禽類MHCⅡ類分子β鏈信號肽保守性分析
2012-10-16 05:30:44宗文明
赤峰學(xué)院學(xué)報·自然科學(xué)版 2012年15期
關(guān)鍵詞:分析
宗文明,葉 紅
(1.安徽農(nóng)業(yè)大學(xué),安徽 合肥 230036;2.安徽省醫(yī)學(xué)科學(xué)研究院,安徽 合肥 230061)
禽類MHCⅡ類分子β鏈信號肽保守性分析
宗文明1,葉 紅2
(1.安徽農(nóng)業(yè)大學(xué),安徽 合肥 230036;2.安徽省醫(yī)學(xué)科學(xué)研究院,安徽 合肥 230061)
Ⅱ類主要組織相容性復(fù)合物(major histocompatibility complex,MHC)的β鏈顯示極為豐富的多態(tài)性.為了分析禽類MHCⅡ類分子β鏈信號肽是否呈現(xiàn)出多態(tài)性的分子生物學(xué)特征,利用NCBI網(wǎng)站查找到原雞等五種禽類編碼MHCⅡ類分子β鏈的mRNA,再用DNAStar軟件轉(zhuǎn)譯成氨基酸序列,然后用信號肽在線預(yù)測軟件SignalP 3.0 Server進行分析,得出信號肽的概率、信號肽的長度以及信號肽酶的酶切位點,發(fā)現(xiàn)這五種禽類MHCⅡ類分子β鏈信號肽的分子生物學(xué)特征有許多相似性,利用Mega4.1軟件進行信號肽氨基酸序列比對分析,揭示出禽類MHCⅡ類分子β鏈信號肽具有高度的保守性.
禽類;MHC;信號肽;保守性
為什么有些蛋白質(zhì)(如線粒體、葉綠體等的蛋白質(zhì))在細胞質(zhì)基質(zhì)中合成,而有些(如分泌性蛋白質(zhì)、膜蛋白等)在粗面內(nèi)質(zhì)網(wǎng)上合成,是什么指令確定蛋白質(zhì)在細胞內(nèi)的合成部位以致最終影響蛋白質(zhì)的定向轉(zhuǎn)位呢?1975年,G.Blobel和D.Sabatini在實驗的基礎(chǔ)上提出了信號肽假說,即蛋白質(zhì)N端存在一段信號肽,指導(dǎo)蛋白質(zhì)遷移到內(nèi)質(zhì)網(wǎng)上合成,蛋白質(zhì)合成結(jié)束前信號肽被信號肽酶切除[1].
隨著生物信息學(xué)技術(shù)的迅速發(fā)展,對蛋白的結(jié)構(gòu)和功能進行預(yù)測分析已成為生物信息學(xué)的重要組成部分,同時也為進一步實驗研究提供相應(yīng)的理論依據(jù),使實驗數(shù)據(jù)更具目的性及有效性.信號肽對于控制蛋白質(zhì)的分泌路徑和指導(dǎo)蛋白質(zhì)定位特定位置有著重要作用,全面系統(tǒng)地研究信號肽不僅有助于認識、分析和解釋各種生理和病理現(xiàn)象,而且在尋求基因診療的新藥領(lǐng)域信號肽已經(jīng)成為一個關(guān)鍵工具[2].MHCⅡ類分子是由α鏈和β鏈組成的異二聚體,屬膜蛋白,其中β鏈顯示極為豐富的多態(tài)性.本文利用NCBI網(wǎng)站查找到原雞等五種禽類的編碼MHCⅡ類分子β鏈蛋白的mRNA序列,運用信號肽在線預(yù)測軟件SignalP 3.0和DNAStar進行分析,探討禽類MHCⅡ類分子β鏈信號肽序列的分子生物學(xué)特征,以發(fā)現(xiàn)它們是否呈現(xiàn)出多態(tài)性.
1 材料
1.1 mRNA序列
來源于NCBI(National Center for Biotechnology Information)數(shù)據(jù)庫,網(wǎng)址為http://www.ncbi.nlm.nih.gov/.
1.2 序列編輯軟件
DNAStar,網(wǎng)址為 http://www.torrents.net/.
1.3 信號肽預(yù)測網(wǎng)站
SignalP3.0,網(wǎng)址為http://www.cbs.dtu.dk/services/.
1.4 序列比對軟件
Mega4.1.
