裂褶菌產(chǎn)內(nèi)切β-1,3-葡聚糖酶培養(yǎng)基組成的優(yōu)化研究
2012-11-15 02:07:48暢曉潔鄭必勝
食品工業(yè)科技 2012年5期
暢曉潔,鄭必勝,趙 欣
(華南理工大學(xué)輕工與食品學(xué)院,廣東廣州510640)
裂褶菌產(chǎn)內(nèi)切β-1,3-葡聚糖酶培養(yǎng)基組成的優(yōu)化研究
暢曉潔,鄭必勝*,趙 欣
(華南理工大學(xué)輕工與食品學(xué)院,廣東廣州510640)
采用恒溫?fù)u床培養(yǎng)法培養(yǎng)裂褶菌,研究了裂褶菌產(chǎn)內(nèi)切β-1,3-葡聚糖酶培養(yǎng)基組成中的碳源、氮源及各種無機鹽的單因素參數(shù),并采用四因素三水平正交實驗對主要影響因素進(jìn)行分析,從而確定最佳的產(chǎn)酶培養(yǎng)基配方為:葡萄糖2%,牛肉膏0.3%,NH4Cl 0.1%,KH2PO40.05%,MgSO40.05%。
裂褶菌,內(nèi)切β-1,3-葡聚糖酶,培養(yǎng)基組成
裂褶多糖(Schizophylluan,簡稱SPG)是由藥用真菌裂褶菌分泌的一種中性胞外多糖,且只含有β-D-葡聚糖[1]。獨特的化學(xué)結(jié)構(gòu)使得它在調(diào)節(jié)免疫功能、抗腫瘤、抗輻射等方面有著顯著的療效。但天然裂褶多糖分子量大,在水中溶解度小、粘度大,若將未經(jīng)降解處理的裂褶多糖用于注射治療腫瘤疾病,會引發(fā)肌肉疼痛、血栓等問題[2]。日本學(xué)者提出在提取天然裂褶多糖時必須獲得有效的“活性多糖”,即在提取過程就要對多糖進(jìn)行降解或者改性[3]。研究表明,裂褶菌可產(chǎn)生胞外內(nèi)切β-1,3-葡聚糖酶[3],在內(nèi)切β-1,3-葡聚糖酶的作用下,多聚糖苷鍵斷裂,葡聚糖水解為寡糖或還原糖,即通過酶切來降低β-D-葡聚糖的聚合度而不改變其天然結(jié)構(gòu),不影響或降低其生物活性[4-5]。本文對裂褶菌產(chǎn)內(nèi)切β-1,3-葡聚糖酶的培養(yǎng)基組成進(jìn)行優(yōu)化,確定最佳的培養(yǎng)基組成,為提高裂褶菌產(chǎn)的內(nèi)切β-1,3-葡聚糖酶活力提供實驗依據(jù)。
1 材料與方法
1.1 材料與儀器
