自噬在熊果酸誘導(dǎo)的人臍靜脈內(nèi)皮細胞損傷中的作用*
2012-11-06 06:06:23畢娟娟葉秀峰
中國病理生理雜志 2012年4期
關(guān)鍵詞:檢測
畢娟娟, 何 林, 余 音, 郭 倩, 葉秀峰,2△
(重慶醫(yī)科大學(xué) 1神經(jīng)科學(xué)研究中心,2病理教研室,重慶 400016)
1000-4718(2012)04-0601-07
2011-11-18
2012-02-21
重慶市教委科技研究項目基金資助項目(No.KJ08032);重慶市科技攻關(guān)計劃項目(No.CSCT,2008AB5118)
△通訊作者 Tel:023-68485789; E-mail:Yxf1960@126.com
自噬在熊果酸誘導(dǎo)的人臍靜脈內(nèi)皮細胞損傷中的作用*
畢娟娟1, 何 林1, 余 音1, 郭 倩1, 葉秀峰1,2△
(重慶醫(yī)科大學(xué)1神經(jīng)科學(xué)研究中心,2病理教研室,重慶 400016)
目的研究自噬在熊果酸誘導(dǎo)的人臍靜脈內(nèi)皮細胞(human umbilical vein endothelial cells,HUVECs)損傷中的作用。方法體外培養(yǎng)HUVECs,運用不同濃度的熊果酸處理36 h,采用MTT法檢測HUVECs增殖率的變化;透射電鏡觀察細胞超微結(jié)構(gòu)的改變;單丹磺酰尸胺(MDC)熒光染色觀察熊果酸對HUVECs自噬表達水平的影響;Western blotting檢測微管相關(guān)蛋白輕鏈3(microtubule-associated protein light chain 3, LC3)及自噬相關(guān)蛋白Beclin-1的變化;RT-PCR檢測LC3及Beclin-1 mRNA的變化;流式細胞術(shù)檢測自噬抑制劑3-甲基腺嘌呤(3-methyladenine, 3-MA)抑制自噬前后經(jīng)熊果酸處理的HUVECs凋亡率的變化。結(jié)果熊果酸抑制HUVECs增殖并呈劑量依賴性;熊果酸可誘導(dǎo)HUVECs產(chǎn)生自噬:透射電鏡和MDC染色均顯示HUVECs經(jīng)熊果酸處理后自噬囊泡明顯增加且自噬在基因及蛋白水平的表達均增加;3-MA與熊果酸聯(lián)合作用于HUVECs時抑制了熊果酸誘導(dǎo)的HUVECs自噬空泡的積聚并且可明顯促進HUVECs凋亡。結(jié)論熊果酸抑制HUVECs增殖,并誘導(dǎo)其發(fā)生自噬,自噬在此過程中對HUVECs起保護作用。自噬抑制劑3-MA能夠增強熊果酸誘導(dǎo)的HUVECs的增殖抑制作用并且促進HUVECs發(fā)生凋亡。
熊果酸; 內(nèi)皮細胞; 自吞噬作用; 細胞凋亡
病理性血管生成具有持續(xù)性、無控性,其在腫瘤的演化過程中起著重要的作用。腫瘤新生血管對其提供氧氣及營養(yǎng)物質(zhì)則作為腫瘤侵襲轉(zhuǎn)移的首要步驟,因此,目前治療腫瘤的焦點已匯集于抑制腫瘤血管生成[1]。
自噬是廣泛存在于真核細胞內(nèi)的一種溶酶體依賴性的降解途徑。眾多研究表明自噬在疾病發(fā)生及發(fā)展過程中具有促進和抑制的雙重作用:自噬能夠在饑餓及外界刺激下回收大分子以及清除受損的線粒體及其它細胞器,為細胞提供能量及蛋白質(zhì)實現(xiàn)再循環(huán)以維持細胞生存。然而,自噬也可以引起細胞生長阻滯,以及一種與過量自噬空泡形成有關(guān)的自噬性細胞死亡[2]。
