黔南晚石炭世生物灰泥丘生長發育特征研究
2012-11-14 02:38:48屠錫艷劉永華張曉光
中國非金屬礦工業導刊 2012年6期
關鍵詞:生物
屠錫艷,李 明,劉永華,李 芃,張曉光
(中國建筑材料工業地質勘查中心遼寧總隊,遼寧 沈陽 110004)
1 引言
生物礁研究一直是地質科學領域中十分重要的課題。盡管關于生物礁的概念說法不一,但總體認識是:生物礁是由各種生物建造而成的碳酸鹽巖隆,具有原地形成的各種生物結構,代表著生物原始的生存位置,反映生物原始的生態格局[1-3]。正是由于生物礁的這一獨特的生物建造特點,讓我們可以借助對生物礁的研究,來恢復其地史時期中生物群落組成、結構、生長發育過程及古生態特征。生物礁是開展沉積學、古生物學、古生態學等學科研究的理想場所。生物礁作為獨特的生物建造,其礁內各種孔隙、空洞非常發育,是石油和天然氣理想的儲集場所。自上世紀20年代,由于在生物礁中發現了大量的油氣,更加大了人們對生物礁的研究力度,另外生物礁與層控礦床也有密切關系,因此,生物礁研究得到普遍關注和重視。
從各地史時期生物礁的研究現狀來看,石炭紀生物礁的研究一直比較薄弱,特別是我國更是如此。多數學者認為石炭紀是地史時期的造礁低潮期,其礁體數量少,類型單一。但是,近幾年以鞏恩普教授為代表的學者對于石炭紀生物礁的研究取得了突破,證實了石炭紀存在大量不同類型的生物礁。
通過對工作區的野外和室內工作,基本查明了黔南晚石炭世生物灰泥丘生長發育特征,通過系統總結和分析,并且對比Waulsortian相研究,筆者認為晚石炭世生物灰泥丘并不是Waulsortian相,這對中國其他地區廣泛開展石炭紀生物灰泥丘研究具有重要指導意義,有助于石炭紀生物礁研究工作的深入。
2 區域地質特征
黔南地區大地構造位置歸屬于滇黔桂盆地,又稱南盤江盆地(圖1)。該盆地作為揚子板塊西南被動陸緣的一部分,從泥盆紀開始出現裂谷化成因的裂陷槽,因大陸拉伸,在南盤江盆地中出現塹壘相間的構造格局[1]。石炭紀時,該盆地與特提斯海盆鄰接,基底活動性強。由于深部熱膨脹引起以南北方向為主的引張力作用,造成地殼微型擴張,形成一系列隆、凹相間構造格局。在擴張過程中,處于固結較早、剛性較強的構造高位發育成碳酸鹽臺地,而拉張下陷部位則演化成臺間深水盆地。隨裂陷加深,構成了淺水補償型碳酸鹽臺地與半深水非補償型臺溝相間的古構造和巖相展布格局[2]。
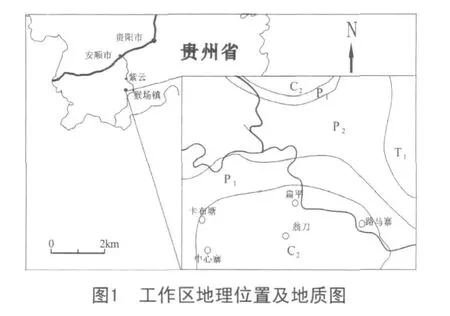
黔南晚石炭世海域進一步擴大,淺水碳酸鹽臺地范圍擴大并連成一片,臺盆相已收縮成孤立的狹長狀[3]。沉積巖石類型為獨山—威寧分區淺水臺地相淺色碳酸鹽巖,主要有淺灰色—灰色中厚層至塊狀生物碎屑灰巖、泥晶灰巖、微晶灰巖和各種礁灰巖,含有豐富的底棲生物。
3 泥丘相地層生物地層學研究
