature [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1996,53(3):621-633.
[19] Steinberg C E W, Hartmann H M. Planktonic bloom‐forming Cyanobacteria and the eutrophication of lakes and rivers [J]. Freshwater Biology, 1988,20(2):279-287.
[20] Kawara O, Yura E, Fujii S, et al. A study on the role of hydraulic retention time in eutrophication of the Asahi River Dam reservoir [J]. Water Science and Technology, 1998,37(2):245-252.
[21] 江啟明,侯 偉,顧繼光,等.廣州市典型中小型水庫營養狀態與藍藻種群特征 [J]. 生態環境學報, 2010,19(010):2461-2467.
[22] Khanna D R, Bhutiani R, Chandra K S. Effect of the euphotic depth and mixing depth on phytoplanktonic growth mechanism [J]. International Journal of Environmental Research, 2009, 3(2):223-228.
[23] Davis J R, Koop K. Eutrophication in Australian rivers, reservoirs and estuaries–a southern hemisphere perspective on the science and its implications [J]. Hydrobiologia, 2006,559(1):23-76.
[24] Liu L, Liu D, Johnson D M. Effects of vertical mixing on phytoplankton blooms in Xiangxi Bay of Three Gorges Reservoir: Implications for management [J]. Water Research, 2012,1(1): 2121–2130
[25] Havens K E, James R T, East T L, et al. N: P ratios, light limitation, and cyanobacterial dominance in a subtropical lake impacted by non-point source nutrient pollution [J]. Environmental Pollution, 2003,122(3):379-390.
[26] 孔繁翔,高 光.大型淺水富營養化湖泊中藍藻水華形成機理的思考 [J]. 生態學報, 2005,25(3):589-595.
[27] 王 煒,余衛東.千島湖藍藻生物量制約因素分析 [J]. 云南環境科學, 2002,21(4):44.
[28] 戴瑾瑾,陳德輝,高云芳,等.藍藻毒素的研究概況 [J]. 武漢植物學研究, 2009,27(001):90-97.
[29] 史小麗,王鳳平,蔣麗娟,等.擾動對外源性磷在模擬水生態系統中遷移的影響 [J]. 中國環境科學, 2002,22(6):537-541.
[30] Jones G J, Poplawski W. Understanding and management of cyanobacterial blooms in sub-tropical reservoirs of Queensland, Australia [J]. Water Science and Technology, 1998,37(2):161-168.
[31] Doyon P, Klein B, Ingram R G, et al. Influence of wind mixing and upper-layer stratification on phytoplankton biomass in the Gulf of St. Lawrence [J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2000,47(3/4):415-433.
[32] Whitton B A, Potts M. The ecology of cyanobacteria: their diversity in time and space [M]. Amsterdam. Kluwer Academic Publishers, 2000.
[33] 紀道斌,劉德富,楊正健,等.汛末蓄水期香溪河庫灣倒灌異重流現象及其對水華的影響 [J]. 水利學報, 2010,6:691-702.
Cyanobacterial bloom in Xiangxi Bay, Three Gorges Reservoir.
TIAN Ze-bin1, LIU De-fu1,2*, YANG Zheng jian1,3, FANG Xiao-feng1, YAO Xu-jiao1, FANG Li-juan1,2(1.Engineering Research Center of Eco-environment in Three Gorges Reservoir Region, Ministry of Education, College of Hydroelectric and Environmental Engineering, China Three Gorges Univerxity, Yichang 443002, China;2.College of Resources and Environment Sciences, Hubei University of Technology, Wuhan 430072, China;3.College of Resources and Environment Sciences, Hubei University of Technology, Wuhan 430072, China). China Environmental Science, 2012,32(11):2083~2089
A cyanobacteria bloom was observed in Xiangxi Bay of Three Gorges Reservoir in summer of 2010. Based on field data collected from 19 July to 30 August 2010, this paper descripted the cyanobacteria bloom process and analyzed the influence factors. The bloom lasting for about 25 days, broke out on 23 July, and ended on 16 August, cell density summit reached 108.03×106cell/L. The dominant species was Anabaena, and the major reasons were the suspension and nitrogen fixation mechanism, along with the capacity to produce microcystin to inhibit the growth of other algal. Specific stratification model of water and adequate nutrients were necessary to the cyanobacteria bloom, and nitrate might be the major nitrogen source. Under the conditions of undisturbed water situation and adequate nutrients, the proliferation rate of cyanobacteria might be accelerated gradually with increasing water temperature and lowering Zmix/Zeu. Especially when Zmix/Zeuwas less than 0.5, it was more favorable to cyanobacteria growth.
Three Gorges Reservoir;Xiangxi Bay;cyanobacteria bloom;mixing depth
2012-02-25
國家水體污染控制與治理科技重大專項(2009ZX07104-001,2009ZX07104-004,2009ZX07104-003-03);“十一五”國家科技支撐計劃(2008BAB29B09)
* 責任作者, 教授, dfliu@ctgu.edu.cn
X524
A
1000-6923(2012)11-2083-07
田澤斌(1989-),女,河北秦皇島人,三峽大學碩士研究生,主要從事生態水工方面的研究.