熊果酸(ursolic acid,UA)又稱烏索酸或烏蘇酸,一種來源于傳統(tǒng)中草藥的提取物,屬五環(huán)三萜類化合物。它廣泛存在于自然界中,并具有多種生物學(xué)效應(yīng),比如抗腫瘤、抗血管生成、抗氧化、抗抑郁、保肝、降血脂、抗病毒及抗菌作用等,且毒副作用較小[3]。 Sohn 等[4]雞胚胎絨毛膜實驗(chicken chorioallantoicmembrane assays,CAM)研究顯示,熊果酸可較強地抑制血管生成,但其作用機制目前尚未明確。我們之前研究表明熊果酸可通過抑制ERK信號通路抑制血管內(nèi)皮細胞增殖[5]。目前,關(guān)于自噬在熊果酸誘導(dǎo)血管內(nèi)皮細胞損傷中的作用報道較少,本文利用自噬抑制劑3-甲基腺嘌呤(3-methyladenine,3-MA)協(xié)同作用,初步探討自噬在熊果酸誘導(dǎo)內(nèi)皮細胞損傷中的作用。
材 料 和 方 法
1細胞
人臍靜脈血管內(nèi)皮細胞(human umbilical vein endothelial cells,HUVECs)株由重慶醫(yī)科大學(xué)病理教研室提供。
2主要試劑
熊果酸(純度>98%)購自陜西旭煌植物科技發(fā)展有限公司;DMEM/high glucose購自HyClone;小牛血清購自Gibco;3-MA、單丹磺酰尸胺(monodansylcadaverine,MDC)、Western Blotting Luminol Reagent和兔抗人微管相關(guān)蛋白輕鏈3(microtubule-associated protein light chain 3,LC3)多克隆抗體購自Sigma;MTT購自北京鼎國昌盛生物技術(shù)有限責(zé)任公司;PRO-PREPTM蛋白提取液購自北京賽百盛基因技術(shù)有限公司;Bradford蛋白質(zhì)定量試劑盒購自上海生工生物工程技術(shù)服務(wù)有限公司;兔抗人Beclin-1多克隆抗體購自Santa Cruz;小鼠抗人β-actin、HRP標記的山羊抗兔IgG和山羊抗鼠IgG購自北京中杉金橋生物技術(shù)有限公司;DNA marker 購自Bioflux;RNA提取試劑盒購自TaKaRa;逆轉(zhuǎn)錄試劑盒購自北京百泰克生物技術(shù)有限公司;Taq酶購自SinoBio。
3細胞培養(yǎng)
HUVECs置于含15%小牛血清的DMEM/high glucose培養(yǎng)液中,于37 ℃、5%CO2培養(yǎng)箱中常規(guī)培養(yǎng),取處于對數(shù)生長期的細胞進行實驗。
4MTT法檢測熊果酸對HUVECs增殖的影響
取對數(shù)生長期的HUVECs,以5 000 cells/well的密度接種于96 孔板,每孔加200 μL培養(yǎng)基,于孵箱中培養(yǎng)24 h,將細胞分為熊果酸組(分別加入5、10、20、40、80 μmol/L熊果酸);3-MA+熊果酸處理組(先用2 mmol/L 3-MA預(yù)處理細胞,1 h后加入不同濃度熊果酸);溶劑對照組(只加相應(yīng)濃度DMSO不加熊果酸)和空白調(diào)零組(只含培養(yǎng)液,無細胞),每組設(shè)5 個復(fù)孔,重復(fù)3 次。作用36 h后,每孔加入MTT(5 g/ L)20 μL,繼續(xù)培養(yǎng)4 h,棄去孔內(nèi)液體,每孔加入150 μL DMSO,振蕩10 min,酶聯(lián)免疫測定儀選擇波長570 nm空白孔調(diào)零,測定各孔吸光度(A)值并計算細胞增殖抑制率和半數(shù)抑制濃度(IC50)。細胞增殖抑制率(%)=[ (溶劑對照組A值-處理組A值)/溶劑對照組A值] ×100 %。
5透射電鏡法檢測熊果酸對HUVECs細胞超微結(jié)構(gòu)的影響