3.1 泥丘相剖面
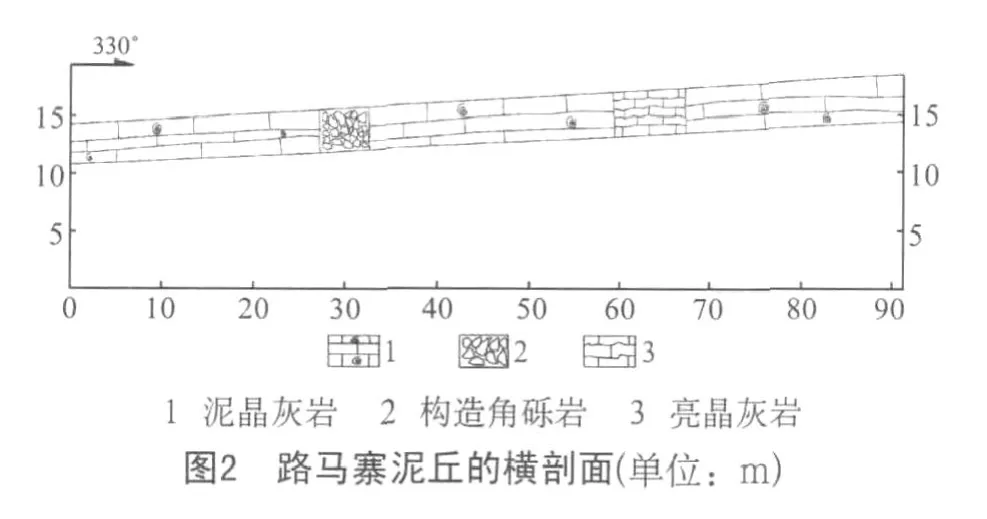
路馬寨泥丘橫剖面(見圖2)長90m,主要為泥晶膠結的粒泥灰巖。地層中生物量很少,偶見海百合莖及有孔蟲,但蜓碎屑相對較發育。在泥丘的橫剖面可以看出,路馬寨灰泥丘在橫向上巖性變化小,只發育一段構造角礫巖和一段亮晶灰巖,這兩段巖性差異主要來源于構造運動以及成巖后期的亮晶方解石充填孔隙、重結晶作用。
(1)泥丘下伏巖層——蜓生物碎屑灰巖。該層灰巖中發育大量的蜓碎屑,偶見海百合莖、有孔蟲、單體珊瑚以及少量腕足動物碎片。
(2)生物灰泥丘,外貌均呈穹形隆起。丘核均為泥晶灰巖,不顯層理,均呈塊狀,且藻絲發育,偶見團塊狀粒狀體(推測為藻顆粒)。丘頂與丘核無明顯差異。丘兩翼均為生物碎屑灰巖,主要為蜓碎屑灰巖。
(3)灰泥丘的上覆巖層——含蜓的亮晶生物碎屑灰巖。該層灰巖中發育大量的蜓碎屑、海百合莖、腕足動物、單體珊瑚以及其他生物。巖石以亮晶膠結為主,屬高能環境產物。
3.2 礁相地層生物群特征
從上述各礁相地層剖面看,工作區廣泛發育一套灰色碳酸鹽巖,其中礁、丘、灘分布普遍,代表了黔南晚石炭世礁相地層典型的巖性特點。在地層中,生物繁盛,其中不乏多種造礁生物,為黔南晚石炭世生物礁的形成和發展奠定了基礎。
蜓是石炭系和二疊系劃分、對比地層的重要標準化石,在本工作區所代表的黔南晚石炭世地層中,它是地層劃分、對比的主要標準。
工作區產出生物灰泥丘地層中,蜓類化石相對其它生物化石,則較為豐富,從總的蜓類生物群特征來看,發育灰泥丘的地層應該是晚石炭世晚期地層。
3.3 泥丘中的其他生物
工作區泥丘相地層中除上述生物外,還發育其他一些生物。它們有的具有造丘作用,有的具有附丘作用,還有的具有毀丘作用,以下對于有關各種生物的特征及其在泥丘中的作用進行簡單論述。
(1)有孔蟲。有孔蟲在灰泥丘中非常少見,且種屬單一,不起造丘作用,呈單體存在。
(2)海百合莖。泥丘相地層中分布一些海百合莖,個體較小,呈小的群體產出。
(3)藻類。泥丘相地層中存在的藻類較單一,具體屬種鑒定比較困難,絕大多數缺少內部結構特征。經觀察研究推測其應為蠕孔藻。
(4)藍細菌。在礁相地層中起粘結作用,形成粘結巖,也是形成規模不同生物泥丘的造丘生物。
4 生物灰泥丘特征
黔南晚石炭世生物灰泥丘為較深水的,通過微生物粘結作用、造巖作用形成的碳酸鹽建隆,其基本特征見圖3。
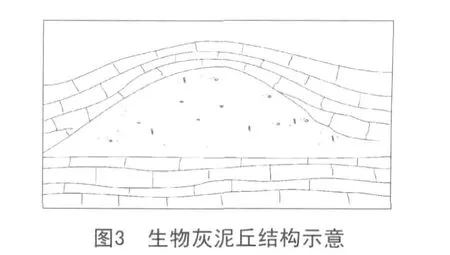
(1)組構特征。外貌均呈穹形隆起,抗浪構造明顯而典型。丘核均為泥晶灰巖,不顯層理,且藻絲發育,偶見團塊狀粒狀體(推測為藻顆粒)。丘頂與丘核無明顯差異。丘兩翼均為生物碎屑灰巖,主要為蜓碎屑灰巖。野外露頭和顯微鏡下觀察均可見晶洞,孔內為亮晶方解石充填。