裂褶菌(Schizophyllum commune Fr.) 廣東省微生物研究所;斜面種子培養(yǎng)基 PDA培養(yǎng)基;蛋白胨、酵母膏、牛肉膏 廣東環(huán)凱生物科技公司,生化試劑;葡萄糖 分析純,上海博奧生物科技有限公司;冰醋酸、NaAC、無水亞硫酸鈉、酒石酸鉀鈉、NaOH 分析純,天津化學(xué)試劑廠;MgSO4·7H2O、KH2PO4·2H2O、尿素、NH4Cl、NH4NO3、NH4SO4、NH4H2PO4分析純,廣州市東紅化工廠;苯酚、3,5-二硝基水楊酸 分析純,汕頭市光華化學(xué)廠;裂褶多糖 本實驗室制備。
pHS-3C型pH計 上海虹益儀器;HQ45B氣浴恒溫?fù)u床 中科院武漢科儀廠;HSP150生化培養(yǎng)箱 武漢中科儀有限公司;UV-2102PC紫外可見光分光光度計 上海UNICO公司。
1.2 實驗方法
1.2.1 培養(yǎng)基的配制 初始產(chǎn)酶培養(yǎng)基:葡萄糖2%,蛋白胨0.3%,硝酸銨0.25%,硫酸鎂0.05%,磷酸二氫鉀0.05%,pH7.0,121℃滅菌15min。發(fā)酵培養(yǎng)基:配制12°Brix麥芽汁1000mL,稱取1.0g/L NH4Cl溶于其中,121℃滅菌15min[6]。
1.2.2 培養(yǎng)方法 從28℃下培養(yǎng)6d的斜面種子培養(yǎng)基上用接種環(huán)小心刮下孢子,用10mL無菌水洗入預(yù)先滅菌的裝有30mL無菌水的帶玻璃珠三角瓶中,置于搖床上振蕩30min,即得到單孢子懸液(孢子濃度為106/mL)。
將單孢子懸液接種于100mL發(fā)酵培養(yǎng)基中,置于30℃,160r/min搖床上控溫發(fā)酵,發(fā)酵7d后測定酶活力。
1.2.3 酶活力測定方法 以裂褶多糖為底物,采用應(yīng)用最廣泛的還原糖法測定β-1,3-葡聚糖酶活力。
1.2.3.1 DNS試劑的配制[7]稱取10g 3,5-二硝基水楊酸于水中,全部溶解后,加入20g NaOH、200g酒石酸鉀鈉,加入蒸餾水,使總體積至500mL左右,加熱溶解后,加2g苯酚、0.5g無水亞硫酸鈉,加熱攪拌至全部溶解,冷卻,用水稀釋至1000mL,儲于棕色瓶中。
1.2.3.2 標(biāo)準(zhǔn)曲線的制作 準(zhǔn)確稱取葡萄糖100mg,用適量蒸餾水溶解并定容于100mL容量瓶中。分別吸取0、0.1、0.2、0.3、0.4、0.5、0.6mL葡萄糖液,各加水至1.0mL,加入DNS溶液1.5mL,沸水浴5min顯色。冷卻后用蒸餾水定容至25mL,搖勻,在520nm條件下測定吸光值,取1.5mL蒸餾水代替DNS溶液作為空白,繪制標(biāo)準(zhǔn)曲線。