取對數(shù)生長期HUVECs,以每瓶5×105的密度接種于100 mL培養(yǎng)瓶中,每瓶加5 mL培養(yǎng)液。熊果酸(20 μmol/L)給藥 36 h后常規(guī)收集細胞,正常對照組用完全培養(yǎng)基培養(yǎng)36 h。PBS離心洗滌1次,1 000 r/min離心10 min, 4%戊二醛固定,于 4 ℃保存,送重慶醫(yī)科大學(xué)生命科學(xué)院電鏡室固定、切片、攝片。
6MDC熒光染色檢測細胞內(nèi)自噬空泡的變化
MDC是自噬空泡的標志物,通過MDC染色可以觀察判斷細胞自噬過程的發(fā)生[6]。取對數(shù)生長期HUVECs,以2×104/well的密度接種于已放置小蓋玻片的24孔板中,每孔加1 mL培養(yǎng)液,培養(yǎng)24 h后加入終濃度為20 μmol/L熊果酸,另設(shè)3-MA(2 mmol/L)+熊果酸(20 μmol/L)處理組和正常對照組。置37 ℃、5% CO2細胞培養(yǎng)箱中繼續(xù)培養(yǎng)。36 h后加入終濃度50 μmol/L MDC,37 ℃孵育60 min,PBS液漂洗5 min×3次。4%多聚甲醛室溫固定細胞15 min,PBS液漂洗5 min×3次,熒光顯微鏡觀察細胞自噬空泡的變化并統(tǒng)計熒光染色陽性細胞百分比。
7Westernblotting檢測LC3和Beclin-1蛋白的表達
取對數(shù)生長期的HUVECs以每瓶5×105的密度接種于100 mL培養(yǎng)瓶中,每瓶加5 mL培養(yǎng)液,24 h后按如下分組進行處理: 20 μmol/L熊果酸分別處理0、12、24、36、48 h。取各組細胞,提取總蛋白,Bradford法測定蛋白濃度。經(jīng)SDS-PAGE分離后,將蛋白轉(zhuǎn)移至PVDF膜上,5 kg/L脫脂奶粉37℃封閉1 h;加入5%脫脂奶粉稀釋的兔抗人LC3(1∶1 000 稀釋)或兔抗人Beclin-1(1∶200稀釋)多克隆抗體或小鼠抗人β-actin 單克隆抗體(1∶500 稀釋),4 ℃孵育過夜;TBST 洗膜后,加入5%脫脂奶粉稀釋、HRP標記的山羊抗兔或山羊抗鼠IgG(1∶1 000 稀釋),37 ℃孵育2 h;ECL顯色,采用ChemiDocXRS化學(xué)發(fā)光成像系統(tǒng)進行曝光采圖。用Quantity One 4.6.2 軟件分析,比較條帶的面積和灰度值,以β-actin為內(nèi)參照,進行半定量。
8RT-PCR檢測自噬相關(guān)蛋白LC3和Beclin-1mRNA的表達
8.1分組 取對數(shù)生長期的HUVECs以每瓶5×105的密度接種于100 mL培養(yǎng)瓶中,每瓶加5 mL培養(yǎng)液,24 h后按如下分組進行處理:(1)20 μmol/L熊果酸分別處理0、12、24、36、48 h;(2)熊果酸(20 μmol/L)組、3-MA(2 mmol/L)+熊果酸(20 μmol/L)組和正常對照組(正常培養(yǎng)細胞)。
8.2引物設(shè)計及合成 根據(jù)Gene中登錄的LC3及Beclin-1序列設(shè)計引物:LC3上游引物5′-TCCCACACCAAGTGCATGTCAG-3′,下游引物 5′-GCCAAGTGGACAGACGCATGC-3′,擴增片段長度為390 bp;Beclin-1上游5′-GGTGGCTTTCCTGGACTGTGTGC-3′,下游引物5′-TCAAAACTGACCAGGGCTGGCA-3′,擴增片段長度為465 bp;內(nèi)參照β-actin 上游引物5′-CCGCCAGCTCACCATGGATGAT-3′,下游引物5′-TCTCTTGCTCTGGGCCTCTG-3′,擴增片段長度為199 bp,由北京賽百盛公司合成。
