歐李果實采后不同形態鈣的質量分數及組成變化
2012-11-24 07:08:30馬建軍張立彬于鳳鳴任艷軍
浙江農林大學學報 2012年3期
關鍵詞:質量
馬建軍,張立彬,杜 彬,于鳳鳴,任艷軍
(1.河北科技師范學院 分析測試中心,河北 秦皇島 066004;2.河北科技師范學院 園藝科技學院,河北秦皇島066004;3.河北科技師范學院 生命科技學院,河北 秦皇島066004)
歐李Prunus humilis是中國特有的一種野生果樹資源,其果實富含糖、蛋白質和維生素C,同時含有大量兒童所必需的氨基酸,尤以果實中鈣、鐵含量高而受到消費者的歡迎[1]。因此,研究揭示野生歐李果實中鈣素營養及果實采后鈣素營養生理變化,對其開發利用具有重要意義。研究表明[2-4]:果實缺鈣或鈣與其他元素之間失衡是果實生理病害發生的主要原因,而歐李果實生理病害少可能與果實鈣含量高有關,因為果實組織中如果維持較高的鈣水平,可以保持果實硬度,降低呼吸速率,抑制乙烯的產生,延長果實儲藏壽命,提高果實的商品價值[5-7]。筆者研究發現[8-9]:歐李果實鈣累積主要發生在幼果細胞分裂期和果實細胞膨大期2個階段,細胞分裂期以果膠鈣形態積累為主,而細胞膨大期以水溶性鈣形態積累為主,尤其是與樹體內水溶性鈣和果膠鈣含量變化密切相關。龔云池等[10]研究比較了鴨梨Pyrus bertschneideri和雪花梨Pyrus bretschneideri,陳麗漩等[11]研究了荔枝Litchi chinensis果實儲藏過程中的不同形態鈣含量變化規律,歐李成熟果實中不同形態鈣的含量組成及其采后鈣素生理變化規律尚未見報道。為此,以實生和嫁接類型歐李為試材,通過研究歐李采收期和儲藏期果實中不同形態鈣質量分數組成及變化,探明歐李果實鈣素形成及轉化規律,以期揭示歐李鈣素營養吸收特性,為開發利用野生歐李資源,提高人類鈣素營養及健康水平提供基礎理論資料。
1 材料和方法
1.1 材料
歐李實驗材料分為2種類型。一是實生類型:包括小葉小果型,葉片淡綠,果實質量5 g左右;大葉大果型,葉片濃綠,果實質量10 g左右,樹齡為10年生。實生材料均來源于燕山山脈野生歐李群體,并定植于河北科技師范學院園藝科技學院試驗站野生歐李資源圃內。圃地土壤肥力中等,樹體自然生長,叢生,株高為70~80 cm。二是嫁接類型:包括高位嫁接型(接口距地面80 cm)和低位嫁接型(接口距地面30 cm)。采用實生類型中的大葉大果型接穗嫁接在毛桃Prunus persica砧木上,樹齡為8年生。2種類型歐李均在3月上旬芽體萌動,4月中旬開花,8月上中旬果實成熟。
1.2 方法
在果實生理成熟期,分別選取2種類型中包括小葉小果型、大葉大果型、高位嫁接型和低位嫁接型的植株各3株,作為各時期鈣組分提取的樣本株,單株3次重復;在每株不同方位的結果基生枝中上部位采集不同類型成熟度一致,大小均勻,無機械損傷和無病蟲害的果實樣品各45個;果實采后放置在室溫(25±1)℃條件下自然儲藏,采后當日(8月13日)從每株采集的果實樣品中隨機選取15個果實清洗干凈,勻漿處理后提取不同形態鈣組分,并于1周(8月21日)和2周(8月29日)后分別提取測定1次,共提取測定鈣3次。