(2)巖石構成。其巖石構成主要為粒泥灰巖,及少量藻凝塊巖,但骨骸顆粒較低而泥晶基質含量較高,表明為較深水的,水動力條件較弱的沉積環境。
(3)古生物化石及其古生態。同層位的灘相沉積富含多種生物,腕足類、蜓、非蜓有孔蟲、海百合、苔蘚蟲以及珊瑚類,與此形成鮮明對比的是,灰泥丘的生物豐度很低且種屬單調。偶見單體珊瑚化石,薄片中見少量海百合、有孔蟲以及蜓類化石。
丘翼主要由亮晶膠結生物碎屑灰巖組成,砂礫屑成分為泥晶方解石,普遍富含藻絲;重結晶作用強,鏡下常見重結晶方解石呈嵌晶狀,將砂屑、生物屑鑲嵌其中。亮晶方解石明顯可分為兩期:第一期為沿孔壁生長的馬牙狀方解石;第二期呈中一粗粒方解石充填孔隙中,在粒狀方解石晶間孔中偶見殘余瀝青。由于強烈的膠結作用和重結晶作用,殘余原生孔隙基本全被兩期方解石膠結物填充。次生孔隙,以砂屑,生物碎屑粒內溶孔和粒間溶孔、方解石膠結物晶間溶孔為主,孔隙度很低。在灰泥丘丘頂與丘核分布有藻絲。丘頂部與圍巖接觸界限明確。
灰泥丘的生長發育過程:由于凝塊石灰泥丘內無宏觀的造架生物,因而長期以來對其成因有多種認識。灰泥丘夾于灰色—灰黑色含化石豐富的灰巖之中。經過觀察以及較為深入的研究,將工作區灰泥丘的生長發育過程分為三個階段。
第一階段:基底的建立(發育期)。
當水深、水溫、水動力條件等控制灰泥丘生長的因素達到適合其生長的標準時,藍細菌等微生物開始在較為平坦的碎屑灘上大量繁殖,在生長過程中一方面粘結灰泥,另一方面鈣化的微生物參與造巖,同時也是灰泥的提供者。在各種微生物的共同作用下,形成了具早期固結特點簡單的基底建造(見圖4)。
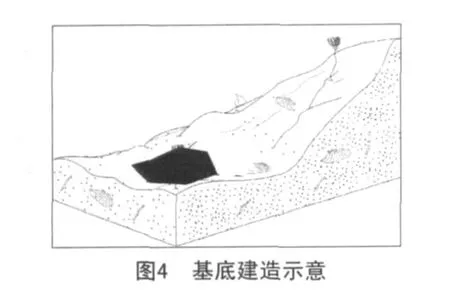
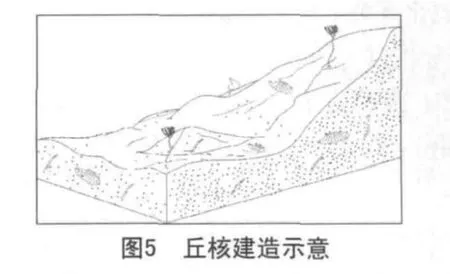
第二階段:丘核的建立(繁盛期)。
在簡單的基底建造之上,藍細菌等微生物進一步繁殖生長,形成了生物灰泥丘的次級生長構造。在開放的格架孔中,可充填灰泥和細小生物碎屑等沉積物,而在遮閉的格架孔中,空洞可被亮晶方解石充填,或者可繼續發育成為凝塊石。藍細菌群落在這種比較安靜的水體中生活,灰泥含量較高。群落內藍細菌是優勢生物,多為絲狀體,在其生長過程中,它以粘結作用和粘結包覆作用來粘結或吸附灰泥顆粒一同沉積下來,進而完成泥丘建造(見圖5)。
第三階段:丘頂的建立(衰退期)。
灰泥丘的終結有兩點原因(見圖6)。
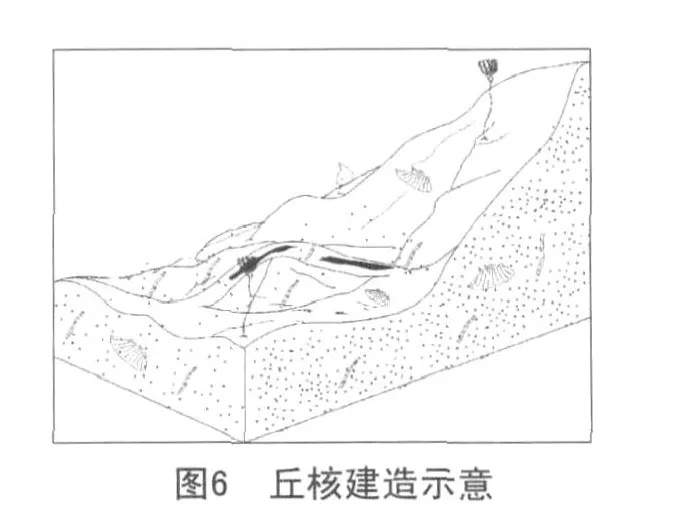
(1)隨著海平面的下降,水動力條件的增強,大量的造礁生物大量繁殖,汲取了大部分的養料,微生物在這種動蕩的沉積環境中很難生存,鈣化停止進而被在水動力作用較強的條件下沉淀的顆粒質灰巖掩埋—灰泥丘終結。