8.3細胞總RNA 的提取 取各組細胞,PBS洗滌2次后加入Trizol提取液,以氯仿/異丙醇法抽提各組細胞總RNA,測定A260/A280比值。
8.4RT-PCR 以提取的細胞總RNA為模板,進行逆轉(zhuǎn)錄,以逆轉(zhuǎn)錄產(chǎn)物為模板進行PCR擴增,以β-actin作為內(nèi)參照。反應(yīng)體系:Taq酶12.5 μL、上、下游引物各1 μL ,cDNA2.5 μL,用滅菌蒸餾水補足體積至25 μL。LC3反應(yīng)條件:94 ℃ 3 min,94 ℃ 30 s,56 ℃ 45 s,72 ℃ 45 s,共35 個循環(huán),72 ℃ 5 min。Beclin-1反應(yīng)條件:94 ℃ 3 min,94 ℃ 30 s,56 ℃ 45 s,72 ℃ 45 s,共35 個循環(huán),72 ℃ 5 min。PCR產(chǎn)物經(jīng)2%瓊脂糖凝膠電泳鑒定并計算LC3、Beclin-1 mRNA/β-actin吸光度值。
9流式細胞術(shù)檢測細胞凋亡率
取對數(shù)生長期HUVECs,以每瓶5×105的密度接種于100 mL培養(yǎng)瓶中,每瓶加5 mL培養(yǎng)液。培養(yǎng)24 h后加入20 μmol/L熊果酸,另設(shè)3-MA(2 mmol/L) +熊果酸(20 μmol/L)處理組、正常對照組。繼續(xù)培養(yǎng)36 h后常規(guī)收集細胞, PBS液沖洗,1 000 r/min離心5 min×3次。送重慶醫(yī)科大學(xué)生命科學(xué)院annexin V-FITC和PI染色后流式細胞儀檢測細胞凋亡率。
10統(tǒng)計學(xué)處理
結(jié) 果
1熊果酸對HUVECs增殖的影響
不同濃度熊果酸(5、10、20、40、80 μmol/L)作用HUVECs 36 h后均有抑制作用,且呈明顯的劑量依賴性,不同濃度與對照組(0 μmol/L)之間的抑制率差異均有統(tǒng)計學(xué)意義(P<0.05),熊果酸處理HUVECs 36 h的IC50為20 μmol/L,以下相關(guān)實驗選用的藥物濃度為20 μmol/L,見圖1。


圖1不同濃度熊果酸對HUVECs增殖的抑制率
2熊果酸可誘導(dǎo)HUVECs產(chǎn)生自噬
2.1透射電鏡觀察熊果酸對HUVECs超微結(jié)構(gòu)的影響 正常對照組細胞核膜完整,染色質(zhì)結(jié)構(gòu)正常,胞漿內(nèi)各細胞器、細胞核形態(tài)和分布基本正常,未見明顯自噬空泡,溶酶體數(shù)目未見明顯增多。熊果酸(20 μmol/L)處理36 h后胞漿中可見大量空泡狀結(jié)構(gòu)及包含部分胞漿成分形成雙層膜結(jié)構(gòu)的自噬體,表明熊果酸可誘導(dǎo)HUVECs發(fā)生自噬,見圖2。
2.2Western blotting檢測LC3和Beclin-1蛋白表達情況 20 μmol/L熊果酸處理HUVECs后,結(jié)果顯示:隨著作用時間的延長LC3Ⅱ/LC3Ⅰ逐漸增高,除48 h外各時間組與對照組之間差異有統(tǒng)計學(xué)意義(P<0.05);隨著作用時間的延長Beclin-1蛋白表達增高,除12 h外各時間組與對照組之間差異有統(tǒng)計學(xué)意義(P<0.05),見圖3。

Figure 2. The effect of ursolic acid (UA) on ultrastructure of HUVECs. A:control; B: UA.