2 方法
2.1 查找序列
利用NCBI網(wǎng)站的數(shù)據(jù)庫,查找得到具有完整CDS(Coding sequence,編碼序列)的編碼五種禽類即原雞,綠頭鴨,鴻雁,鵪鶉,雉雞五種禽類編碼MHCⅡ類分子β鏈的mRNA序列.
2.2 查找開放閱讀框
利用DNAStar的EditSep對五條序列進行分析,查找相應(yīng)的開放閱讀框(Open Reading Frame,ORF)(表1),并將核苷酸序列翻譯為氨基酸序列.

表1 五種禽類β鏈Genebank登錄號及ORF長度
2.3 分析氨基酸序列
使用SignalP 3.0 Server分析ORF的N端氨基酸序列,根據(jù)從SignalP中分析獲取的Cmax值 (the cleavage site score)、Smean 值(the signal peptide score)和 HMM 值(hidden Markov model)判定是否存在信號肽,并測出信號肽酶切位點,從而確定信號肽序列存在區(qū)段;
2.4 分析信號肽序列的保守性
五條禽類MHCⅡ類分子β鏈信號肽氨基酸序列用Mega軟件進行比對,分析氨基酸的保守位點.
3 結(jié)果與分析
3.1 五種禽類MHCⅡ類分子β鏈信號肽分析
SignalP 3.0 Server是基于神經(jīng)網(wǎng)絡(luò) (Neural Networks,NN)和隱馬可夫模型(Hidden Markov,HMM)兩種算法預(yù)測原核生物和真核生物蛋白質(zhì)信號肽的軟件.前者預(yù)測值表示為Smean,后者預(yù)測值表示為Sprob[3].運用信號肽分析軟件SignalP 3.0分析原雞(Gallus gallus)MHCⅡ類分子β鏈的信號肽存在情況(圖1),SignalP-HMM的結(jié)果表明,該編碼序列含有信號肽的概率為99.9%;SignalP-NN的結(jié)果表明,該編碼序列含有能被信號肽酶切除的信號肽,酶切位點在第26,27位氨基酸之間,N端前26個氨基酸為信號肽區(qū),進一步用DNAStar的EditSep分析信號肽的氨基酸序列,結(jié)果表明26個氨基酸中,有17個非極性氨基酸,占65.38%;6個極性氨基酸,占23.08%;3個帶正電荷的堿性氨基酸,占13.04%.
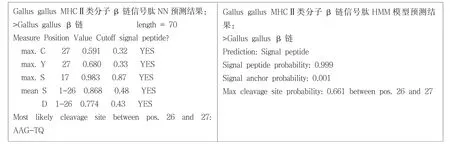
圖1 原雞MHCⅡβ鏈的信號肽分析
運用信號肽分析軟件SignalP 3.0分析綠頭鴨(Anas platyrhynchos)MHCⅡ類分子β鏈信號肽存在情況 (圖2).SignalP-HMM結(jié)果表明,該編碼序列含有信號肽的概率為99.2%,SignalP-NN的結(jié)果表明,該編碼序列含有能被信號肽酶切除的信號肽,酶切位點在第26,27位氨基酸之間,N端前26個氨基酸為信號肽區(qū),進一步用DNAStar的EditSep分析信號肽的氨基酸序列,結(jié)果表明26個氨基酸中,有16個非極性氨基酸,占61.54%;8個極性氨基酸,占30.77%;2個帶正電荷的堿性氨基酸,占7.69%.

圖2 綠頭鴨MHCⅡβ鏈的信號肽分析
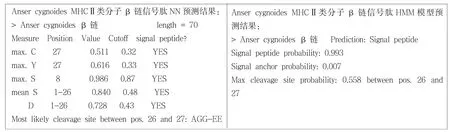
圖3 鴻雁MHCⅡβ鏈的信號肽分析
運用信號肽分析軟件SignalP 3.0分析鴻雁(Anser cygnoides)MHCⅡ類分子β鏈信號肽存在情況 (圖3).SignalP-HMM結(jié)果表明,該編碼序列含有信號肽的概率為99.3%,SignalP-NN的結(jié)果表明,該編碼序列含有能被信號肽酶切除的信號肽,酶切位點在第26,27位氨基酸之間,N端前26個氨基酸為信號肽區(qū),進一步用DNAStar的EditSep分析信號肽的氨基酸序列,結(jié)果表明26個氨基酸中,有16個非極性氨基酸,占61.54%;8個極性氨基酸,占30.77%;2個帶正電的堿性氨基酸,占7.69%.