1.2.3.3 裂褶菌產(chǎn)內(nèi)切β-1,3-葡聚糖酶活力的測定 1mL NaAC-HAC緩沖液(0.05mol/L,pH5.0)、1mL酶液、1mL 0.5%裂褶多糖組成反應(yīng)體系,于50℃水浴振蕩反應(yīng)30min,沸水浴5min滅酶活,取上清液1mL用還原糖法測定酶水解液中的還原糖量(以葡萄糖計),以滅活酶作空白。
酶活力單位:在上述反應(yīng)條件下,1min水解β-1,3-葡聚糖釋放出1μmol葡萄糖所需的酶量,即為一個酶活力單位,以U表示。
2 結(jié)果與分析
2.1 還原糖標(biāo)準(zhǔn)曲線
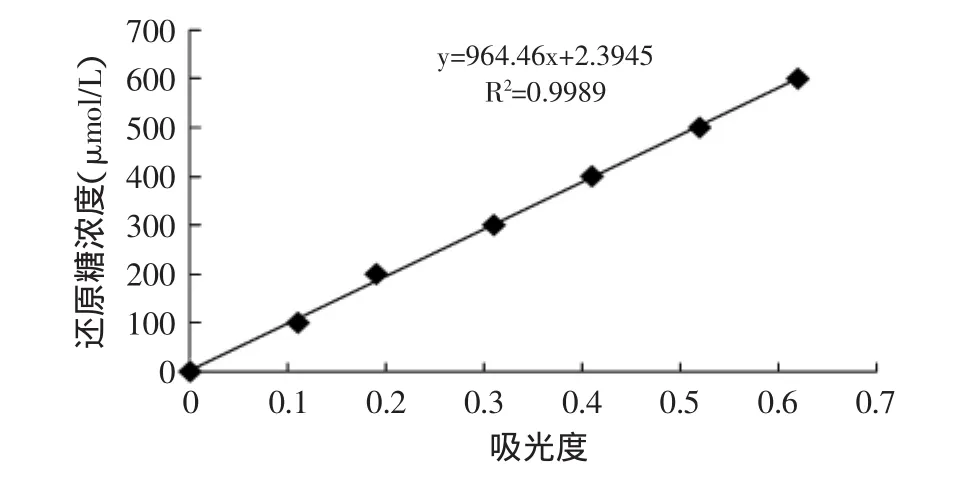
圖1 還原糖法標(biāo)準(zhǔn)曲線Fig.1 Standard curve of reduced sugars
2.2 內(nèi)切β-1,3-葡聚糖酶活力的測定
初步設(shè)計的裂褶菌搖瓶發(fā)酵初始產(chǎn)酶培養(yǎng)基,測得其發(fā)酵液酶活U為3.52u/mL,在此基礎(chǔ)上進(jìn)行了產(chǎn)酶培養(yǎng)基的優(yōu)化研究。
2.2.1 葡萄糖添加量對產(chǎn)酶的影響 研究表明:裂褶菌利用葡萄糖的能力高于二糖和三糖,在培養(yǎng)基中添加少量葡萄糖能夠大幅度地提高產(chǎn)酶量[4]。葡萄糖的起始添加量對產(chǎn)酶影響很大,當(dāng)添加量過低時,裂褶菌提前發(fā)生自溶現(xiàn)象;但添加量過高,酶會受到分解代謝阻遏。
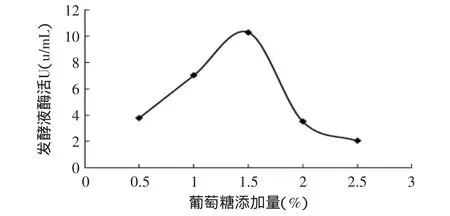
圖2 葡萄糖添加量對產(chǎn)酶的影響Fig.2 Effect of glucose additioin on enzyme activity
分別以0.5%、1%、1.5%、2%、2.5%不同添加量的葡萄糖作為碳源,研究葡萄糖的添加量對產(chǎn)酶的影響。從圖2可知,在葡萄糖添加量較低時,發(fā)酵液酶活U隨著葡萄糖添加量的增大而增大,當(dāng)葡萄糖的添加量較高時,發(fā)酵液酶活U大幅度下降,說明酶受降解物阻遏明顯;最適葡萄糖添加量為1.5%。