圖2熊果酸對HUVECs細胞超微結(jié)構(gòu)的影響
2.3RT-PCR檢測LC3和Beclin-1 mRNA表達情況 20 μmol/L熊果酸處理HUVECs后,結(jié)果顯示:隨著作用時間的延長LC3 mRNA表達逐漸增高,于36 h達高峰,各時間組與對照組之間差異有統(tǒng)計學(xué)意義(P<0.05);隨著作用時間的延長Beclin-1 mRNA表達增高,除12 h組外,各時間組與對照組之間差異有統(tǒng)計學(xué)意義(P<0.05),見圖4。
3抑制自噬增強了熊果酸對HUVECs的增殖抑制作用
3.1MDC熒光染色檢測細胞內(nèi)自噬空泡的變化 MDC染色結(jié)果顯示單獨應(yīng)用熊果酸作用于HUVECs時,細胞內(nèi)出現(xiàn)明顯自噬空泡積聚,陽性細胞率為(67.12±2.85)%,與正常對照組陽性細胞率[(5.37±1.34)%]相比有明顯差異,表明有自噬發(fā)生;熊果酸與3-MA聯(lián)合作用(24.07±0.60)%時,自噬空泡明顯減少但仍高于正常對照組,表明自噬被抑制,見圖5。

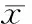
圖320μmol/L熊果酸處理不同時間HUVECs中LC3及Beclin-1蛋白的表達
3.2RT-PCR檢測3-MA對熊果酸誘導(dǎo)的HUVECs LC3和Beclin-1 mRNA表達的影響 20 μmol/L熊果酸單獨處理或2 mmol/L 3-MA與20 μmol/L熊果酸聯(lián)合處理的HUVECs中, LC3和Beclin-1 mRNA的轉(zhuǎn)錄水平明顯高于正常對照組,其中熊果酸單獨處理組高于3-MA與熊果酸聯(lián)合處理組,差異顯著(P<0.05),見圖6。
3.33-MA對熊果酸抑制HUVECs增殖的影響 MTT結(jié)果顯示:熊果酸單獨作用HUVECs或3-MA與熊果酸聯(lián)合作用對細胞增殖均有抑制作用,后者抑制作用更為顯著,其細胞存活率差異顯著(P<0.05);3-MA 與對照組相比,差異沒有統(tǒng)計學(xué)意義,見圖7。
4抑制自噬能促進熊果酸誘導(dǎo)的HUVECs凋亡的發(fā)生
與正常對照組[細胞凋亡率(3.34±0.32)%]相比,熊果酸單獨作用HUVECs 36 h,細胞凋亡率增加[ (5.21±0.22)%];3-MA和熊果酸聯(lián)合作用時與單獨應(yīng)用熊果酸組相比,細胞凋亡率明顯增加[(12.7±0.56)%],見圖8。

圖420μmol/L熊果酸處理不同時間HUVECs中LC3及Beclin-1mRNA的表達


圖5MDC染色檢測熊果酸和3-MA作用36hHUVECs自噬空泡的變化
討 論
腫瘤血管生成是導(dǎo)致腫瘤侵襲、轉(zhuǎn)移的重要因素。近年來通過抑制腫瘤血管生成進而抑制腫瘤生長和轉(zhuǎn)移已成為腫瘤研究的熱點。傳統(tǒng)抗腫瘤血管生成主要是通過抑制血管生成因子的產(chǎn)生、抑制配基與受體相互作用而抑制血管生成因子的活性、選擇性拮抗劑直接作用于血管生長因子受體,調(diào)節(jié)受體在內(nèi)皮細胞上的表達水平或者引入野生型受體與負性非功能受體與之競爭、抑制內(nèi)皮細胞增殖、遷移及分化成新生血管的能力、抑制或者中和具有降解基底膜和細胞外基質(zhì)酶的活性[7]。但由于大多數(shù)抗血管生成藥物具有不穩(wěn)定性、毒性較大且價格昂貴 ,因此臨床應(yīng)用受到限制。熊果酸作為一種高效低毒的天然藥物,可抑制多種腫瘤細胞的生長,具有廣闊應(yīng)用前景,有望成為一種新型抗腫瘤藥物[8-9]。已有報道熊果酸可通過多種途徑抑制血管內(nèi)皮細胞增殖[10-11]。