(2)隨著海平面的上升,水動力條件的減弱,藍細菌賴以生存的陽光很難到達海底,水溫也降低。由于缺少陽光以及溫度的影響,藍細菌(自養)不能完成正常的新陳代謝而走向死亡—灰泥丘終結。
綜上所述,微生物的沉積、造巖造丘作用明顯,灰泥丘的發育主要是微生物生命活動的結果。
5 黔南生物灰泥丘與世界Waulortian丘對比研究
(1)規模對比。就丘體的規模而言,黔南生物灰泥丘的厚度和直徑從幾米到幾十米且成群出現,一般規模較小;而Waulsortian丘的厚度和直徑從一米到上百米,成群出現或橫向聯結形成上千米長的大泥灘,一般規模較大。
(2)巖相對比。就巖相方面比較,黔南生物灰泥丘的巖相組成為粒泥灰巖,屬弱水動力條件的沉積環境;Waulsortian丘的巖相組成粒泥灰巖、泥粒灰巖、顆粒灰巖、漂浮巖,巖石顆粒大于黔南生物灰泥丘,說明Waulsortian丘生成環境的水動力條件略強于黔南生物灰泥丘的生成水動力條件。
(3)造丘生物對比。黔南生物灰泥丘的造丘生物為自養細菌,其生活環境應為富氧、透光的環境;Waulsortian丘的造丘生物為異養細菌,其生活環境應為富養、不透光或亞透光的環境。說明Waulsortian丘的生成水深要大于黔南生物灰泥丘的生成水深。
(4)丘內常見生物對比。黔南生物灰泥丘內常見的有孔蟲、海百合莖、苔蘚蟲以及蜓不起造丘作用,只是懸浮于灰泥基質中的一種附屬品。且這些化石的個體都很小,呈小的單體,內部結構簡單;Waulsortian丘的常見的珊瑚、鈣質海綿、有孔蟲、腕足類、投足類、海百合莖以及苔蘚蟲不起造丘作用,只是懸浮于灰泥基質中的一種附屬品。大部分珊瑚和海綿呈指狀或層狀,藉以適應泥質環境。此外,這些珊瑚化石的個體都很小,呈小的單體,隔壁發育,間室構造簡單,地層跨度長。但Waulsortian丘這些生物量很大,遠大于在黔南灰泥丘中的含量。在通過對比說明,黔南生物灰泥丘和Waulsortian丘的生存環境均不適合其它生物的生長。
(5)成丘地層對比。黔南生物灰泥丘通過蜓類鑒定,確定其生成地層為晚石炭世地層;而Waulsortian丘的生成地層為早石炭世(杜內期—韋憲期)的地層。Waulsortian丘早于黔南生物灰泥丘形成。
(6)晶洞對比。黔南生物灰泥丘和Waulsortian丘都遍存在洞穴(層狀和條帶狀孔洞),代表軟泥中的流體排泄構造,部分或全部充以微亮晶或亮晶方解石膠結物。
(7)丘體結構對比。黔南生物灰泥丘和Waulsortian丘都具備丘頂、兩翼、丘核的結構,但Waulsortian丘的三結構具有明顯的巖相差異。①海百合的停機坪構造(海百合丘頂):該停機坪有4個優勢相,即顆粒灰巖、粒泥灰巖、泥粒灰巖及漂浮巖相,孔隙被方解石充填。該相缺乏其他類型的淺成灰巖,整個構造近乎水平;②苔蘚蟲丘核(丘核):巖石為粗集粒灰巖和漂浮巖,其中含有具有窗孔構造的苔蘚蟲及少量的海百合莖。