運用信號肽分析軟件SignalP 3.0分析鵪鶉(Coturnix japonica)MHCⅡ類分子β鏈的信號肽存在情況(圖4).SignalP-HMM結(jié)果表明,該編碼序列含有信號肽的概率為99.9%,SignalP-NN的結(jié)果表明,該編碼序列含有能被信號肽酶切除的信號肽,酶切位點在第26,27位氨基酸之間,N端前26個氨基酸為信號肽區(qū),進一步用DNAStar的EditSep分析信號肽的氨基酸序列,結(jié)果表明26個氨基酸中,有17個非極性氨基酸,占65.38%;7個極性氨基酸,占26.92%;2個帶正電的堿性氨基酸,占7.69%.

圖4 鵪鶉MHCⅡβ鏈的信號肽分析
運用信號肽分析軟件SignalP 3.0分析雉雞(Phasianus colchicus)MHCⅡ類分子β鏈的信號肽存在情況(圖5).SignalP-HMM結(jié)果表明,該編碼序列含有信號肽的概率為99.9%,SignalP-NN的結(jié)果表明,該編碼序列含有能被信號肽酶切除的信號肽,酶切位點在第26,27位氨基酸之間,N端前26個氨基酸為信號肽區(qū),進一步用DNAStar的EditSep分析信號肽的氨基酸序列,結(jié)果表明26個氨基酸中,有18個非極性氨基酸,占69.23%;6個極性氨基酸,占23.07%;2個帶正電的堿性氨基酸,占7.69%.

圖5 雉雞MHCⅡβ鏈的信號肽分析
總結(jié)以上五條禽類MHCⅡ類分子β鏈信號肽生物信息學(xué)的分析結(jié)果(表2),表明五條禽類MHCⅡ類分子β鏈含有信號肽概率均在0.99以上,說明禽類MHCⅡ類分子β鏈N端均含有信號肽,且信號肽長度均為26個氨基酸,酶切位點在第26,27位氨基酸之間.因此,可以確定五條禽類MHCⅡ類分子β鏈的N端前26個氨基酸是信號肽區(qū).五條鏈中從第8位到第21位的氨基酸全部都是疏水性氨基酸,說明從第8位到第21位的氨基酸是信號肽的疏水核心區(qū),而且信號肽酶切位點處的氨基酸殘基是親水性氨基酸殘基,具有ARG-X的模式(R為A或Y或G,X表示親水性氨基酸殘基).

表2 禽類MHCⅡ類分子β鏈信號肽生物信息學(xué)分析
3.2 禽類MHCⅡ類分子β鏈信號肽的保守性分析
五條禽類MHCⅡ類分子β鏈信號肽氨基酸序列用Mega軟件進行比對,如圖6.

圖6 五條禽類MHCⅡ類分子β鏈信號肽的氨基酸序列比較保守性氨基酸殘基用紅色矩形框標出
結(jié)果表明,禽類MHCⅡ類分子β鏈信號肽的26個氨基酸中共有17個位點的氨基酸相同,占信號肽氨基酸總數(shù)的65.4%,保守的氨基酸分別是第一位的甲硫氨酸,第2、4位的甘氨酸,第5位的精氨酸,第9、11位的丙氨酸,第12位的纈氨酸,第13位的亮氨酸,第14位的纈氨酸,第15位的丙氨酸,第16、19位的亮氨酸,第20位的甘氨酸,第21位的甘氨酸,第22位的丙氨酸,第23位的精氨酸,第25位的丙氨酸,第27位的甘氨酸.禽類MHCⅡ類分子β鏈信號肽的26個氨基酸中有16~18個非極性氨基酸,6~8個極性氨基酸,2~3個帶正電的堿性氨基酸,26個氨基酸中有17個位點的氨基酸相同,這說明五種禽類MHCⅡ類分子β鏈的信號肽具有高度的保守性.
4 討論
信號肽位于蛋白質(zhì)的N端,一般由16~26個氨基酸殘基,其中包括疏水核心區(qū)、信號肽的C端和N端3部分.當信號肽序列合成后,被信號識別顆粒(SRP)所識別,蛋白質(zhì)合成暫停或減緩,信號識別顆粒將核糖體攜帶至內(nèi)質(zhì)網(wǎng)上,蛋白質(zhì)合成重新開始.在信號肽的引導(dǎo)下,新合成的蛋白質(zhì)進入內(nèi)質(zhì)網(wǎng)腔,而信號肽序列則在信號肽酶的作用下被切除[4-6].