自噬在疾病進展中的角色隨疾病病程不斷發(fā)生變化,細胞受到外界刺激發(fā)生自噬時,胞漿內(nèi)出現(xiàn)大量游離的膜性結(jié)構(gòu),逐漸包繞受損傷細胞器和部分胞漿形成自噬體,之后再與溶酶體融合形成自噬溶酶體,降解其所包裹的內(nèi)容物,以實現(xiàn)細胞本身的代謝。自噬在很多情況下都可以被激活,當營養(yǎng)缺乏或者各種生存條件不足時細胞可通過自噬降解細胞內(nèi)物質(zhì)為細胞提供營養(yǎng)及能量實現(xiàn)再循環(huán)。因此,從這個意義上講自噬是細胞面對不良刺激時所發(fā)生的一種防御行為[12]。然而,自噬具有兩面性,一方面可通過降解有害物質(zhì)保護細胞以避免壞死和凋亡的危險,而過度的激活則會通過降解細胞器導(dǎo)致細胞死亡。研究表明:自噬在氧化低密度脂蛋白誘導(dǎo)內(nèi)皮細胞損傷中起一定的保護作用,抑制自噬會促進內(nèi)皮細胞發(fā)生凋亡,對細胞產(chǎn)生損傷作用[13];另有研究報道抑制自噬可促進Nara-H細胞發(fā)生凋亡[14]。熊果酸誘導(dǎo)內(nèi)皮細胞損傷過程中是否誘導(dǎo)自噬的發(fā)生,自噬在此過程中起保護作用還是抑制作用則成為本課題研究的重點。

圖63-MA對熊果酸誘導(dǎo)的LC3和Beclin-1mRNA表達的影響

圖73-MA對熊果酸誘導(dǎo)的HUVECs細胞生存率的影響
本研究通過MTT檢測證明熊果酸對HUVECs具有細胞毒作用,并呈明顯的劑量依賴性;透射電鏡下觀察到經(jīng)熊果酸處理的HUVECs胞漿中可見明顯的自噬體及自噬溶酶體,與文獻中描述的自噬過程出現(xiàn)的空泡一致[15]。Atg為自噬相關(guān)蛋白,LC3為Atg8在哺乳動物中的同源體。Atg12結(jié)合過程和LC3修飾過程在哺乳動物自噬泡形成過程中起著重要的作用。Atg12結(jié)合過程參與前自噬泡的形成,LC3具有2種形式:LC3-I 和LC3-II。LC3前體形成后,在 Atg7 的催化作用下發(fā)生一系列反應(yīng):首先加工成胞漿可溶性形式的 LC3-I,隨后,LC3-I 被活化并轉(zhuǎn)運至 Atg3,并被修飾成膜結(jié)合形式的LC3-II。LC3-II定位于前自噬體和自噬體,其為自噬體的標志分子。一旦自噬體與溶酶體融合,自噬體內(nèi)的LC3- II便被溶酶體中的水解酶降解。 LC3-II含量或 LC3-II/LC3-I 的比值與自噬泡的數(shù)量呈正相關(guān)[16]。因此,通過檢測細胞內(nèi)LC3-II 的含量變化,可以基本判斷是否發(fā)生自噬及其自噬是被誘導(dǎo)還是被抑制。本實驗Western blotting顯示熊果酸處理HUVECs后與對照組相比, LC3-II/LC3-I 的比值明顯增加。同時,RT-PCR檢測顯示HUVECs經(jīng)熊果酸處理后與對照組相比LC3 mRNA表達明顯增多。則從另一個角度說明熊果酸在抑制內(nèi)皮細胞增殖的過程中誘導(dǎo)自噬的發(fā)生。MDC熒光染色發(fā)現(xiàn)單純熊果酸作用于HUVECs后細胞內(nèi)出現(xiàn)大量自噬空泡積聚,而自噬特異性抑制劑3-MA與熊果酸聯(lián)合作用后細胞內(nèi)自噬空泡明顯減少,明顯抑制了自噬的發(fā)生。