不同形態鈣的提取參照小西茂毅等[12]的方法,并加以改進。精確稱取經組織搗碎機勻漿后的歐李果實樣品 10.00 g,分別采用無離子水、 1 mol°L-1氯化鈉、 20 g°kg-1乙酸、 50 g°kg-1鹽酸浸提劑逐級提取的方法,分別提取水溶性鈣(活性鈣)、氯化鈉溶性鈣(果膠鈣)、醋酸溶性鈣(磷酸鈣)和鹽酸溶性鈣(草酸鈣)。將浸提液樣品轉移至離心管中,25℃恒溫水浴提取1 h,后用高速冷凍離心機離心10 min;吸取上清液轉移至100 mL容量瓶中,加入相應提取液洗滌沉淀物,如此反復提取3~4次;之后用相應的浸提液定容,同時加入釋放劑氯化鍶溶液(質量濃度為1 g°L-1)排除干擾影響;剩余殘渣中主要成分是硅酸鈣(簡稱為剩余鈣)[12],采用 HNO3-HClO4(5 ∶1)混合酸消化樣品待測。
樣品中不同形態鈣質量分數采用3200型原子吸收分光光度計(上海分析儀器廠)測定,測定波長為422.7 nm,鈣質量分數(mg°kg-1)=待測液測得質量濃度(mg°L-1)×定容體積(V)/樣品質量(W)。數據處理采用SPSS統計分析軟件。
2 結果與分析
2.1 采收期實生類型果實中不同形態鈣的質量分數變化
由表1可見:歐李實生類型果實中不同形態鈣質量分數因果型大小存在差異,小果型果實中水溶鈣、磷酸鈣和草酸鈣的質量分數極顯著高于大果型果實中相應的鈣形態組成(P<0.01),而大果型果實中的果膠鈣質量分數極顯著高于小果型果實中果膠鈣的質量分數(P<0.01);大小果型果實中的活性鈣組分(水溶鈣和果膠鈣)分別占大小果型鈣組分總量的69.11%和63.79%;大小果型果實中不同形態鈣提取率分別占總鈣量的98.43%和98.76%。研究表明:歐李小果型果實中鈣質量分數顯著高于大果型果實中的鈣質量分數(P<0.05),大小果型不同形態鈣質量分數大小為水溶鈣>果膠鈣>磷酸鈣>草酸鈣>剩余鈣。
2.2 采收期嫁接類型果實中不同形態鈣的質量分數變化
由表1可見:歐李嫁接類型果實中不同形態鈣質量分數因接穗嫁接高低存在差異。低位嫁接類型果實中水溶鈣、磷酸鈣和草酸鈣質量分數顯著或極顯著高于高位嫁接類型果實中相應鈣形態組成(P<0.01或P<0.05)。高位和低位嫁接類型果實中的活性鈣組分(水溶鈣和果膠鈣)分別占相應嫁接類型鈣組分總量的74.86%和64.99%;高位和低位嫁接類型果實中不同形態鈣提取率分別占總鈣量的98.24%和99.32%。研究表明:歐李低位嫁接類型果實中鈣質量分數顯著高于高位嫁接類型果實中的鈣質量分數(P<0.05)。由表1還可以看出:與大果型果實比較,低位嫁接果實中水溶鈣質量分數顯著高于實生類型(P<0.05),而高位嫁接基本與實生類型持平,水溶鈣可提取率增加;嫁接類型果實中果膠鈣質量分數極顯著低于實生類型中果膠鈣組分(P<0.01),果膠鈣可提取率下降;嫁接類型果實中不同形態鈣質量分數占果實總鈣量的比率其大小變化與實生類型一致。由此可見,歐李嫁接并未改變果實中不同形態鈣的質量分數組成大小,但歐李嫁接和嫁接接口高低在一定程度上影響了果實中鈣及不同形態鈣的質量分數變化。

表1 歐李實生和嫁接類型果實中不同形態鈣質量分數及組成Table 1 Content and composition of diverse forms calcium in Prunus humilis grafting fruits and seedling stand fruits during harvest period