MHCⅡ類分子作為重要的免疫分子,主要作用是識別和提呈外源性抗原肽,進而激活T淋巴細胞,啟動特異性免疫應(yīng)答.MHCⅡ類分子的生物合成起源于內(nèi)質(zhì)網(wǎng),初步加工后便離開內(nèi)質(zhì)網(wǎng)到達高爾基體,在高爾基體中繼續(xù)進行蛋白質(zhì)的修飾與加工,然后再進一步運輸至早期/晚期溶酶體,在溶酶體內(nèi)與外源性抗原肽結(jié)合,再通過膜泡運送到細胞表面[7,8],供T淋巴細胞識別.在MHCⅡ類分子的生物合成中信號肽對于控制蛋白質(zhì)的分泌路徑和指導(dǎo)蛋白質(zhì)定位特定位置有重要意義.
運用信號肽分析軟件SignalP3.0對五條禽類MHCⅡ類分子β鏈信號肽氨基酸序列進行分析可知,五條禽類MHCⅡ類分子β鏈均含有明確的信號肽,即前26個氨基酸是信號肽區(qū),第8至21位是由中性氨基酸構(gòu)成的疏水區(qū),符合信號肽的特征.MHCⅡ分子具有豐富的多態(tài)性和多基因性,通過分析五條禽類MHCⅡ類分子β鏈的信號肽氨基酸序列可知MHCⅡ類分子β鏈的信號肽的26個氨基酸中有17個氨基酸位點是高度保守的,說明禽類MHCⅡ類分子β鏈的信號肽具有高度保守性,并且它們執(zhí)行相似的功能,即指導(dǎo)MHCⅡ類分子β肽鏈從細胞質(zhì)中遷移到內(nèi)質(zhì)網(wǎng)中繼續(xù)合成.
信號肽的分析對利用標簽蛋白研究目的蛋白在細胞中的表達和定位有重要意義,根據(jù)信號肽的存在與否才能確定報告基因與目的基因的上游還是下游融合,進而避免標簽蛋白與目的蛋白N端的信號肽區(qū)同時被信號肽酶切除,使得融合基因的表達不僅很好地保留熒光蛋白的特性,而且同時也保證了目的蛋白在細胞中的定位功能[9].
〔1〕翟中和,王喜忠,丁明孝.細胞生物學(xué)(第 3 版)[M].北京:高等教育出版社,2007.200~215.
〔2〕劉惠,楊杰,陳軍,等.基于全序列比對相似度預(yù)測信號肽[J].上海交通大學(xué)學(xué)報,2008(42):11~15.
〔3〕孫平楠,周小玲,王正祥.信號肽生物信息學(xué)分析在Neurospora crassa phyA基因鑒定中的應(yīng)用[J].南方醫(yī)科大學(xué)學(xué)報,2009(6):1098~1101.
〔4〕彭佳師,龔繼明.信號肽與蛋白質(zhì)的分選轉(zhuǎn)運[J].植物生理學(xué)報,2011(47):9~17.
〔5〕譚曉林,劉朝奇,鄭蘭英.信號肽對酵母外源蛋白質(zhì)分泌效率的影響[J].生物技術(shù),2010(3):10~16.
〔6〕孫翰昌,龐敏,靳濤.斑馬魚成纖維細胞生長因子(FGFs)信號肽的預(yù)測分析[J].中國獸醫(yī)雜志,2010(6):33~35.
〔7〕Neumann J,Koch N.Assembly of major histocompatibility complex classⅡsubunits with invariant chain[J].FEBS Letters,2005,579:6055~6059.
〔8〕Busch R,Cloutier I,Sekaly RP,et al.Invariant chain protects classⅡhistocompatibility antigens from binding intactpolypeptidesin the endoplasmic reticulum [J].EMBO J,1996,15:418~428.
〔9〕葉紅.新城疫病毒F蛋白-Ii嵌合體與MHC分子結(jié)合特性的研究[D].安徽農(nóng)業(yè)大學(xué),2009.63.
S858
A
1673-260X(2012)08-0020-04
安徽省高校省級科研重點項目(KJ2012A104)
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