我們同時經(jīng)RT-PCR證實了這一點。研究發(fā)現(xiàn):3-MA能夠一定程度地抑制H2O2誘導(dǎo)的U251細胞發(fā)生自噬并且促進其發(fā)生凋亡[17]。本實驗也證實了這一點,我們通過MDC、RT-PCR、檢測發(fā)現(xiàn)3-MA與熊果酸聯(lián)合作用組自噬相關(guān)基因表達較熊果酸單獨處理組明顯降低,同時MTT法檢測顯示抑制自噬后熊果酸誘導(dǎo)的HUVECs增殖抑制明顯增強;且抑制自噬可以促進熊果酸誘導(dǎo)的HUVECs發(fā)生凋亡。

圖83-MA對熊果酸誘導(dǎo)的HUVECs凋亡的影響
綜上所述,熊果酸在抑制HUVECs增殖過程中可誘導(dǎo)自噬的發(fā)生,自噬在此過程中對HUVECs起保護作用。抑制自噬,可增強熊果酸對HUVECs的抑制作用且促進HUVECs發(fā)生凋亡。
[1] Kunimasa K, Ohta T, Tani H ,et al.Resveratrol derivative-rich melinjo(GnetumgnemonL.) seed extract suppresses multiple angiogenesis-related endothelial cell functions and tumor angiogenesis[J]. Mol Nutr Food Res, 2011,55(11):1730-1734.
[2] Wu W,Wei W, Ablimit M,et al.Responses of two insect cell lines to starvation: autophagy prevents them from undergoing apoptosis and necrosis, respectively[J].J Insect Physiol, 2011, 57(6):723-734.
[3] Kuttan G, Pratheeshkumar P, Manu KA,et al. Inhibition of tumor progression by naturally occurring terpenoids[J]. Pharm Biol, 2011,49(10):995-1007.
[4] Sohn KH,Lee HY,Chung HY,et al.Anti-angiogenic activity of triterpene acid[J]. Cancer Lett,1995,94(2):213-218.
[5] 李 偉,劉 瓊,何 林,等.熊果酸對血管內(nèi)皮細胞增殖的影響及其機制[J].中國生物制品學(xué)雜志,2011, 24(2):157-162.
[7] Carmeliet P, Jain RK. Principles and mechanisms of vessel normalization for cancer and other angiogenic diseases[J]. Nat Rev Drug Discov,2011,10(6):417-427.
[8] Shan JZ, Xuan YY, Ruan SQ,et al. Proliferation-inhibiting and apoptosis-inducing effects of ursolic acid and oleanolic acid on multi-drug resistance cancer cellsinvitro[J]. Chin J Integr Med, 2011,17(8):607-611.