2.3 儲藏期實生類型果實中不同形態鈣的組成百分率變化
研究結果顯示(圖1和圖2):歐李實生類型果實中不同形態鈣隨果實儲藏期的延長其鈣組成百分率變化呈現一定的變化規律,2種果型果實中水溶鈣呈增加趨勢,果膠鈣呈下降趨勢,磷酸鈣和剩余鈣變化不明顯,草酸鈣在大果型果實中呈先增后降趨勢,在小果型果實中呈逐漸下降趨勢;但不同形態鈣組成百分率變化幅度因果型大小而存在差異,大果型和小果型果實儲藏后期(8月29日)較采收期(8月13日),水溶鈣形態分別增加了20.52%(P<0.01)和5.07%(P>0.05)。果膠鈣形態分別下降了13.18% (P<0.05)和0.72%(P>0.05),草酸鈣形態分別下降了7.37%(P>0.05)和5.53%(P>0.05)。由此可見:野生歐李果實采后儲藏存在鈣形態轉化,且其轉化程度與果型大小有關。
2.4 儲藏期嫁接類型果實中不同形態鈣的組成百分率變化

圖1 歐李大果型果實儲藏期各形態鈣的組成百分率的變化Figure 1 Composition percentage change of diverse froms calcium of Prunus humilis large fruits during storage period
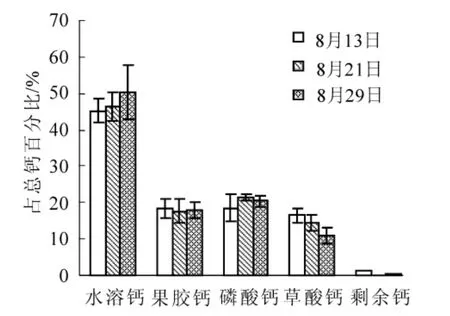
圖2 歐李小果型果實儲藏期各形態鈣的組成百分率的變化Figure 2 Composition percentage change of diverse froms calcium of Prunus humilis small fruits during storage period
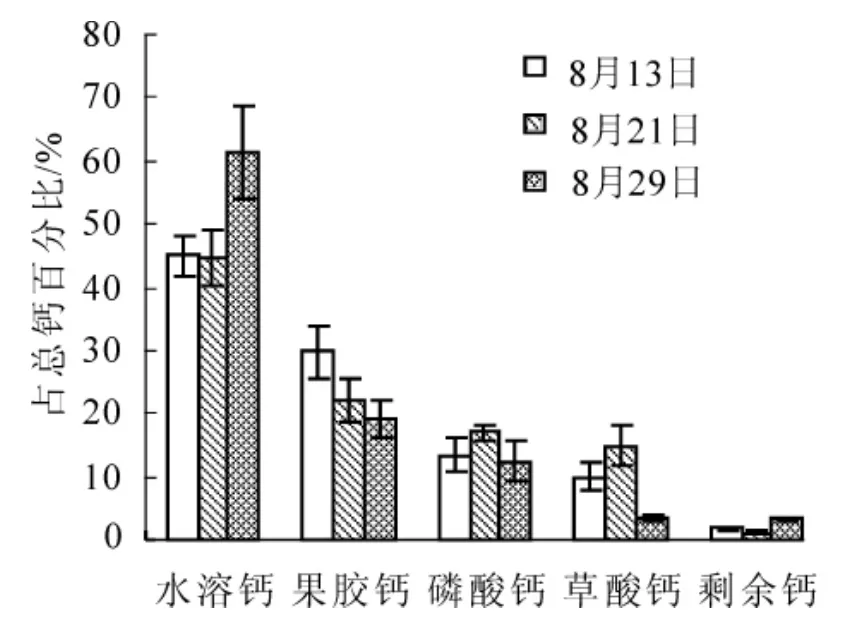
圖3 歐李高位嫁接果實儲藏期各形態鈣的組成百分率的變化Figure 3 Composition percentage change of diverse froms calcium of Prunus humilis topgraftingfruits during storage period
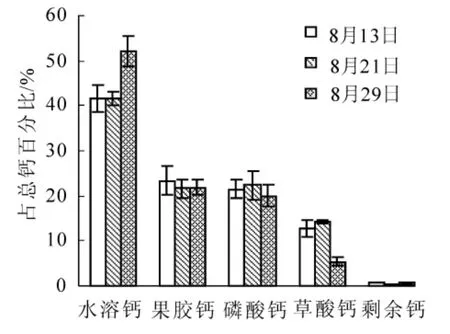
圖4 歐李低位嫁接果實儲藏期各形態鈣的組成百分率的變化Figure 4 Composition percentage change of diverse froms calcium of Prunus humilis lowgraftingfruits during storage period
研究結果顯示(圖3和圖4):歐李嫁接類型果實中不同形態鈣隨果實儲藏期的延長其鈣組成百分率變化與實生類型變化規律基本一致,但不同形態鈣組成百分率變化幅度因嫁接類型不同而存在差異,高位嫁接和低位嫁接果實儲藏后期(8月29日)較采收期(8月13日),水溶鈣形態分別增加了16.36%(P<0.01)和10.55%(P<0.05),果膠鈣形態分別下降了10.65%(P<0.05)和1.52%(P>0.05),草酸鈣形態分別下降了6.31%(P>0.05)和7.40%(P>0.05)。由此可見:野生歐李嫁接類型果實采后儲藏同樣存在鈣形態轉化,且與實生類型轉化規律一致。
3 討論