2.2.2 不同有機氮源對產(chǎn)酶的影響 分別以蛋白胨、酵母膏、牛肉膏、尿素作為有機氮源,在初始產(chǎn)酶條件下進(jìn)行發(fā)酵,研究等量的不同有機氮源對β-1,3-葡聚糖酶產(chǎn)量的影響。
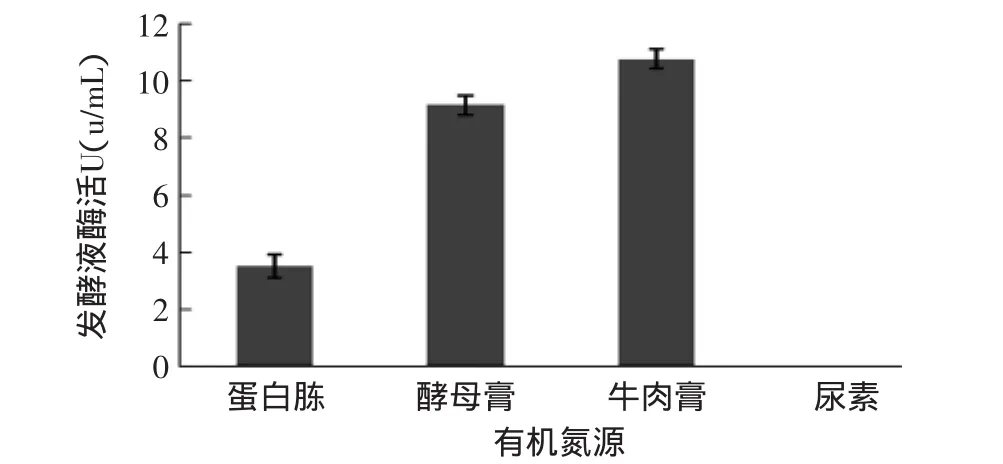
圖3 不同有機氮源對產(chǎn)酶的影響Fig.3 Effect of different organic nitrogenous on enzyme activity
由圖3可知,以牛肉膏為唯一有機氮源的培養(yǎng)基所產(chǎn)的β-1,3-葡聚糖酶活力最高;其次是酵母膏。由于酵母膏和牛肉膏成分復(fù)雜,可能含有微生物生長需要的生長因子,從而促進(jìn)了產(chǎn)酶。以尿素為唯一有機氮源的培養(yǎng)基中裂褶菌生長不好,測不到酶活力。可能是因為經(jīng)過高溫滅菌和發(fā)酵,尿素中的營養(yǎng)成分早已損失殆盡,或是因為裂褶菌屬于木腐菌,尿素加入到培養(yǎng)基中不但在高溫下會分解出游離氨,而且在菌絲生長后也會被菌絲的代謝物降解出游離氨,當(dāng)培養(yǎng)基不能吸收氨態(tài)氮時,氨便以氣態(tài)方式存在,當(dāng)培養(yǎng)基中氨過高時,菌絲便會停止生長至死亡[8]。因此,最佳有機氮源為牛肉膏。
2.2.3 不同無機氮源對產(chǎn)酶的影響 確定葡萄糖的添加量和最佳有機氮源后,對無機氮源進(jìn)行單因素實驗。分別以NH4Cl、NH4NO3、NH4SO4、NH4H2PO4作為無機氮源,添加量以氮元素摩爾數(shù)相等為依據(jù),在初始產(chǎn)酶條件下進(jìn)行發(fā)酵。
由圖4可知,不同無機氮源產(chǎn)酶活力大小順序為:NH4Cl>NH4NO3>NH4H2PO4>(NH4)2SO4。使用(NH4)2SO4、NH4H2PO4和NH4NO3作無機氮源時,裂褶菌生長不好,酶活也不高。可能是由于NH4+被利用后剩余的NO3-、SO42-和H2PO4-使培養(yǎng)基呈酸性,菌絲雖能利用但生長緩慢。用NH4Cl作無機氮源時,對生長和產(chǎn)酶都有明顯的促進(jìn)作用。因此,NH4Cl是最佳無機氮源。
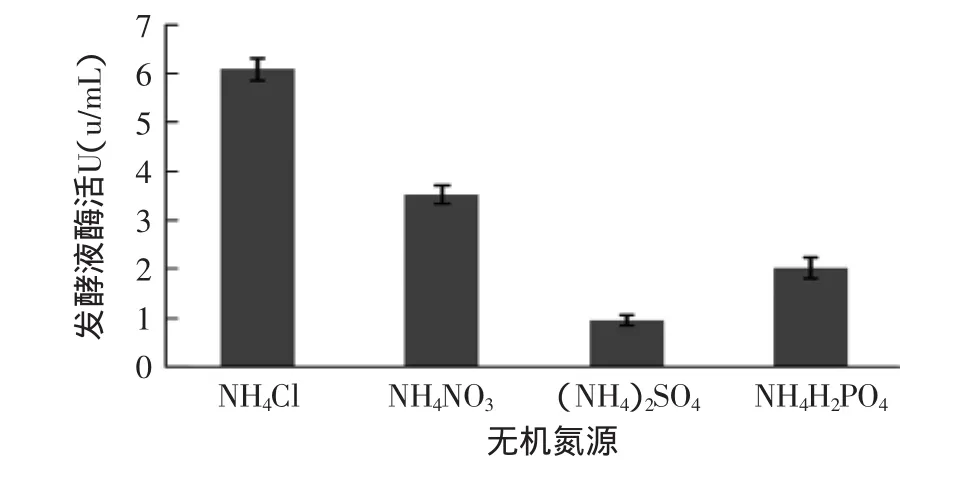