[9] Kim KH, Seo HS, Choi HS,et al. Induction of apoptotic cell death by ursolic acid through mitochondrial death pathway and extrinsic death receptor pathway in MDA-MB-231 cells[J]. Arch Pharm Res, 2011,34(8):1363-1372.
[10] Messner B, Zeller I, Ploner C,et al.Ursolic acid causes DNA-damage, P53-mediated, mitochondria- and caspase-dependent human endothelial cell apoptosis, and accelerates atherosclerotic plaque formationinvivo[J]. Atherosclerosis, 2011,219(2):402-408.
[11] Kanjoormana M, Kuttan G. Antiangiogenic activity of ursolic acid[J]. Integr Cancer Ther,2010 ,9(2):224-235.
[12] Tan ML, Ooi JP, Ismail N,et al. Programmed cell death pathways and current antitumor targets[J]. Biochem Biophys Res Commun,2010 ,394(3):760-766.
[13] Zhang YL,Cao YJ,Zhang X,et al.The autophagy-lysosome pathway: a novel mechanism involvedin the processing of oxidized LDL in human vascular endothelial cells[J]. Biochem Biophys Res Commun,2010,394(2):377-382.
[14] Nakamura O, Hitora T, Akisue T,et al.Inhibition of induced autophagy increases apoptosis of Nara-H cells[J]. Int J Oncol, 2011,39(6):1545-1552.
[15] Zeng R, He J, Peng J,et al. The time-dependent autophagy protects against apoptosis with possible involvement of Sirt1 protein in multiple myeloma under nutrient depletion[J]. Ann Hematol, 2012, 91(3):407-417.
[16] Chaachouay H, Ohneseit P, Toulany M,et al. Autophagy contributes to resistance of tumor cells to ionizing radiation[J]. Radiother Oncol, 2011,99(3):287-292.
[17] 孔曉霞,張宏宇,鐘加滕,等. 自噬抑制劑3-甲基腺嘌呤在H2O2引起的神經(jīng)膠質(zhì)瘤U251細胞損傷中的作用[J].中國病理生理雜志, 2011,27(4):695-698.
Roleofautophagyinursolicacid-inducedinjuryofhumanumbilicalveinendothelialcells
BI Juan-juan1, HE Lin1, YU Yin1, GUO Qian1, YE Xiu-feng1,2
(1InstituteofNeuroscience,2DepartmentofPathology,ChongqingMedicalUniversity,Chongqing400016,China.E-mail:Yxf1960@126.com)
AIM: To investigate the role of autophagy in the injury of human umbilical vein endothelial cells (HUVECs) induced by ursolic acid (UA).METHODSHUVECs were culturedinvitrowith UA at various concentrations for 36 h and the proliferation inhibitory rate of HUVECs was determined by MTT method. The change of ultrastructure was observed under transmission electronic microscope (TEM). The autophagy was observed using fluorescent microscope by monodansylcadaverin (MDC) staining. The protein level and mRNA expression of microtubule-associated protein light chain 3(LC3) and Beclin-1 were detected by Western blotting and RT-PCR, respectively. Cell apoptotic rate was measured by flow cytometry analysis.RESULTSUA at various concentrations showed significantly dose-dependent inhibitory effect on the proliferation of HUVECs. Autophagy was induced in HUVECs treated with UA as detected by MDC staining and TEM. The protein level and mRNA expression of LC3 and Beclin-1 in HUVECs were significantly increased following the treatment with UA, which was also in a time-dependent manner. Compared with UA group, addition of 3-methyladenine(3-MA) inhibited the increase in autophagic vacuoles and exacerbated the apoptosis.CONCLUSIONAutophagy shows protective effect on the proliferation inhibition of HUVECs induced by UA and the proliferation inhibition can be enhanced by the autophagy inhibitor 3-MA. 3-MA may enhance the apoptotic rate of HUVECs induced by UA.
Ursolic acid; Endothelial cells; Autophagy; Apoptosis
R363
A
10.3969/j.issn.1000-4718.2012.04.005
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