3.1 采收期歐李果實中不同形態鈣質量分數變化
鈣一般以水溶鈣、果膠鈣、磷酸鈣、草酸鈣及殘余的硅酸鈣(剩余鈣)等形態存在于樹體和果實中,其中果膠鈣是細胞壁的主要鈣形態,草酸鈣和大部分磷酸鈣則沉淀在液泡內[12-13],果膠鈣和水溶鈣為活性鈣[14],尤其是水溶鈣有利于鈣離子的轉移和吸收利用。野生歐李與鴨梨、雪花梨以及荔枝果實中不同形態鈣的含量組成相比較[10-11],均以水溶鈣形態含量組成比例最高,并且小果型果實中總鈣和水溶鈣含量明顯高于大果型果實(P<0.05或P<0.01),故小果型歐李更適合人體補充鈣質。同時結果顯示:果實中不同形態鈣質量分數變化與果實果型大小有關,其中大果型果實中果膠鈣的質量分數顯著高于小果型果實中相應鈣形態質量分數(P<0.01),表明大果型果實中在細胞壁中膠層結合了更多的鈣,這對增強細胞壁的剛性是有益的;而小果型果實中磷酸鈣和草酸鈣形態的質量分數顯著高于大果型果實中相應鈣形態的質量分數(P<0.01),一方面可以推斷,小果型果實生長發育過程中代謝過程比較旺盛,與代謝過程中產生的有機酸如草酸結合形成草酸鈣結晶,避免因有機酸過量積累而產生毒害[15],而磷酸鈣的形成具有一種解毒作用[16];另一方面說明,小果型果實發育過程中旺盛的代謝活動,果實發育庫源需求更多鈣的供應,可能是導致小果型果實水溶鈣形態組分顯著高于大果型果實相應鈣形態的主要原因。當然,果實因果型大小不同造成其不同形態鈣組成差異的原因尚有待進一步研究。嫁接歐李果實中不同形態鈣的含量組成比例無明顯改變,表明嫁接歐李果實中保持了鈣素營養的生理遺傳特性,但嫁接歐李類型因嫁接和嫁接接口的高低對果實中總鈣及不同形態鈣的質量分數產生一定的影響,這可能與嫁接砧木根系分布及形態特征差異以及高位或低位嫁接影響鈣運輸距離等因素有關,其機制有待進一步研究。
3.2 儲藏期歐李果實中不同形態鈣的組成百分率變化
雪花梨果肉中水溶鈣形態組成百分率明顯下降,草酸鈣形態組成百分率明顯增加[10];鴨梨果肉中水溶鈣形態組成百分率增加,果膠鈣形態組成百分率下降[10];荔枝果實中水溶鈣形態組成百分率逐漸下降,果膠鈣形態和草酸鈣形態有所增加[11],表明不同品種果實儲藏期間鈣形態的轉化差異與果實采后成熟生理變化差異有關。歐李果實采后儲藏水溶鈣形態組成百分率明顯增加,表明歐李果實采后儲藏一段時期補鈣效果更好,更有利于人體對鈣質的吸收和利用;果膠鈣形態組成百分率降低,表明果肉細胞壁中膠層結合的鈣量減少,可能會影響原生質的黏滯性、細胞壁的剛性以及細胞膜的穩定性,而原生質黏滯性的降低、細胞壁剛性的減弱等生理變化是果實儲藏過程中衰老的表現[10]。另一方面,歐李果實中草酸鈣形態組分百分率的下降,可能是由于隨果實儲藏期的延長,細胞呼吸代謝衰減產生的有機酸(如草酸等)量減少所致。由此看出:細胞溶質中調節鈣離子含量機能的任何破壞,將引起細胞代謝活動的嚴重紊亂[17],其機制有待進一步深入研究。
[1]曹琴,杜俊杰,劉和,等.野生歐李營養特性分析[J].中國野生植物資源,1999,18(1):34-36.CAO Qin,DU Junjie,LIU He,et al.Nutrient characteristic analysis of Prunus humilis [J].Chin Wild Plant Resour,1999,8 (1): 34-36.