圖4 不同無機氮源對產(chǎn)酶的影響Fig.4 Effect of different inorganic nitrogenous on enzyme activity
2.2.4 MgSO4對產(chǎn)酶的影響 Mg2+是金屬離子中較為重要的一種,它是許多酶活性中心不可缺少的一部分[9]。分別以MgSO4添加量為0.02%、0.05%、0.1%和0.15%在初始條件下進(jìn)行發(fā)酵。
由圖5可知,改變MgSO4的添加量對裂褶菌產(chǎn)內(nèi)切β-1,3-葡聚糖酶的影響并不顯著,當(dāng)MgSO4添加量為0.05%時,內(nèi)切β-1,3-葡聚糖酶活力略高,因此,MgSO4最適添加量為0.05%。
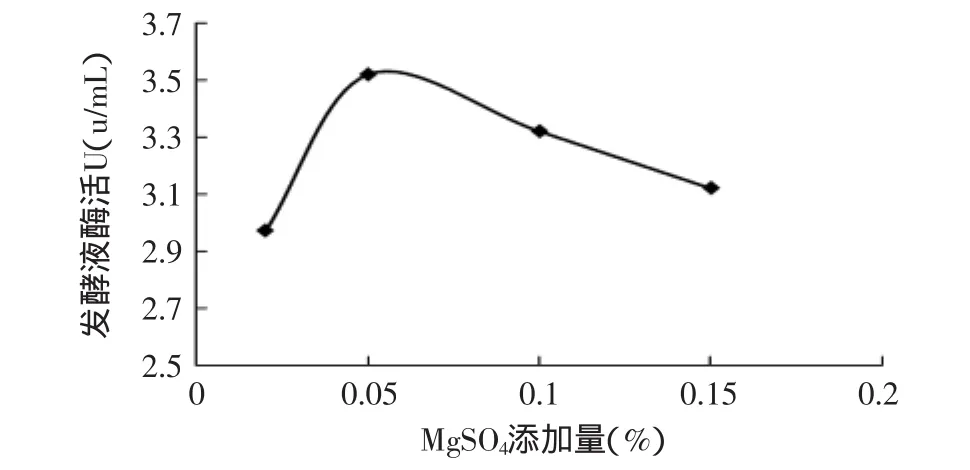
圖5 MgSO4添加量對產(chǎn)酶的影響Fig.5 Effect of MgSO4on enzyme activity
2.2.5 KH2PO4對產(chǎn)酶的影響 磷酸鹽不僅是核酸、酶和能量代謝的組成部分,也是菌絲生長中必不可少的元素。當(dāng)缺乏磷源時,碳源和氮源也不能被很好地利用[10]。因此,磷酸鹽的濃度對裂褶菌的生長和內(nèi)切β-1,3-葡聚糖酶的產(chǎn)生有著較大的影響。
分別以KH2PO4添加量為0.02%、0.05%、0.1%和0.15%在初始條件下進(jìn)行發(fā)酵,結(jié)果見圖6。
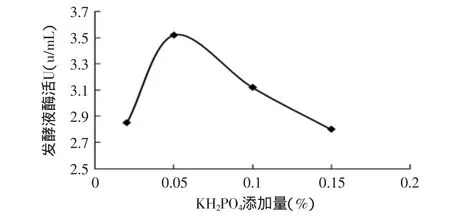
圖6 KH2PO4添加量對產(chǎn)酶的影響Fig.6 Effect of KH2PO4on enzyme activity
從圖6可知,KH2PO4的濃度對內(nèi)切β-1,3-葡聚糖酶產(chǎn)酶水平的影響表現(xiàn)出最適濃度效應(yīng)。低濃度的KH2PO4有利于內(nèi)切β-1,3-葡聚糖酶的產(chǎn)生,內(nèi)切β-1,3-葡聚糖酶的產(chǎn)酶水平隨著KH2PO4濃度的增加而升高;當(dāng)KH2PO4濃度達(dá)一定值后繼續(xù)升高時,內(nèi)切β-1,3-葡聚糖酶活性反而降低。因此,KH2PO4最適添加量為0.05%。
2.3 正交實驗
在確定了最佳碳源和最佳氮源(包括有機氮源和無機氮源)基礎(chǔ)上,對葡萄糖、牛肉膏和NH4Cl采用正交法設(shè)計實驗,選用四因素三水平正交實驗表L9(34)(見表2)以確定最佳培養(yǎng)基配方。