丘核主要為泥晶膠結,含有海百合莖、窗孔苔蘚蟲、有孔蟲、腕足類等生物,生物種屬單一,個體小且數量少,地層跨度長;③兩翼(丘翼):其巖石為粒泥灰巖、泥粒灰巖及漂浮巖,含有少量的海百合及介形蟲,具有纖細的泥質迭片結構,坡角為30~50°;而對于黔南生物灰泥丘的三結構不具備明顯的巖相差異,丘頂和丘核主要為泥晶膠結的粒泥灰巖和微晶灰巖,兩翼為泥晶膠結的蜓碎屑灰巖。
(8)成巖作用對比。Waulsortian丘的成巖作用包括:方解石膠結、機械壓實作用、白云巖化、硅化以及碳氫巖化。丘核巖相包括水底束狀的方解石、葉片狀/凌柱狀方解石團塊具有明亮的光譜特征,孔隙被亮晶方解石充填。海百合停機坪構造包含共軸膠結物與海百合有序膠結,即4種白云巖化的類型被認知:①局部置換;②化學壓實;③白云巖浸透;④氨基轉換。
對于黔南生物灰泥丘的成巖作用較為簡單,主要為重結晶、膠結作用。白云巖化、硅化作用在泥丘成巖過程中并未起主要作用,只是在后期的構造裂隙中偶然發生且不強烈。證明泥丘被埋藏后受熱液影響較小。
(9)沉積環境對比。根據以上分析確定黔南生物灰泥丘的沉積環境為較深水、富氧、弱水動力條件的透光帶斜坡上;而Waulsortian丘的沉積環境深水、貧氧、弱水動力條件的亞透光帶或不透光斜坡上。
綜上所述,黔南地區所發育的灰泥丘與Waulsortian丘具有明顯的差異,因此確定黔南地區所發育的灰泥丘并非Waulsortian丘,而是一種特殊的石炭紀灰泥建造。
6 結論
(1)灰泥丘的類型單一,規模小。
(2)造礁生物為自養的藍細菌微生物。
(3)丘內生物種類單一,內部結構簡單。
(4)查明了生物灰泥丘的建造過程。
(5)確定生物灰泥丘的生成環境為較深水、富氧、弱水動力條件的透光帶斜坡上。
(6)確定了生物灰泥丘的產丘層位為上石炭統上部地層,即馬平組下部地層。
(7)與Waulsortian相對比研究表明,黔南地區所發育的灰泥丘與Waulsortian丘具有明顯的差異,因此確定黔南地區所發育的灰泥丘并非Waulsortian相。
中國南方晚石炭世生物灰泥丘研究并不多,除本工作區之外,其他地區還沒有開展相關研究。所以生物灰泥丘的研究對全面認識晚石炭世生物礁的基本發育特征具有重要意義。
[1]趙煥庭,宋朝景,朱袁智.南沙群島“危險地帶”腹地珊瑚礁的地貌與現代沉積特征[J].第四紀研究,1992(4):368-377.
[2]趙美霞,余克服,張喬民.珊瑚礁區的生物多樣性及其生態功能[J].生態學報,2006,26(1):186-194.
[3]楊惠民,劉炳溫,鄧宗淮.滇黔桂海相碳酸鹽巖地區最佳油氣保存單元的評價與選擇[M].貴陽:貴州科技出版社,1999,44-47.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