[2]FERGUSON I B,WATKINS C B.Biteer pit in apple fruit[J].Hort Rev,1989,11: 289-355.
[3]WITNEY G W,KUSHAD M M.Correlation of pyruvate kinase activity with bitter pit development in apple fruit [J].Sci Hort,1990,43: 247-253.
[4]YAMAMOTO T,SATOH H,WATANABE S.The effects of calcium and naphthalene acetic acid sprays on cracking index and natural rain cracking in sweet cherry fruits [J].J Jpn Soc Hort Sci,1992,61 (3): 507-511.
[5]FERGUSON I B,DROBAK B K.Calcium and the regulation of plant growth and Senescence [J].Hort Sci,1988,23:262-266.
[6]CHEVERRY J L,PONLIQUEN J,GUYADER H L E,et al.Calcium regulation of exogenous and endogenous aminocy clopropane carboxylic acid bioconversion to ethylene [J].Physiol Plant,1988,74: 53-57.
[7]SINGH S B.Effect of different pre and post harrest treatments on storage life of malta CV blood red fruits [J].Prog Hortic,1987,19 (1/2): 10-16.
[8]馬建軍,張立彬,于鳳鳴,等.野生歐李果實中不同形態鈣的含量及分布[J].園藝學報,2007,34(3):755-759.MA Jianjun,ZHANG Libin,YU Fengming,et al.Contents and distribution of different calcium in Prunus humilis[J].Acta Horiic Sin,2007,34 (3): 755-759.
[9]馬建軍,張立彬,劉玉艷,等.野生歐李生長期組織器官中不同形態鈣含量的變化及其相關性[J].園藝學報,2008,35 (5): 631-636.MA Jianjun,ZHANG Libin,LIU Yuyan,et al.Variation of different forms of calcium contents and their correlation during the growth stage of Prunus humilis tissues [J].Acta Hortic Sin,2008,35 (5): 631-636.
[10]龔云池,徐季娥,呂瑞江.梨果實中不同形態鈣的含量及其變化的研究[J].園藝學報,1992,19(2):129-134.GONG Yunchi,XU Ji’e,Lü Ruijiang.Studies on the content of different forms of calcium compound and their change in the fruits of pear [J].Acta Hortic Sin,1992,19 (2): 129-134.
[11]陳麗漩,黃玉環,尤瑞璨,等.荔枝儲藏過程的鈣處理及其影響[J].福建果樹,1998(2):4-6.CHEN Lixuan,HUANG Yuhuan,YOU Ruichen,et al.Calcium studies of litchee fruits during storage [J].Fujian Fruits,1998 (2): 4-6.
[12]小西 茂毅,葛西 善三郎.タ(Ⅲ)コ草枼の AgeingにともtíうヵFIシウxviの代謝(Thの 1):生育各期に吸收fh xv ft45Caの形態変化[J].日本土壤肥料學雑,1963,34(3): 67-70.KONISHI Shigeki,KASAI Zenzaburo.Calcium metabolism in the Ageing process of tobacco leaves (part 1) :metabolism of45Ca absorbed at the early and the flowering stage of growth [J].J Sci Soil Manure Jpn,1963,34(3): 67-70
[13]MARSCHNER H.高等植物的礦質營養[M].曹一平,譯.北京:北京農業大學出版社,1991:58-60.
[14]HIMELRICK D G.Determination of total and ionic calcium in apple leaf fruit tissues[J].J Am Soc Horticl Sci,1981,106(5): 619-621.
[15]何念祖,孟賜福.植物營養原理[M].上海:上海科學技術出版社,1987:195-201.
[16]POOVAIAH B W.Molecular and cellular aspects of calcium action in plants [J].HortScience,1988,23 (2): 267-271.
[17]FERGUSON I B,DROBAK B K.Calcium and the regulation of plant growth and senescence [J].HortScience,1988,23 (2): 262-266.
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