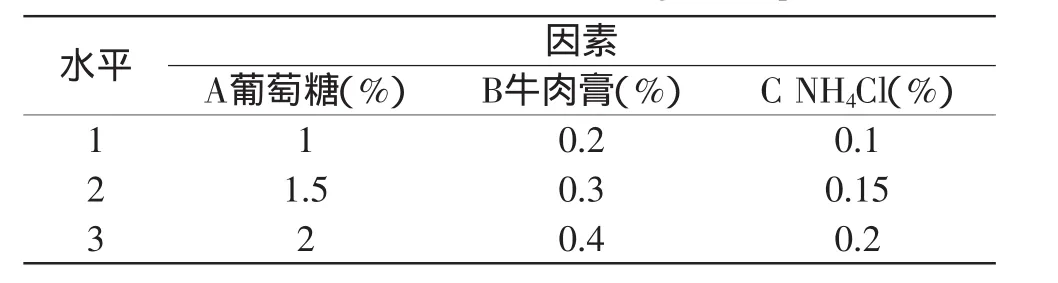
表1 正交實驗因素水平表Table 1 Factors and levels of orthogonal experiment
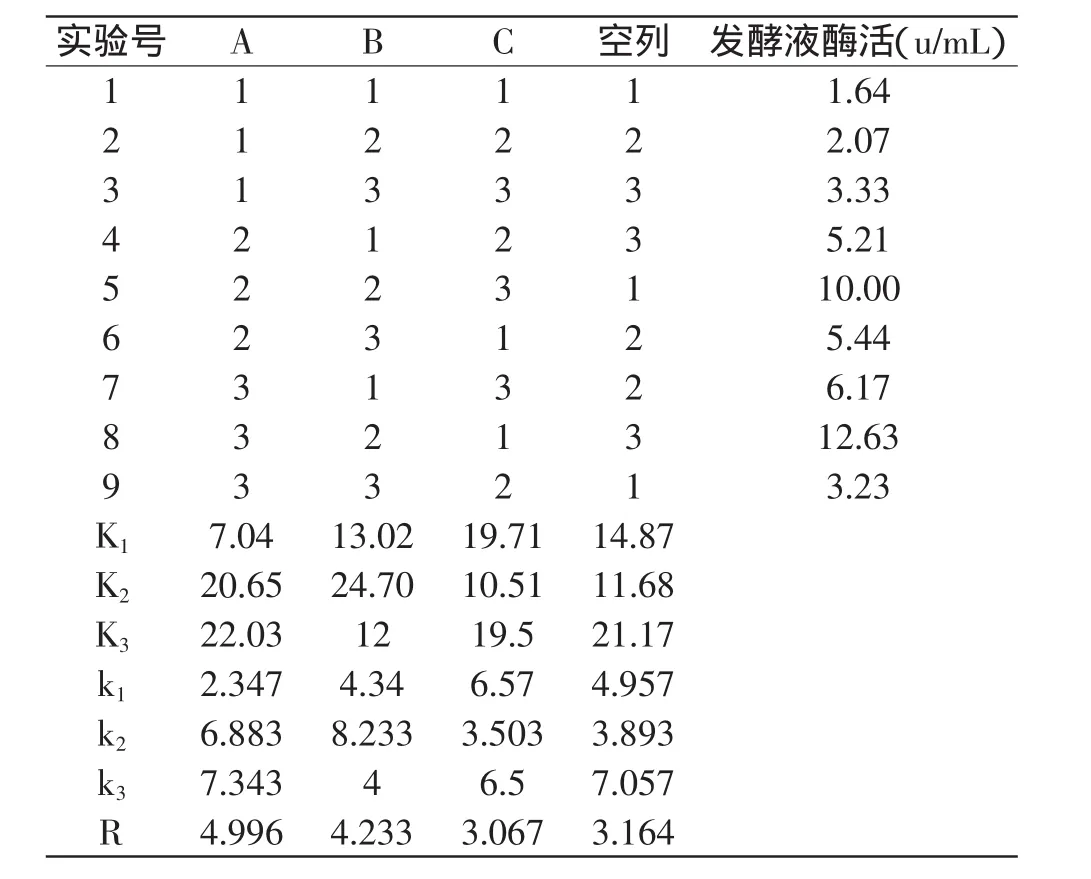
表2 正交實驗結(jié)果與分析Table 2 Results and analysis of orthogonal experiment
根據(jù)表2結(jié)果可知,三因素對產(chǎn)酶影響的主次順序為:葡萄糖>牛肉膏>NH4Cl。同時比較同一因素在不同水平下所對應(yīng)酶活的大小,從而得到的各個因素的最優(yōu)水平組合為A3B2C1,即這三個因素的添加量為葡萄糖2%,牛肉膏0.3%,NH4Cl 0.1%。
采用實驗所得的最佳培養(yǎng)基配方,即葡萄糖2%,牛肉膏0.3%,NH4Cl 0.1%,KH2PO40.05%,MgSO40.05%,經(jīng)過三次平行實驗得到的酶活U分別為12.13、13.17、11.48u/mL,說明該菌株在上述培養(yǎng)基組合下產(chǎn)酶較穩(wěn)定。由此確定以上優(yōu)化的培養(yǎng)基配方為最佳產(chǎn)酶培養(yǎng)基配方。
3 結(jié)論
3.1 不同有機氮源在一定程度上均能促進(jìn)酶的合成,以牛肉膏為最佳。
3.2 添加以相同摩爾數(shù)氮元素的不同無機氮源時,NH4Cl是最佳無機氮源。
3.3 MgSO4和KH2PO4對產(chǎn)酶的影響都不是很大,MgSO4和KH2PO4添加量均為0.05%時,產(chǎn)酶效果最好。
3.4 正交實驗中以葡萄糖、牛肉膏和NH4Cl作為優(yōu)化菌株產(chǎn)內(nèi)切β-1,3-葡聚糖酶的實驗因素,發(fā)現(xiàn)對產(chǎn)酶的影響主次因素是:葡萄糖gt;牛肉膏gt;NH4Cl,從而得到裂褶菌產(chǎn)內(nèi)切β-1,3-葡聚糖酶的最佳培養(yǎng)基配方為:葡萄糖2%,牛肉膏0.3%,NH4Cl 0.1%,KH2PO40.05%,MgSO40.05%。
[1]李兆蘭.裂褶多糖的結(jié)構(gòu)研究[J].南京大學(xué)學(xué)報:自然科學(xué)版,1994,30(3):482-486.
[2]Kikumoto S,Yamamoto O,Komatsu N.Method of produing neoschizophyllan having novel pharmacological activity[J].United States Patent,1978(4):661.
[3]Prokop A,Rapp P,wanger F.Production purification and characterization of an extracellular endo-β-1,3-glucanase from amonokaryon of schizophyllum commune ATCC 38548 defective in exo-β-1,3-glucanase formation[J].Can J Microbiol,1995,40:18-23.
[4]Chernin L,Chet I.Microbial enzymes in the biocontrol of plant pathogens and pests.In:Dick R P and Burns R G eds.,Enzyme in the Environment[J].New Youk:Marcel Dekker,2002: 171-225.
[5]Chet I,Chernin L.Biocontrol,microbiol agents in soil// Bitton G ed,Encyclopedia of environmental microbiology[J].New York:John Willey and Son Inc,2002:450-465.
[6]鄭文科,喬長晟,郝利民,等.四種生長因子對裂褶菌胞外多糖產(chǎn)量影響的優(yōu)化研究[J].食品工業(yè)科技,2009,30(5):181-183,187.
[7]張龍翔.生化實驗方法和技術(shù)[M].北京:人民教育出版社,1982.
[8]Jianguang Luo,Jun Liu,Chunling Ke.Optimization of medium composition forthe production ofexopolysaccharidesfrom Phellinus baumii Pilát in submerged culture and the immunostimulating activity of exopolysaccharides[J].Carbohydrate Polymers,2009,78(3):409-415.
[9]Katarína Kolenová, Mária Vr?anská.Purification and characterization of two minor endo-β-1,4-xylanases of Schizophyllum commune[J].Enzyme and Microbial Technology,2005,36(7):903-910.
[10]S G Jonathan,I O Fasidi.Studies on phytohormones,vitamins and mineral element requirements of Lentinus subnudus(Berk)and schizophyllum commune(Fr.Ex.Fr)from Nigeria[J].Food Chemistry,2001,75(3):303-307.
Study on the optimization of medium composition of endo-β-1,3-glucanase producing by Schizophyllum commune Fr.
CHANG Xiao-jie,ZHENG Bi-sheng*,ZHAO Xin
(College of Light Industry and Food Sciences,South China University of Technology,Guangzhou 510640,China)
The constant shaking was used to culture Schizophyllum commune Fr.,and the different single-factor parameters of carbon,nitrogen and various salt forming the medium of endo-β-1,3-glucanase producing by Schizophyllum commune Fr.was studied.At the same time,the four factors and three level orthogonal experimental was used to analyze the main factor.Finally,determining the optimum medium composition:glucose 2%,beef extract 0.3%,NH4Cl 0.1%,KH2PO40.05%,MgSO40.05%.
Schizophyllum commune Fr.;endo-β-1,3-glucanase;medium composition
TS201.3
A
1002-0306(2012)05-0167-04
2011-03-09 *通訊聯(lián)系人
暢曉潔(1987-),女,碩士生,研究方向:糖類分離提純新方法新技術(shù)。
